




Absence or severe reduction in the amount of ergosterol in the fungal membrane and its replacement with other sterols have been described as potential antifungal resistance mechanisms in fungi.
Aims and methodsThe ergosterol content in a collection of 51 clinical yeast isolates, including susceptible and resistant strains to amphotericin B and azoles, was estimated by a simple chromatographic method (HPLC-UV).
ResultsA high content of ergosterol was detected for several strains of Candida glabrata, Candida tropicalis or Pichia membranifaciens. In contrast, strains of Cryptococcus neoformans and Dipodascus capitatus had the lowest ergosterol concentrations. No significant correlation was observed between antifungal susceptibility patterns and ergosterol content.
ConclusionsWe conclude from this study that ergosterol content on yeasts may not be associated with specific resistant patterns.
La ausencia o disminución en la cantidad de ergosterol, así como su sustitución por otros esteroles en la membrana, se ha considerado como un posible mecanismo de resistencia de la célula fúngica.
Objetivos y métodosEn este trabajo hemos evaluado la cantidad de ergosterol de una colección de 51 aislamientos clínicos de levaduras, incluyendo cepas sensibles y resistentes a antifúngicos, mediante un sencillo método cromatográfico (HPLC-UV).
ResultadosAlgunas cepas de Candida glabrata, Candida tropicalis y Pichia membranifaciens mostraron mayor contenido en ergosterol que el resto, mientras que las de Cryptococcus neoformans y Dipodascus capitatus presentaron el contenido más bajo. No se observó ninguna relación con suficiente potencia estadística entre el patrón de sensibilidad in vitro y el contenido de ergosterol.
ConclusionesPodemos concluir de este estudio que el contenido en ergosterol no se relaciona sistemáticamente con un patrón de resistencia definido.
Ergosterol is a specific lipid of fungal membranes,22 absent in plants and animals, widely used as an index of fungal mass, particularly in the food-agriculture industry.9,11,18 This special quality has also been exploited by pharmaceutical research as a specific target for antifungal drug development, with this sterol being the focus of many reports in the literature. The mechanisms of action of polyenes and azoles involve ergosterol. Polyenes appear to interact with sterols directly, inducing formation of aqueous pores in lipid bilayers and leading to cell death, whereas azoles inhibit the ergosterol biosynthetic pathway, leading to inhibition of cell growth and replication.5 Both classes of antifungal compounds remain among the most effective available agents in the clinics. Absence, severe reduction, high amount or even ergosterol replacement with other sterols have all been considered as potential mechanisms of resistance against both antifungal classes in fungi.8,13,19,20,21 The fact that several studies on resistant mutant strains have shown changes in sterol pattern suggests a connection between ergosterol content and antifungal drug resistance.24,25 We describe here a simple chromatographic method to analyze ergosterol from yeast cells in order to investigate its potential association with antifungal drug susceptibility patterns.
A total of 51 clinical yeast strains (from 15 different species) were studied, including strains with resistance or decreased susceptibility in vitro to amphotericin B (AMB) and azoles. Isolates were identified by standard mycological methods.7 Two ATCC strains (C. krusei ATCC 6258 and C. parapsilosis ATCC 22019) were included in order to confirm the reproducibility of results.
For susceptibility testing we followed strictly the recommendations proposed by EUCAST.14 Minor modifications were included to improve the growth of Cryptococcus neoformans strains and other species showing poor growth (Rhodotorula spp., Dipodascus spp.).6 Antifungal agents tested were AMB (16-0.03μg/ml); fluconazole, FLC (64-0.125μg/ml); itraconazole, ITC (8-0.015μg/ml); and voriconazole, VRC (8-0.015μg/ml).
Strains were classified as resistant or susceptible taking into account interpretative breakpoints proposed by EUCAST for FLC and VRC,15,16 or CLSI for the remaining agents.12 Of note, AMB breakpoints are not available yet, but clinical resistance has been associated with MIC>1μg/ml.4
Yeast cells were cultured in 10ml of RPMI 1640 medium-2% glucose at 35°C for 48h (2.5×105 cells/ml). Cells were then harvested by centrifugation at 2,500 x g for 5min. The resultant pellet was washed and weighed out after removing all visible water (wet weight). Total sterols were extracted from each pellet as reported previously.1 Prior to chromatographic analysis, each dry residue was dissolved in 1ml of methanol, filtered to remove any particle in suspension, and finally aliquots of 50μl were injected into the HPLC system (LC Module I HPLC Waters). Loss of ergosterol was not detected after the filtration process (data not shown). A flow rate of 1ml/min (methanol 100%) was established, and a detector wavelength of 282nm was selected for quantification purposes. Six solutions of known concentrations of ergosterol (from 1 to 50μg/ml) were also injected into the system to serve as reference for qualitative and quantitative analysis (Standard Curve). These solutions were prepared diluting purified ergosterol powder (Fluka, Madrid, Spain) in methanol. (It is recommended to wait around 10minutes for its complete solubilization). Concentration values, expressed as μg of ergosterol per ml of solvent (methanol), were determined by the wet weight (mg) and ultimately expressed as μg/mg. To test extraction efficiency several experiments were performed by comparing results from samples spiked with known amounts of ergosterol. Experiments were repeated up three times. Moreover, cultures of ATCC strains were also tested repeatedly in order to evaluate the robustness of the method.
Ergosterol was detected and characterized approximately 7.5±0.3min after injection. The standard curve was linear over a concentration range between 1 and 50μg/ml of ergosterol (r2=0.991). The assay showed an acceptable reproducibility, with variation coefficients lower than 10%. The lower limit of detection was established at 1μg/ml. The recovery rate from spiked samples was in the range of 72-76%.
For ATCC strains acceptable variability in results was found (0.21±0.04μg/mg and 0.14±0.04μg/mg for C. krusei and C. parapsilosis respectively after five independent experiments).
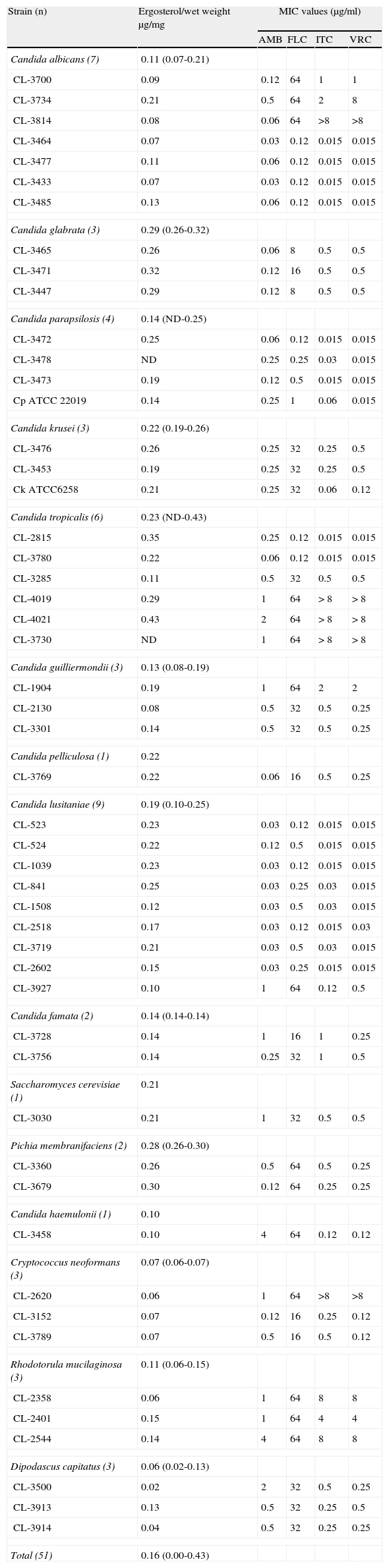
Table 1 summarizes the ergosterol amounts for all yeast strains included in the study. The average ergosterol content was 0.16 (ranging from not detected to 0.43) μg/mg of weight. The wet weight ranged between 28.1 and 151.3mg.
Ergosterol content (means of triplicate) and antifungal susceptibility profile of strains included in the study.
| Strain (n) | Ergosterol/wet weight μg/mg | MIC values (μg/ml) | |||
| AMB | FLC | ITC | VRC | ||
| Candida albicans (7) | 0.11 (0.07-0.21) | ||||
| CL-3700 | 0.09 | 0.12 | 64 | 1 | 1 |
| CL-3734 | 0.21 | 0.5 | 64 | 2 | 8 |
| CL-3814 | 0.08 | 0.06 | 64 | >8 | >8 |
| CL-3464 | 0.07 | 0.03 | 0.12 | 0.015 | 0.015 |
| CL-3477 | 0.11 | 0.06 | 0.12 | 0.015 | 0.015 |
| CL-3433 | 0.07 | 0.03 | 0.12 | 0.015 | 0.015 |
| CL-3485 | 0.13 | 0.06 | 0.12 | 0.015 | 0.015 |
| Candida glabrata (3) | 0.29 (0.26-0.32) | ||||
| CL-3465 | 0.26 | 0.06 | 8 | 0.5 | 0.5 |
| CL-3471 | 0.32 | 0.12 | 16 | 0.5 | 0.5 |
| CL-3447 | 0.29 | 0.12 | 8 | 0.5 | 0.5 |
| Candida parapsilosis (4) | 0.14 (ND-0.25) | ||||
| CL-3472 | 0.25 | 0.06 | 0.12 | 0.015 | 0.015 |
| CL-3478 | ND | 0.25 | 0.25 | 0.03 | 0.015 |
| CL-3473 | 0.19 | 0.12 | 0.5 | 0.015 | 0.015 |
| Cp ATCC 22019 | 0.14 | 0.25 | 1 | 0.06 | 0.015 |
| Candida krusei (3) | 0.22 (0.19-0.26) | ||||
| CL-3476 | 0.26 | 0.25 | 32 | 0.25 | 0.5 |
| CL-3453 | 0.19 | 0.25 | 32 | 0.25 | 0.5 |
| Ck ATCC6258 | 0.21 | 0.25 | 32 | 0.06 | 0.12 |
| Candida tropicalis (6) | 0.23 (ND-0.43) | ||||
| CL-2815 | 0.35 | 0.25 | 0.12 | 0.015 | 0.015 |
| CL-3780 | 0.22 | 0.06 | 0.12 | 0.015 | 0.015 |
| CL-3285 | 0.11 | 0.5 | 32 | 0.5 | 0.5 |
| CL-4019 | 0.29 | 1 | 64 | > 8 | > 8 |
| CL-4021 | 0.43 | 2 | 64 | > 8 | > 8 |
| CL-3730 | ND | 1 | 64 | > 8 | > 8 |
| Candida guilliermondii (3) | 0.13 (0.08-0.19) | ||||
| CL-1904 | 0.19 | 1 | 64 | 2 | 2 |
| CL-2130 | 0.08 | 0.5 | 32 | 0.5 | 0.25 |
| CL-3301 | 0.14 | 0.5 | 32 | 0.5 | 0.25 |
| Candida pelliculosa (1) | 0.22 | ||||
| CL-3769 | 0.22 | 0.06 | 16 | 0.5 | 0.25 |
| Candida lusitaniae (9) | 0.19 (0.10-0.25) | ||||
| CL-523 | 0.23 | 0.03 | 0.12 | 0.015 | 0.015 |
| CL-524 | 0.22 | 0.12 | 0.5 | 0.015 | 0.015 |
| CL-1039 | 0.23 | 0.03 | 0.12 | 0.015 | 0.015 |
| CL-841 | 0.25 | 0.03 | 0.25 | 0.03 | 0.015 |
| CL-1508 | 0.12 | 0.03 | 0.5 | 0.03 | 0.015 |
| CL-2518 | 0.17 | 0.03 | 0.12 | 0.015 | 0.03 |
| CL-3719 | 0.21 | 0.03 | 0.5 | 0.03 | 0.015 |
| CL-2602 | 0.15 | 0.03 | 0.25 | 0.015 | 0.015 |
| CL-3927 | 0.10 | 1 | 64 | 0.12 | 0.5 |
| Candida famata (2) | 0.14 (0.14-0.14) | ||||
| CL-3728 | 0.14 | 1 | 16 | 1 | 0.25 |
| CL-3756 | 0.14 | 0.25 | 32 | 1 | 0.5 |
| Saccharomyces cerevisiae (1) | 0.21 | ||||
| CL-3030 | 0.21 | 1 | 32 | 0.5 | 0.5 |
| Pichia membranifaciens (2) | 0.28 (0.26-0.30) | ||||
| CL-3360 | 0.26 | 0.5 | 64 | 0.5 | 0.25 |
| CL-3679 | 0.30 | 0.12 | 64 | 0.25 | 0.25 |
| Candida haemulonii (1) | 0.10 | ||||
| CL-3458 | 0.10 | 4 | 64 | 0.12 | 0.12 |
| Cryptococcus neoformans (3) | 0.07 (0.06-0.07) | ||||
| CL-2620 | 0.06 | 1 | 64 | >8 | >8 |
| CL-3152 | 0.07 | 0.12 | 16 | 0.25 | 0.12 |
| CL-3789 | 0.07 | 0.5 | 16 | 0.5 | 0.12 |
| Rhodotorula mucilaginosa (3) | 0.11 (0.06-0.15) | ||||
| CL-2358 | 0.06 | 1 | 64 | 8 | 8 |
| CL-2401 | 0.15 | 1 | 64 | 4 | 4 |
| CL-2544 | 0.14 | 4 | 64 | 8 | 8 |
| Dipodascus capitatus (3) | 0.06 (0.02-0.13) | ||||
| CL-3500 | 0.02 | 2 | 32 | 0.5 | 0.25 |
| CL-3913 | 0.13 | 0.5 | 32 | 0.25 | 0.5 |
| CL-3914 | 0.04 | 0.5 | 32 | 0.25 | 0.25 |
| Total (51) | 0.16 (0.00-0.43) | ||||
ND: not detected; ergosterol content were under the limit of quantification defined for the method described in the text. $Clinical resistance has been associated with AMB MIC>1, FLC>2, ITC>0.125 and VRC>0.125 μg/ml (see text and references). In bold those MICs above breakpoints.
Several strains of C. glabrata (3 out of 3 included), C. tropicalis (3/6), Pichia membranifaciens (3/3) or C. krusei (1/3) showed a high content of ergosterol (> 0.25μg/mg). Conversely C. neoformans (3/3) or Dipodascus capitatus strains (2/3) had the lowest concentration (less than 0.07μg/mg). It is worth noting that C. neoformans is an encapsulated yeast, so its ergosterol content may seem lower when correlated to wet weight. However, the low number of representative strains of each species included in the study made it difficult to detect significant differences between species.
The antifungal susceptibility profile is also summarized in Table 1. Results do not show any direct correlation between MICs and ergosterol levels for any of the antifungal agents tested. Calculated Spearman Rho coefficients (SPSS, version 17.0, Madrid, Spain) were as follows: –0.201, –0.126, –0.107 and –0.039 for AMB, FLC, ITC and VRC respectively (p>0.05). For strains showing AMB MICs higher than 1μg/ml a great variability in ergosterol content was detected (Table 1) and differences with AMB susceptible strains were not statistically significant. In the same way, no differences were noted between FLC, ITC or VRC resistant and susceptible strains studied.
Results described here show no significant correlation between MICs and ergosterol content. In fact the ergosterol content estimated in such an heterogeneous collection of yeasts revealed a great degree of variability, even for members belonging to the same species or those included in the same category (resistant or susceptible to a given antifungal). It is worth noting that the few representative strains of some categories (only 4 AMB resistant strains) limit any conclusions that can be drawn. However, several interesting observations can be made. Our results support those recently described by other authors reporting that several mechanisms might contribute to a step-by-step acquisition of resistance in yeasts17 and also, to a variable ergosterol composition.3,19,20,21 In fact, the susceptibility profile may not depend on the membrane sterol content alone. Other constituents of the cell, and especially the cell wall, might contribute to resistance.10 Therefore, only membrane sterol content does not enable us to explain resistance or susceptibility of yeasts to AMB or azoles, since resistant strains could be as rich in ergosterol as susceptible strains,2,21 as it has also been described here. It is unquestionable that molecular characterization of mechanisms for the acquisition of resistance in these strains might clarify the real role of ergosterol in antifungal resistance. In our opinion this is a limitation of this study, so further molecular approaches are warranted to better characterize the role of ergosterol content in yeast resistance.
To our knowledge, this is the first report correlating ergosterol content and antifungal susceptibility in a collection of susceptible and resistant strains using a chromatographic method (HPLC-UV). This HPLC assay has proved to be a useful method for ergosterol characterization and quantification, even superior to those that use sole ultraviolet absorption spectroscopy.23 However, taken into account the multifactorial nature of the resistance in yeasts, any change in the ergosterol content may not be directly associated with a defined pattern of resistance, although it might contribute to the prediction of possible mechanisms. Further molecular studies are needed to fully determine and characterize resistance mechanism(s) in these fungal cells
Conflicts of interestIn the past 5 years, M.C.E. has received grant support from Astellas Pharma, bioMérieux, Gilead Sciences, Merck Sharp and Dohme, Pfizer, Schering Plough, Soria Melguizo SA, the European Union, the ALBAN program, the Spanish Agency for International Cooperation, the Spanish Ministry of Culture and Education, the Spanish Health Research Fund, the Instituto de Salud Carlos III, the Ramon Areces Foundation, the Mutua Madrileña Foundation. He has been an advisor/consultant to the Panamerican Health Organization, Gilead Sciences, Merck Sharp and Dohme, Pfizer, and Schering Plough. He has been a paid guest speaker on behalf of Gilead Sciences, Merck Sharp and Dohme, Pfizer, and Schering Plough.
In the past 5 years, J.L.R.T. has received grant support from Astellas Pharma, Gilead Sciences, Merck Sharp and Dohme, Pfizer, Schering Plough, Soria Melguizo SA, the European Union, the Spanish Agency for International Cooperation, the Spanish Ministry of Culture and Education, the Spanish Health Research Fund, the Instituto de Salud Carlos III, the Ramon Areces Foundation, the Mutua Madrileña Foundation. He has been an advisor/consultant to the Panamerican Health Organization, Gilead Sciences, Merck Sharp and Dohme, Mycognostica, Pfizer, and Schering Plough. He has been a paid guest speaker on behalf of Gilead Sciences, Merck Sharp and Dohme, Pfizer, and Schering Plough.
Other authors: No conflicts

