




Twenty-four worms were collected from the bronchioles in both lungs of a male nine-banded armadillo Dasypus novemcinctus Linnaeus, 1758 captured in Teacalco, Morelos, Mexico. The worms, herein named Metathelazia mexicana n. sp., show a constriction in the esophagus at the level of the nerve ring; males of the species have 7 pairs of papillae, fewer than the other species in the genus. Metathelazia capsulata is the most similar species to Metathelazia mexicana; however, the latter has much shorter spicules. This is the fourth species in the genus known to occur in the New World.
Se recolectó un total de 24 nemátodos de las cavidades bronquiales de un armadillo de 9 bandas Dasypus novemcinctus Lineo, 1758 en la localidad de Teacalco, Morelos, México. Los nemátodos, descritos con el nombre Metathelazia mexicana, muestran una constricción en el esófago al nivel del anillo nervioso y 7 pares de papilas caudales en los machos, número que es menor al observado en las otras especies del género. Metathelazia mexicana es semejante a Metathelazia capsulata en varios rasgos morfológicos pero difiere en el tamaño de las espículas, que son más cortas en la especie aquí descrita. Esta es la cuarta especie del género registrada en el continente americano.
Three species of the genus MetathelaziaSkinker, 1931 have been reported from the lungs of carnivores of the New World (Skinker, 1931; Pence and Dowler, 1979), including Metathelazia californicaSkinker, 1931, Metathelazia bassarisci(Pence and Stone, 1977) Pence and Stone, 1979, and Metathelazia capsulataGerichter, 1948. With the exception of the latter, they all appear to have a Nearctic distribution. Metathelazia capsulata was originally described from the bronchi of carnivores from the Old World (Gerichter, 1948), including the European badger Meles meles, the red fox Vulpes vulpes(Linnaeus, 1758), and the marbled polecat Vormela peregusna(Güldenstädt, 1770). In the New World, the parasite has been reported in the bronchi of the American badger Taxidea taxus (Schreber, 1777) (Pence and Dowler, 1979).
We herein describe a new species of spirurid nematode that was found in the lungs of 1 armadillo collected in Mexico. Interestingly, this represents the first record of a species of Pneumospiruridae in the country.
Materials and methodsTwo nine-banded armadillos were obtained from local hunters in Teacalco, Morelos on 4 January, 2001. They were transported to the Laboratorio de Helmintología Eduardo Caballero y Caballero at UNAM, Mexico City and examined for endoparasites. Protocols for necropsy, extraction and processing of specimens follow Jiménez-Ruiz et al (2006). Voucher specimens of Metathelazia capsulataGerichter, 1948 from the USNPC (nos. 074892 and 084080) were cleared on temporary mounts of lactophenol or glycerin to be studied.
DescriptionMetathelazia mexicana n. sp.
, and relative distances of nerve ring and excretory pore from anterior end, lateral view. 3, ventral view of tail showing the distribution of the caudal papillae. 4, lateral view of tail, showing shape of spicules and gubernaculum.")
Metathelazia mexicana n. sp., scale bar= 100 µm, except figure 1. 1, en face view, ?, showing disposition of cephalic papillae and teeth. Scale bar= 20 µm. 2, anterior end of holotype, showing esophagus, amphid and valvular structures (esophageal sphincters), and relative distances of nerve ring and excretory pore from anterior end, lateral view. 3, ventral view of tail showing the distribution of the caudal papillae. 4, lateral view of tail, showing shape of spicules and gubernaculum.
 and musculoglandular portions (right). The arrow labeled “Lumen” indicates point of change. C= constriction in muscular portion; Exc. Cells= excretory cells. 9, anterior end showing texture of esophagus and relative positions of buccal capsule (BC), division of muscular and musculoglandular esophagus (Div. Es.), and excretory pore (Ex. Pore). 10, spicule of holotype.")
Metathelazia mexicana n. sp. Scale bar= 100 µm for figures 5-7, 9; scale= 50 µm for Figs. 8, 10. 5, relative distances of vulva and anus to the posterior end. 6, esophagus showing constriction in the muscular part of esophagus and nerve ring. 7, esophagus showing texture of the muscular and musculoglandular portions. 8, detail of esophagus showing differences in the lumen in both muscular (left) and musculoglandular portions (right). The arrow labeled “Lumen” indicates point of change. C= constriction in muscular portion; Exc. Cells= excretory cells. 9, anterior end showing texture of esophagus and relative positions of buccal capsule (BC), division of muscular and musculoglandular esophagus (Div. Es.), and excretory pore (Ex. Pore). 10, spicule of holotype.
General. Whitish nematodes when alive, sexually dimorphic. Tegumental sheath covering body; with fine longitudinal striations. Tail ending in a digitiform projection. Stoma surrounded by 2 lateral pseudolabia, each consisting of a central lobe with dorsal and submedial processes (Fig. 1). Four pairs of cephalic papillae; 2 pairs large, sublateral, and posterior to pseudolabia; 2 submedian pairs next to submedial processes (Fig. 1). Amphids at the basis of each central lobe. Buccal capsule heavily cuticularized, dorsoventrally elongated, narrowing posteriad; en face view with 6 teeth arranged in pairs situated dorsally, medially, and ventrally (Fig. 1). Deirids conspicuous, situated at the level of nerve ring. Esophagus clearly divided, anteriorly showing a short tubular section, followed by a wider claviform portion, ending in a protruding valvular structure (Figs. 2, 9). Muscular part includes both short tubular structure and anterior end of claviform portion (Fig 6.). Claviform portion consists of both muscular and glandular tissues (Figs. 2, 7, 8). Intestine with microvilli, walls gradually widening until becoming non-uniform. When contracted, walls fold, forming diverticulae.
Males (Table 1). Nematodes with blunt ends of uniform width along most of body. Seven pairs and 1 unpaired caudal papillae. Precloacal papillae in 2 sublateral pairs plus a single medial papillae on anterior margin of cloaca; postcloacal papillae in 3 sublateral pairs and 2 submedial (Fig. 3). Spicules equal, gubernaculum present (Figs. 4, 10). Tail blunt, ending in digitiform projection (Fig. 4).
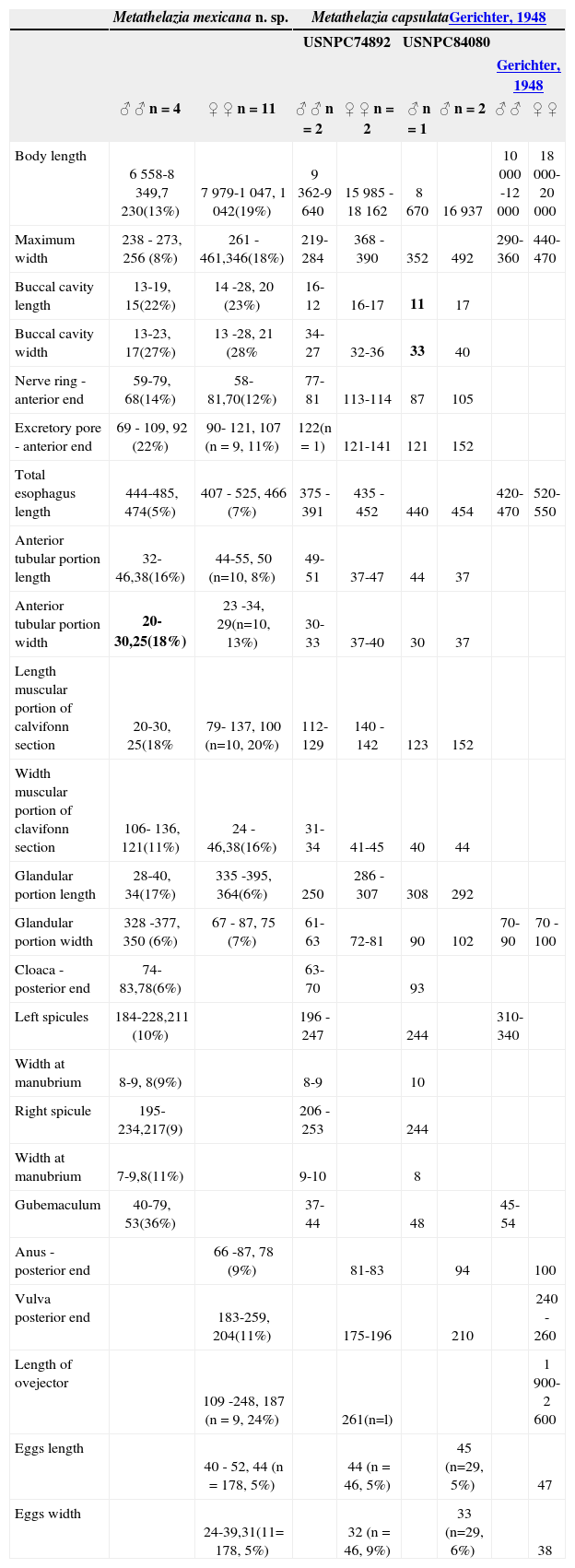
Comparative measurements for Metathelazia mexicana n. sp. and Metathelazia capsulata in the New World. Measurements for M. mexicana include the range, average and coefficient of variation in parenthesis
| Metathelazia mexicana n. sp. | Metathelazia capsulataGerichter, 1948 | |||||||
|---|---|---|---|---|---|---|---|---|
| USNPC74892 | USNPC84080 | |||||||
| Gerichter, 1948 | ||||||||
| ♂ ♂ n = 4 | ♀ ♀ n = 11 | ♂ ♂ n = 2 | ♀ ♀ n = 2 | ♂ n = 1 | ♂ n = 2 | ♂ ♂ | ♀ ♀ | |
| Body length | 6 558-8 349,7 230(13%) | 7 979-1 047, 1 042(19%) | 9 362-9 640 | 15 985 - 18 162 | 8 670 | 16 937 | 10 000 -12 000 | 18 000-20 000 |
| Maximum width | 238 - 273, 256 (8%) | 261 -461,346(18%) | 219-284 | 368 - 390 | 352 | 492 | 290-360 | 440-470 |
| Buccal cavity length | 13-19, 15(22%) | 14 -28, 20 (23%) | 16-12 | 16-17 | 11 | 17 | ||
| Buccal cavity width | 13-23, 17(27%) | 13 -28, 21 (28% | 34- 27 | 32-36 | 33 | 40 | ||
| Nerve ring -anterior end | 59-79, 68(14%) | 58- 81,70(12%) | 77-81 | 113-114 | 87 | 105 | ||
| Excretory pore - anterior end | 69 - 109, 92 (22%) | 90- 121, 107 (n = 9, 11%) | 122(n = 1) | 121-141 | 121 | 152 | ||
| Total esophagus length | 444-485, 474(5%) | 407 - 525, 466 (7%) | 375 -391 | 435 - 452 | 440 | 454 | 420-470 | 520-550 |
| Anterior tubular portion length | 32-46,38(16%) | 44-55, 50 (n=10, 8%) | 49-51 | 37-47 | 44 | 37 | ||
| Anterior tubular portion width | 20-30,25(18%) | 23 -34, 29(n=10, 13%) | 30-33 | 37-40 | 30 | 37 | ||
| Length muscular portion of calvifonn section | 20-30, 25(18% | 79- 137, 100 (n=10, 20%) | 112-129 | 140 -142 | 123 | 152 | ||
| Width muscular portion of clavifonn section | 106- 136, 121(11%) | 24 -46,38(16%) | 31-34 | 41-45 | 40 | 44 | ||
| Glandular portion length | 28-40, 34(17%) | 335 -395, 364(6%) | 250 | 286 - 307 | 308 | 292 | ||
| Glandular portion width | 328 -377, 350 (6%) | 67 - 87, 75 (7%) | 61-63 | 72-81 | 90 | 102 | 70-90 | 70 - 100 |
| Cloaca - posterior end | 74-83,78(6%) | 63-70 | 93 | |||||
| Left spicules | 184-228,211 (10%) | 196 - 247 | 244 | 310-340 | ||||
| Width at manubrium | 8-9, 8(9%) | 8-9 | 10 | |||||
| Right spicule | 195-234,217(9) | 206 - 253 | 244 | |||||
| Width at manubrium | 7-9,8(11%) | 9-10 | 8 | |||||
| Gubemaculum | 40-79, 53(36%) | 37-44 | 48 | 45-54 | ||||
| Anus - posterior end | 66 -87, 78 (9%) | 81-83 | 94 | 100 | ||||
| Vulva posterior end | 183-259, 204(11%) | 175-196 | 210 | 240 - 260 | ||||
| Length of ovejector | 109 -248, 187 (n = 9, 24%) | 261(n=l) | 1 900-2 600 | |||||
| Eggs length | 40 - 52, 44 (n = 178, 5%) | 44 (n = 46, 5%) | 45 (n=29, 5%) | 47 | ||||
| Eggs width | 24-39,31(11= 178, 5%) | 32 (n = 46, 9%) | 33 (n=29, 6%) | 38 | ||||
Females (Table 1). Body widens gradually posteriad. Ovary separated by a conspicuous constriction from uterus. Vulva and anus near posterior end. Ovejector conspicuous. Eggs embryonated. Digitiform terminus at end of tail (Fig. 5).
Taxonomic summary
Type host: Dasypus novemcinctus Linnaeus, 1758.
Type locality: Mexico: Morelos, Teacalco, 18°37’12” N, 99°27’20” W.
Date of collection: 4 January 2001.
Prevalence: 1 of 2 examined, 50%.
Specimens deposited: holotype, male CNHE5434.
Allotype, female CNHE5435; paratypes HWML48407 through HWML48410.
Habitat: secondary bronchioles.
Etymology: the species name refers to the geographic provenance of the worm.
Diagnosis. Metathelazia mexicana can be recognized as unique from all other species in the genus in the shape and structure of the esophagus, which has a constriction at the level of the nerve ring (Figs. 2, 6) being divided into both muscular and musculo-glandular portions, and in the number of caudal papillae (7 pairs). This species is also recognized as unique by the following combination of characters: stoma surrounded by 2 pseudolabia, cuticle with fine longitudinal striations, esophagus separated by a constriction into a tubular and claviform portion, males with 7 pairs of caudal papillae.
Remarks. Metathelazia mexicana differs from M.californica and M. bassarisci in possessing a cuticle with no tuberculations or any other form of projections and in possessing fewer caudal papillae. In addition, M. mexicana can be recognized as distinct from M. californica by possessing 2 pseudolabia and a highly cuticularized buccal capsule. The new species is different from M. bassarisci in that the stoma appears to form petal–like structures and the buccal capsule has the shape of an hour-glass. As noted above, M.mexicana is similar to M. capsulata in that both species posses 4 submedial processes and 2 pseudolabia surrounding the mouth, as well as 6 teeth in the buccal capsule (Fig. 1). However, M. mexicana has only 7 pairs of caudal papillae (compared to 9 pairs in M. capsulata), both the spicules and ovejector are proportionally shorter in M. mexicana, and the esophagus has a constriction at the level of the nerve ring (Table 1, Figs. 4, 5-8). This contrasts with the gradually widening esophagus present in M. capsulata, which in most cases shows an inconspicuous division between muscular and musculoglandular portions.
DiscussionSince there is no consensus in reliable characters to use in the diagnosis, the number of genera included in Pneumospiruridae is still unclear (Pence and Stone, 1977; Wertheim and Chabaud, 1977). Commonly, the structures used to identify the genera include shape of lips, esophagus, spicules, and buccal capsule. An additional criterion is the habitat in the lungs in which the individuals occur. However, Metathelazia naghiensisSpratt, 1980 shows many of the characters used to diagnose both Metathelazia and Pneumospirura and it occurs in diverse parts of the lungs of the long-nosed bandicoot (Spratt, 1980). For those reasons Spratt (1980) assigned the species he described to the oldest genus in the family, and he called for a systematic revision of the family. We found that a similar combination of characters occurs in M. mexicana, and we have used a similar rationale to assign our species to that genus.
Following the classification proposed by Wertheim and Chabaud (1977), M. mexicana should be included in the “capsulata” group, due to the configuration of the pseudolabia and accessory teeth in the buccal capsule. The present constitutes the fourth species of Metathelazia in the New World, and the first reported in the nine-banded armadillo.
In a survey of the helminth parasites of the American badger -Taxidea taxus- in the Great Plains, Pence and Dowler (1979) reported M. capsulata from Kansas with a prevalence of 56%. An additional record includes worms collected from the same host species in Saint Claude, Manitoba (USNPC84080). After measuring (Table 1) and observing the vouchers of M. capsulata(USNPC74892 and USNPC84080), we found them to be different from M. mexicana in 2 features. First, the buccal capsule appears to be dorso-ventrally wider in M. capsulata, and second, there is an additional pair of adcloacal sublateral papillae in M. capsulata.
Pence and Dowler (1979) also noted the absence of intestinal diverticula as a difference between the worms they found and the original description. In his description of the species, Gerichter (1948, p. 77), represents the diverticula as appearing following a smooth portion of the intestine. During our observations we found that most of the worms identified as M. capsulata occurring in American badgers (USNPC74892, 84080) have intestinal diverticula, yet, the degree of folding of the intestinal walls that form the diverticula may be an artifact of the fixation technique used. As an example, individuals of M. mexicana fixed directly in 95% ethanol, without prior relaxation (HWML48407), show these structures much clearer than the homologous structures present in individuals fixed with glacial acetic acid (CNHE5434, CNHE5435, and HWML48408). Additionally, the specimens identified as M. capsulata in the United States and Canada are different from the original description in both measurements of genitalia (Table 1), in the number of caudal papillae, and in the shape of the esophagus. This structure shows a short tubular section in worms occurring in the American badger. The differences among worms from mustelids in North America and carnivores in Palestine, leads us to suspect that specimens identified as M. capsulata in the New World represent a different species. However, we were not able to examine the type specimens of M. capsulata and as a consequence; we are not able to reach a firm conclusion relative to their taxonomic status.
We thank Eric Hoberg and Pat Pillit (USNPC) for sending to us the material they have under their care. Both work in the field and writing of this paper were supported by NSF grants BSR-8612329, BSR-9024816, DEB-9496263, DEB-9631295, DBI-0097019 to SLG, the Walker Graduate Student Funds and the Initiative in Ecology and Evolutionary Ecology of the School of Biological Sciences of UNL to FAJ, and CONACyT-México.