







A new species of Xenosaurus from the Sierra Gorda Biosphere Reserve of northeastern Querétaro, Mexico, is described. The new species differs from all of the other described species of the genus by having usually 2 postrostral scales on each side of the midline (in 84.6% of the specimens, n=26); largest supraoculars that are not, or only slightly, wider than long; postorbital and zygomatic ridges that are widely separated from each other by an intervening row of scales; labiomental rows that usually extend posteriorly from the second or third chinshield (in 92.3% of the specimens, n=26); 23–26 lamellae under the fourth toe (x¯=24.3, n=25); a venter that is immaculate or with only diffuse, scattered dark specks on the sides, and a postorbital region rounded, lacking a canthus temporalis demarcated by enlarged or well-defined scales. The new species inhabits oak forest and a transitional zone between oak forest and subperennial tropical forest at approximately 1 100–1 400m of elevation. The new species is morphologically most similar to X. platyceps from Tamaulipas, but does not appear to be its sister taxon.
Se describe una nueva especie de Xenosaurus de la Reserva de la Biosfera Sierra Gorda en el noreste de Querétaro, México. La nueva especie difiere de todas las demás especies descritas en el género en poseer usualmente 2 escamas postrostrales a cada lado de la línea media (en 84.6% de los ejemplares, n=26); las supraoculares más grandes tan largas como anchas o ligeramente más anchas que largas; crestas postorbital y zigomática ampliamente separadas por una hilera de escamas intermedias; hileras labiomentonales que usualmente se extienden posteriormente a partir del segundo o tercer escudo genial (en 92.3% de los ejemplares, n=26); 23–26 escamas en la superficie ventral del cuarto dedo de la extremidad posterior (x¯=24.3, n=25); vientre inmaculado o con sólo motas oscuras, difusas, dispersas en los lados y una región postorbital redondeada, sin un canthus temporalis delimitado por escamas agrandadas o bien definidas. La nueva especie habita en bosque de encino y una zona de transición entre bosque de encino y bosque tropical subperennifolio aproximadamente entre los 1 100m y 1 400m de elevación. La nueva especie es más similar morfológicamente a X. platyceps de Tamaulipas, pero no parece ser su taxón hermano.
The Middle American lizard genus Xenosaurus Peters, 1861, comprises 11 species and subspecies (see below) that collectively occur from Tamaulipas and Guerrero, on the Atlantic and Pacific versants of Mexico, respectively, south and east to Guatemala (King and Thompson, 1968; Smith and Iverson, 1993; Pérez-Ramos et al., 2000; Nieto-Montes de Oca et al., 2001; Canseco-Márquez, 2005; Zamora-Abrego, 2009). Species of Xenosaurus occur in a considerable altitudinal range (ca. 300-2 360m; King and Thompson, 1968; Smith and Iverson, 1993) and are inhabitants of a wide variety of habitats, ranging from xerophytic tropical scrub to cloud forest to tropical rain forest (King and Thompson, 1968; Ballinger et al., 2000).
In the only monographic treatment of the genus, King and Thompson (1968) recognized only 3 species of Xenosaurus. Two of these species (Xenosaurus newmanorum Taylor, 1949, and Xenosaurus platycepsKing and Thompson, 1968) are restricted to southeastern San Luis Potosí and southwestern Tamaulipas, respectively, in northeastern Mexico. The third species, Xenosaurus granáis (Gray, 1856), was treated as a polytypic species with 5 subspecies: Xenosaurus grandis agrenonKing and Thompson, 1968, which occurs in several localities in the Sierra Madre del Sur in southern Oaxaca; Xenosaurus grandis arboreus Lynch and Smith, 1965, known only from the mountains of the Sierra Madre north of Zanatepec in extreme southeastern Oaxaca; Xenosaurus grandis grandis (Gray, 1856), from west-central Veracruz; Xenosaurus grandis rackhami Stuart, 1941, widely distributed from west-central Chiapas in southeasten Mexico east to Alta Verapaz in Guatemala; and Xenosaurus grandis sanmartinensis Werler and Shannon, 1961, known only from the Sierra de los Tuxtlas in southern Veracruz. In addition, King and Thompson (1968) reported intergrades between X. g. grandis and X. g. rackhami from several localities in north-central Oaxaca.
Four species of Xenosaurus were described after the monographic work of King and Thompson (1968): Xenosaurus rectocollarisSmith and Iverson, 1993, from near Chapulco in southeastern Puebla (Smith and Iverson, 1993), Xenosaurus penai Pérez-Ramos, Saldaña-de la Riva and Campbell, 2000, from isolated highlands of the Sierra Madre del Sur in Guerrero (Pérez-Ramos et al., 2000), Xenosaurus phalaroanthereonNieto-Montes de Oca, Flores-Villela and Cambpell, 2001, from near San Juan Acaltepec in the Sierra Madre del Sur in Oaxaca (Nieto-Montes de Oca et al., 2001), and Xenosaurus tzacualtipantecusWoolrich-Piña and Smith, 2012, from the vicinity of Zacualtipán in the Sierra Madre Oriental of Hidalgo (Woolrich-Piña and Smith, 2012), raising the number of described species and subspecies of Xenosaurus to 11.
However, in an unpublished grant final report, Nieto-Montes de Oca (1999) elevated all of the subspecies of X. grandis to species level. Similarly, in an unpublished M. Sc. thesis, Canseco-Márquez (2005) elevated Xenosaurus g. agrenon, X. g. arboreus, X. g. grandis, and X. g. rackhami to species level, but considered X. g. sanmartinensis as a junior synonym of X. rackhami. These changes were supported by molecular evidence in a phylogenetic study of Xenosaurus by Zamora-Abrego (2009), except that conspecificity of X. g. sanmartinensis and X. g. rackhami was not conclusively demonstrated. In a recent phylogenetic study of Xenosaurus and its fossil relatives, Bhullar (2011) followed the changes proposed by Canseco-Márquez (2005) for the species of Xenosaurus included in his study (X. agrenon, X. grandis, and X. rackhami). In contrast, Woolrich-Piña and Smith (2012) recognized X. agrenon and X. rackhami as distinct species, but treated X. grandis as a polytypic species with 3 subspecies (X. g. arboreus, X. g. grandis, and X. g. sanmartinensis), without any justification.
Dixon et al. (1972) first reported an unidentified species of Xenosaurus in the state of Querétaro. More than 20 years later, Nieto-Montes de Oca (1999) and Nieto-Montes de Oca and Pérez-Ramos (1999), in unpublished final grant reports, reported the presence of an undescribed species of Xenosaurus in the same state. Since then, the Xenosaurus from Querétaro has been treated as a “distinct population” of X. platyceps by González-Espinosa (2002), Lemos-Espinal et al. (2004), and Zamora-Abrego (2004); and as an undescribed species in a number of publications dealing with its ecology and natural history (see below). In addition, Dixon and Lemos-Espinal (2010) and Lemos-Espinal et al. (2012) described the external morphology of the Xenosaurus from Querétaro in some detail. Furthermore, Zamora-Abrego (2009) included the species in a molecular phylogenetic study of Xenosaurus, which supported its distinctness (see below). Nonetheless, the species has not been formally described.
Herein, we formally describe the new species from Querétaro based on 26 specimens, compare it with all known taxa in the genus, and summarize available information about its natural history.
Materials and methodsField work conducted in the state of Querétaro during the last 2 decades by herpetologists at the MZFC led to the collection of a total of 26 specimens of Xenosaurus. These specimens were fixed in 10% buffered formalin, stored in 70% ethanol, and deposited at the MZFC. We compared these specimens with specimens of all of the described species and subspecies of Xenosaurus, including the type series of X. arboreus, X. agrenon, X. penai, and X. phalaroanthereon. A list of the specimens examined is provided in the Appendix. Institutional abbreviations for museums and collections follow Sabaj-Pérez (2010), except for IBH-LT (Estación de Biología Tropical “Los Tuxtlas,” Instituto de Biología, Universidad Nacional Autónoma de México, Veracruz, México).
Whereas we consider the evidence for the specific status of X. agrenon, X. arboreus, X. grandis, and X. rackhami as compelling, we regard the taxonomic status of X. sanmartinensis as still uncertain (Zamora-Abrego, 2009). However, for the sake of completeness, we provisionally include it (as a subspecies of X. rackhami) in the comparisons with the new species.
Nomenclature of scales follows King and Thompson (1968) and Smith and Iverson (1993). Scale counts were performed using a dissecting microscope. Measurements were taken with calipers to the nearest 0.1mm. Head length and snout length were measured from the tip of the snout to the anterior margin of the ear and from the tip of the snout to the anterior margin of the orbit, respectively. In the case of characters examined on both the left and right sides of each specimen, the corresponding conditions are reported in this order, separated by a slash.
DescriptionXenosaurus mendozai sp. nov.
 dorsal view; (middle) lateral view; and (bottom) ventral view. Horizontal lines=4mm.")
 dorsal view; and (right) ventral view.")

Xenosaurus platyceps-González-Espinosa 2002 (in part). Lemos-Espinal et al. 2004 (in part). Zamora-Abrego 2004 (in part).
Diagnosis (Table 1). Xenosaurus mendozai does not appear to possess morphological autapomorphies. However, it may be readily distinguished from all congeners by a unique combination of characters.
Variation in selected characters in the genus Xenosaurus (modified from Nieto-Montes de Oca et al., 2001)
| Characters/taxon | agrenon n=14 | arboreus n=5 | granáis n=14 | rackhami rackhami1n=15 | rackhami sanmartinensis n=16 | newmanorum n=11 | penai n=7 | phalaro anthereon n=16 | platyceps n=13 | rectocollaris n=11 | tzacualti pantecus2n=8 | menáozai3n=26 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Postro strals arranged on each side of midline (medial postrostral absent) | Usually (78.6%) | No | No | No | No | Rarely (9.1%) | No | No | Rarely (7.7%) | No | Rarely (28.6%) | Usually (84.6%) |
| Postorbital and zygomatic ridges | Separate | Separate | Separate | In contact | In contact | Separate | Separate | Usually in contact (81.3%) | Separate | In contact | In contact | Separate |
| Canthus temporalis | Weak | Weak | Weak | Well developed | Well developed | Absent | Well developed | Absent | Absent | Absent | Usually weak, angular along its anterior half | Absent |
| Subdigital lamellae on fourth toe | x¯=25.5(23-28) | x¯=24.4(23-26) | x¯=27.5(24-29) | x¯=28.2(25-31) | x¯=28.4(27-30) | x¯=30.3(29-32) | x¯=25.0(24-27) | x¯=19.4(19-22) | x¯=26.0(23-28) | x¯=20.5(20-22) | x¯=27.5(34-31) | x¯=24.3(23-26) |
| Venter | With dark transverse bars | Uniform pale gray | With dark transverse bars | With dark transverse bars | Usually with dark transverse bars (93.8%) | Usually immaculate or with few dark specks on sides (81.8%) | With dark transverse bars | Immaculate (69%) or with few, dark specks on sides | Usually immaculate or with few dark specks on sides (92.3%) | Immaculate | Usually with small, dark scattered spots | Usually with few, scattered dark specks on sides |
Xenosaurus mendozai differs from all of the other species of the genus north of the Mexican Transvolcanic Belt in usually having (in 84.6% of the specimens, n=26) 2 postrostral scales on each side of the midline (usually 1 medial and 2–3 lateral postrostral scales between the medial postrostral and nasal scales on each side in the other taxa [in 90.9%, 92.3%, and 71.4% of the specimens, n=11, 13, and 8, in X. newmanorum, X. platyceps, and X. tzacualtipantecus, respectively]). It may be further distinguished from these species as follows: from X. newmanorum, by having fewer lamellae under the fourth toe (23–26, x¯=24.3, n=25; 29-32, x¯=30.3, n=11, in X newmanorum); from X. platyceps, by having labiomental rows that usually extend posteriorly from the second or third chinshield (in 92.3% of the specimens, n=26; labiomentals rows usually extending posteriorly from the first chinshield [in 73.1% of the specimens, n=13], in X. platyceps), and from X. tzacualtipantecus, by having the postorbital and zygomatic ridges widely separated from each other by an intervening row of scales (postorbital and zygomatic ridges in contact in X. tzacualtipantecus).
Xenosaurus mendozai may be distinguished from all the other species and subspecies of Xenosaurus south of the Mexican Transvolcanic Belt by 2 combinations of characters. First, X. mendozai differs from all of these taxa, except X. agrenon, in that the largest supraoculars are not, or only slightly, wider than long (largest supraoculars distinctly wider than long in the other taxa), and in usually having (in 84.6% of the specimens, n=26) 2 postrostral scales on each side of the midline (1 medial and 2–3 lateral postrostral scales between the medial postrostral and nasal scales on each side in the other taxa). Second, X. mendozai differs from X. agrenon, X. grandis, X. penai, X. r. rackhami, and X. r. sanmartinensis by having a venter that is immaculate or with only diffuse, scattered dark specks on the sides (usually distinct, dark scattered scales and crossbars on the venter in the other taxa) and the postorbital region rounded, lacking a canthus temporalis demarcated by enlarged or well-defined scales. In the other taxa, the canthus temporalis varies from weak (yet distinct) to well developed, and consists of a longitudinal postorbital series of enlarged scales distinctly set off from the smaller granular temporal scales.
Xenosaurus mendozai may be further distinguished from X. phalaroanthereon and X. rectocollaris by having the postorbital and zygomatic ridges widely separated from each other by an intervening row of scales (postorbital and zygomatic ridges usually in contact [in 81.3% of the specimens, n=16, in X. phalaroanthereon; in all of the specimens in X. rectocollaris]) and by having more lamellae under the fourth toe (23–26, x¯=24.3, n=25; 1922, x¯=19.4 n=16, in X. phalaroanthereon; 20–22, x¯=20.5, n=11, in X. rectocollaris).
Holotype (Figs. 1, 2). Adult male (MZFC 9443), 101.7mm in snout-vent length (SVL), with a moderately stocky body, short limbs (shank length= 12.5mm, 12.3% SVL), and a comparatively short tail (length= 83.0mm, 81.6% SVL).
Head intermediate in size, broadly triangular in dorsal view, moderately thick (length= 26.1mm [25.7% SVL], maximum width=21.0mm [80.2% length], maximum height=12.8mm [48.8% length]). Snout comparatively short (length=9.5mm, 36.5% head length), pointed.
Dorsal head scales generally moderately large, juxtaposed, slightly to markedly convex, conical; many scales on head bluntly keeled, including most scales on snout, between supraorbital semicircles, and between interparietal and supraorbital semicircles, and some scattered supraocular and parietal scales; remaining scales smooth. Rostral moderate in size, 0.6 times as deep as wide, about 0.6 times as wide as mental; posteromedial process absent; with one sulcus at about midway between midline and each lateral border extending anteroventrally for about one-third its depth from its posterior end. Postrostrals 2/2 between midline and nasal scale, moderate in size; medial ones slightly longer than wide; lateral ones slightly larger than medial ones, about 1.5 times as wide as long; 2 small scales between lateral postrostral and nasal scale on left side. Scales on remaining dorsal surface of snout moderately large, irregularly elevated, undifferentiated, without a discernable pattern; 4 enlarged scales between supranasals at level of nostrils; 5 enlarged scales across snout in front of anterior end of supraorbital semicircles. Supraorbital semicircles separated along midline by 1 scale row, except in narrow contact (involving 1 scale of each semicircle) at level of mid orbit. Supraoculo-orbital scales mostly granular. Supraocular scales generally intermediate in size, rounded, convex, bluntly pointed, narrowly separated from each other by single granular scale rows, arranged in 5 rows: 1 main oblique row composed of 2 and 3 largest supraoculars on left and right side, respectively; 1 oblique, immediately medial row parallel to supraorbital semicircles; 1 slightly oblique, immediately lateral row; 1 longitudinal row parallel to superciliaries; and 1 shorter row extending posteriorly from anterior portion of supraorbital semicircles to level of mid orbit between last 2 rows. Largest supraoculars abruptly enlarged relative to anterior and posterior scales in row, slightly larger than scales in immediately medial row, distinctly larger than scales in remaining rows. Superciliaries 8/9, small, roughly square to slightly longer than wide. Minimum count of scales between interparietal and supraorbital semicircle 2/2. Interparietal small. Scales on medial third of parietal area small, irregularly elevated, juxtaposed or narrowly separated from each other by few, scattered granular scales; scales on either lateral third of parietal area distinctly larger and less elevated than those on medial third, rounded or hexagonal, usually separated from each other by 1 row of granular scales; scales in each third roughly uniform in size, not arranged in any definite pattern.
Nasal scale moderately large, about 0.75 times as high as wide, completely encircling nostril at its posterodorsal corner. Supranasals 6/4, small to middle-sized, 4- to 6-sided; postnasals 4/4, small, 4- to 6-sided. Scales in preocular-subocular-postocular series 10/11, small, keeled; preoculars approximately square; suboculars and postoculars moderately elongate. Canthus rostralis indistinct, bordered laterally by uppermost preocular, loreal, and postnasal scales. Loreals 12/12, small to moderately large, irregular, elevated, usually keeled; scales reaching canthus rostralis region 3/3; mínimum scale count between supralabial row and canthus rostralis region 2/3; scales in contact with posterior supranasals and postnasals 3/3; in contact with supralabial row 4/4. Lorilabial scales slightly smaller than lower loreals; markedly larger than suboculars; irregularly elevated, keeled. Zygomatic ridge well developed, composed of moderately large, roughly hexagonal, elevated, keeled scales extending dorsally to parietal-temporal contact region; postocular and zygomatic ridges separated by middle-sized, elevated intervening scales except at their dorsal and ventral ends (dorsal- and ventral- most scales in postocular and zygomatic ridges in contact with each other on each side). Canthus temporalis rounded (slightly angular at its anterior end); parietal and temporal scales grading into each other; adjacent to each other on anterior half of canthus, narrowly separated by 1 row of granular scales posteriorly. Temporal tubercles posterior to zygomatic ridge intermediate in size, moderately convex, bluntly pointed, extending posteriorly to anterior margin of tympanum, ventrally to level of labiomental row; largest ones separated from each other by 1 or 2 granular scale rows (distance between tubercles usually one-third to one- fourth their own diameter). Tympanum large, vertically oval (width=2mm, depth=5mm), its depth 19.1% of head length; covered with a thick membrane with small, granular scales. Supralabials 10/12; anterior ones smooth; gradually becoming slightly higher, keeled posteriorly from level of mid orbit. Infralabials 9/approximately 11 (infralabial-labiomental region misshaped below level of orbit on right side); those anterior to level of orbit large, deep, relatively smooth; gradually becoming smaller, shallower, higher, and keeled posteriorly.
Mental large, bordered posteriorly by 2 enlarged chinshields between infralabials; each chinshield followed by 7 chinshields (similar in size to anteriormost chinshield and each other) in a row extending posteriorly to level of posterior border of orbit (right chinshield row misshaped below level of orbit). First and second scales in infralabial and chinshield rows in contact except for 1 small, isolated scale at their common joint on left side; chinshield and infralabial rows separated posteriorly by labiomental scale row extending posteriorly from lateral side of second chinshield. Scales in labiomental rows small anteriorly, gradually becoming larger, increasingly elevated and keeled posteriorly (row misshaped on right side below level of orbit). First chinshields in medial contact with each other along their anterior halves; their posterior halves and remaining chinshields bordered medially by small to middle-sized, 4- to 6-sided scales; rest of throat and gular region covered by small, rounded gular scales. Gular fold well developed. Additional, well developed transverse pregular fold at level of posterior border of tympanum.
Tubercles on dorsal surface of trunk small, round to oval, slightly convex, roughly arranged in about 34 transverse rows between levels of axilla and groin; tubercles in adjacent rows separated from each other by granular scale rows (distance from slightly less to slightly more their own diameter); tubercles in same row separated from each other by distance between 1 and 2 times their own diameter. Tubercles and granular scales between tubercles slightly larger and slightly smaller, respectively, on paravertebral area than on dorsolateral and lateral areas.
Lateral fold poorly developed; slightly sunken longitudinal strip between dorsal and ventral surfaces, extending from axilla to groin, covered with granular scales abruptly smaller than dorsal and ventral scales. Ventral scales between gular fold and level of axilla intermediate in size, mostly oval, flat, arranged in oblique rows; gradually becoming rectangular, arranged in transverse rows, posterior to level of axilla. Scales on chest and venter roughly square to rectangular, longer than wide; some with central, small, circular depression. Ventral scales along midline between levels of axilla and groin approximately 35; scales in a transverse row at level of midbody about 23. About 4 transverse rows of enlarged, smooth, oval to square to rectangular preanal scales (medial scales in each row larger than lateral ones; posteriormost 2 medial ones largest).
Tubercles on dorsal surface of arms moderate in size, convex, bluntly pointed (point often off center), narrowly separated from each other by 1 granular scale row (distance of about one-fourth to one-half their own diameter). Scales on dorsal surface of thigh moderate in size, mostly flat, juxtaposed or narrowly separated by one granular scale row on anterior surface; gradually becoming slightly smaller, convex, bluntly pointed, and more widely separated by one granular scale row on central surface; markedly smaller, less numerous, flattened, and more widely separated from each other by 2 or 3 granular scale rows on posterior surface. Subdigital lamellae on fourth toe 24/24; length of fourth toe 13mm. Tail circular in cross section, moderately wide at base. Scales on dorsal, lateral, and ventral surfaces arranged in transverse rows. Except on its anterior and posterior ends, rows on ventral surface gradually narrowing dorsally and encircling rest of tail alternating with rows not narrowing dorsally but bifurcating from about middle height into 2 narrow rows encircling tail dorsolaterally and dorsally (i.e., combined length of 2 adjacent rows on ventral surface ≅ combined length of 3 adjacent rows on dorsal surface). Scales on dorsal surface moderate in size, rounded, slightly elevated, separated from each other by 1 granular scale row anteriorly; gradually becoming flat, juxtaposed, square, then rectangular (slightly longer than wide) posteriorly; scales on ventral surface moderately large, juxtaposed, roughly square on anterior rows; gradually becoming smaller, rectangular (longer than wide) on posterior rows.
Color in preservative (Fig. 3). Head uniformly brown except for irregular dark brown area around interparietal scale, one irregular dark brown mark along anterior end of canthus temporalis, and few small, irregular, scattered dark brown spots on rest of head.
Narrow, U-shaped, solid black collar on posterior half of neck, extending anterolaterally to near ear on each side; connected along midline to posterior end of head by narrow, black middorsal line. Neck collar bordered anteriorly by pale brown subocular stripe extending dorsomedially from ventral half of tympanum to black middorsal line on each side; bordered posteriorly by wide, pale stripe extending posterodorsally from gular fold on each side, nearly fusing with stripe from opposite side along midline (separated by narrow black middorsal line extending posteriorly from collar); pale stripes forming concave interspace between collar and first dark crossband on body at level of anterior insertion of arms. Six irregular white spots on neck collar and adjacent portion of neck: 4 at or near midline (1 small one on each side of midline within collar, 1 small and 1 exceedingly small ones on black middorsal line connecting collar and head) and 1 moderately large one at posterior end of each subocular stripe; subocular stripes invaded by 1 narrow, dark brown, roughly transverse bar along their middle and 1 short longitudinal bar extending anteriorly from collar on each side; white spots and dark bars rendering anterior border of collar indistinct.
Five wide, slightly irregular crossbands on dorsum between posterior border of neck collar and level of groin; crossbands separated from each other by narrow, pale brown, poorly defined transverse interspaces; crossbands darker along their anterior and posterior margins than along their mid axis; interspaces with 1–2 white tubercle rows. Arms and legs dull brown, unmarked except for 1 whitish, irregular blotch around each elbow.
Ventral surface ground color dirty cream, immaculate on head, neck and chest; with numerous, somewhat diffuse, scattered dark brown specks surrounded by dark stippling on each side of venter. Ventral surface of arms unmarked; that of legs with 1 wide, short, irregular brown mark on anterior half of thigh and 1 narrower, longer, curved brown line on its posterior half. One small dark spot on center of preanal area.
Tail with 9 wide, uniform dark brown rings; rings separated from posteriormost crossband on body by narrow, poorly defined pale interspace with several white tubercles on each side, alternating with much narrower cream rings along tail; each dark ring (except for first one) with 1 midventral cream spot; spot large, transverselly oriented on anterior rings; gradually becoming smaller, restricted to
Color in life. There are no photographs of the holotype in life. Color in life of other specimens of Xenosaurus mendozai is illustrated in Fig. 3. Dorsal and lateral groundcolor of head and body uniform dark brown; neck collar black; dark crossbands on trunk dark brown, black along their anterior and posterior edges; anterior and posterior margins of collar and interspaces between dark crossbands on trunk cream; interspaces with 1 row of white tubercles; tail with alternating black and cream rings; ventral surface groundcolor cream, usually with few to numerous, small, dark scattered spots and dark stippling on sides of venter. Dixon and Lemos-Espinal (2010) and Lemos-Espinal et al. (2012) described the pale interspace bordering the neck collar as “cream,” the interspaces on the trunk as “rust-colored,” and the eye as “gold colored.” Additional color photographs of X. mendozai are available in Dixon and Lemos-Espinal (2010).
Variation. Rostral partially divided into 3 parts of approximately equal width by 2 narrow, shallow grooves curving ventrolaterally from its dorsal margin to about one-half to two-thirds its height in 10 specimens; divided into 2 parts (left approximately twice as wide as right) in 1 specimen; rostral undivided in remaining specimens. Four postrostrals (2 between midline and nasal scale on each side) in 21 specimens (1 postrostral partly invading midline in 3 specimens); 5 (1 medial and 2 lateral ones on each side) in 4. Internasals 3-5, x¯=4.3 (n=24). Usually 4 small scales bordering nasal scale dorsally (1 obliquely oriented 1 at posterodorsal corner of nasal, 1 above nostril, and 2 above anterior portion of nasal [3 on left side in 4 specimens]). Supraorbital semicircles (n=22) separated by 1 row of small scales in 5 specimens; in narrow contact (involving 1 scale of each semicircle) in 17 specimens; in wider contact (involving 2 or 3 scales of each semicircle) in 5. Supraoculars (n=24) as in holotype in 8 specimens, in 4 and 5 more-or-less complete rows in 8 specimens each. Enlarged supraoculars on 1 side (48 counts) 2–4 (2/3 and 3/2 in 1 specimen each; 3/3 in 6; 3/4 or 4/3 in 8; 4/4 in 8). Superciliaries (48 counts) 7–10, x¯=8.3. Supralabials (49 counts) 11–14, x¯=12.5. Infralabials (50 counts) 811, x¯=9.7. Labiomental row extending anteriorly to first chinshield on each side in 2 specimens; to second chinshield on each side in 15; to second chinshield on 1 side and third chinshield on opposite side in 5; to third chinshield on each side in 3. Anterior chinshields (n=24) in medial contact with each other in 22 specimens; separated by 1 small scale in 2. Second chinshields separated by 2 gular scales in all specimens.
Transverse ventral scale8rows between levels of axilla and groin (n=23) 31–37, x¯=34.5. Ventral scales in a transverse row at level of midbody (n=24) 21–27, x¯=23.1. Subdigital lamellae on fourth toe (n=24) 23–26, x¯=24.3.
Color (in preservative). One to 3 white spots at or near midline on black neck collar in all specimens (1, in 9 specimens; 2 in 5; 3 in 11); spots small to moderately large; usually rounded (elongate, extending into posterior end of middorsal black line connecting collar to head, in 3 specimens). One or 2 white spots, small to middle-sized, rounded or elongate, on posterior half of middorsal black line connecting collar to head in 15 specimens. One large, irregular white spot at posterior end of each subocular stripe (adjacent to neck collar) in 8 specimens. Subocular stripes invaded by few to numerous small dark spots in all specimens; spots without a discernable arrangement in 8 specimens; some roughly arranged in 1 poorly or well defined transverse line at middle of stripes, dividing them into 2, in 14 and 3 specimens, respectively (remaining spots scattered on rest of subocular stripes without any discernable pattern). Neck collar widened anteriorly, covering most of neck and reducing anterior and posterior halves of divided subocular stripes to narrow areas, in 4 specimens. White spots on neck collar and black middorsal line connecting collar to head, and white and dark spots on subocular stripes rendering pale anterior border of collar indistinct in all specimens.
Dark crossbands between neck collar and tail (n=24) 4–6 (4 continuous across dorsum in 13 specimens; 5, in 8; 6, in 1; 1 interspace extending across half of body, absent in opposite half, in 2 specimens [4/5 and 5/4 crossbands in 1 specimen each]). Pale interspaces visible on middorsal surface, absent on sides, in 1 specimen. Two middle- sized, white paravertebral spots on each interspace in 1 specimen.
Ventral surface (n=25) immaculate or nearly immaculate (few diffuse gray specks on sides of venter) in 9 specimens; pattern as that of holotype in remaining 16. Tail black rings in dorsal view (n=25) 9–11, x¯=9.8.
Measurements. Body proportions of 8 adult males (snoutvent length or SVL=97.7–105.6mm, x¯=101.2) and 10 adult females (SVL=99.3–109.2mm, x¯=105.8) are as follows (HW= head width; HL= head length; HH= head height; SHL= shank length; TL= tail length): HW/HL ♂ 0.72–0.84 (x¯=0.78), ♀ 0.71–0.79 (x¯=0.75); HL/SVL ♂ 0.25–0.28 (x¯=0.27), ♀ 0.24–0.27 x¯=0.26); HH/HL ♂ 0.40–0.52 (x¯=0.44), ♀ 0.37–0.51 (x¯=0.42); SHL/SVL ♂ 0.11–0.13 (x¯=0.12), ♀ 0.10–0.12 (x¯=0.11); TL/SVL ♂ 0.76–0.9 (x¯=0.84), ♀ 0.78–0.89 (x¯=0.83).
Taxonomía summaryType material. Holotype. MZFC 9443 (field number RRA 164), adult male from approximately 7km NE of Tilaco, municipality of Landa de Matamoros, Querétaro, México 21°10'53'' N, 99°09'02'' W), 1 348m elevation. Paratypes. Twenty five specimens, all from the municipality of Landa de Matamoros, Querétaro, México: 7 from the same locality as the holotype (MZFC 9422, 9440–42, 9444, 11396–97), 3 from Acatitlán de Zaragoza, 21°11'08.9'' N, 99°09'52.4'' W, 1 260 m (MZFC 5367 [2 specimens], 6285); 5 from 5.5km NNE of Tilaco, 21°10'49.9'' N, 99°08'52.7'' W, 1 302m (MZFC 8475-79); 4 from Tilaco, 21°11'02.8'' N, 99°09'56.2'' W, 1 154m (MZFC 27387-90); 3 from km 10 on road Lagunilla-Tilaco, 21°12'11.9'' N, 99°12'51.1'' W, 1 387m (MZFC 1488-89, 8004); 3 from Cerro La Cañada, approximately 3 km from Tilaco on road to Santa Inés, 21°11'43.4'' N, 99°11'04.2'' W, 1 244m (MZFC 8005, 27385-86).
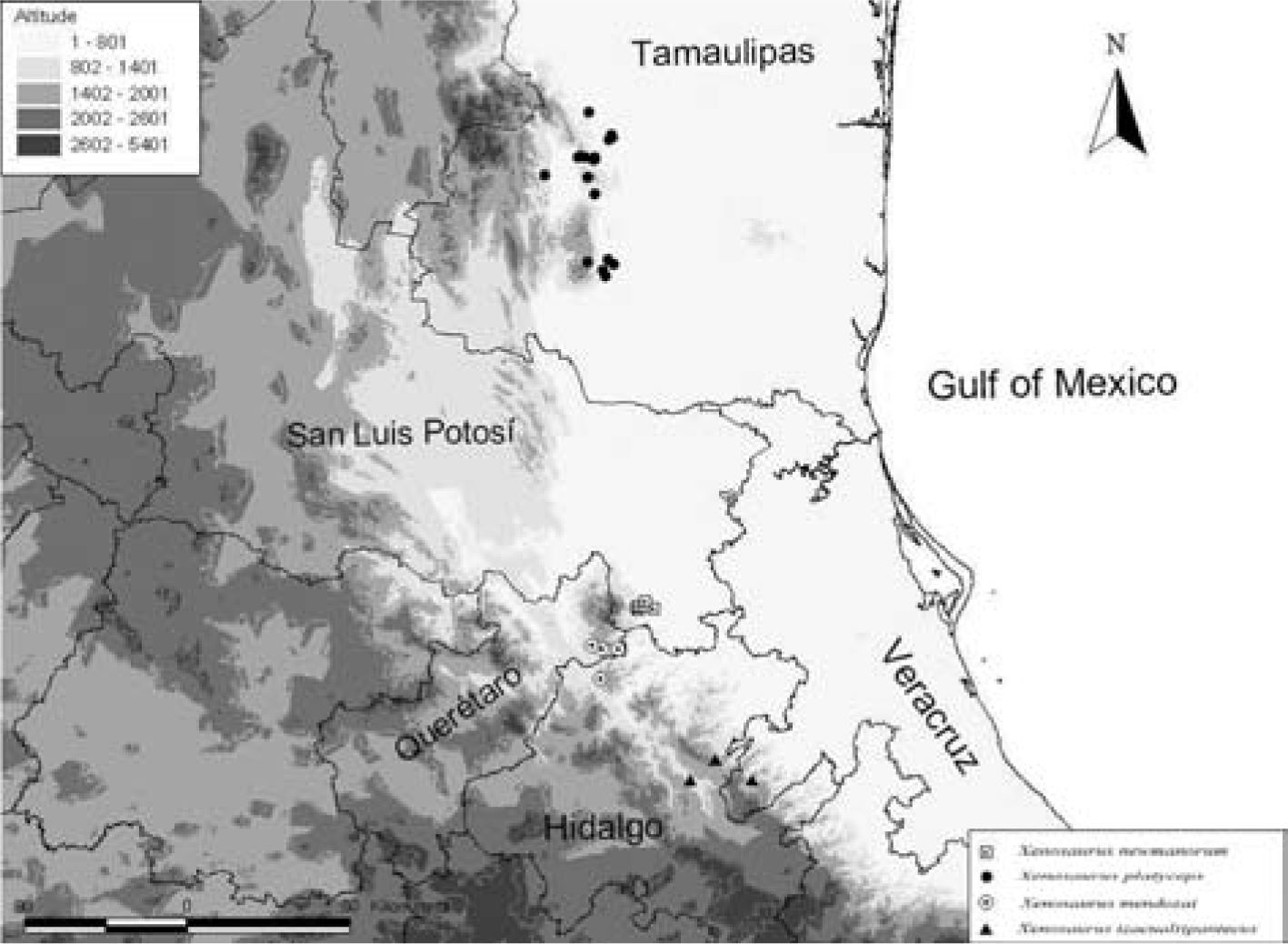
Distribution and ecology. Xenosaurus mendozai is known only from the vicinities of the towns of Tilaco and Acatitlán de Zaragoza in the Municipality of Landa de Matamoros in northeastern Querétaro, Mexico, within the Sierra Gorda Biosphere Reserve (Fig. 4; Dixon and Lemos-Espinal, 2010). Also, it has been recently collected in the vicinity of El Pinalito, municipality of Jacala, Hidalgo (Zamora-Abrego, 2009). Similar to other species of the genus, it exclusively inhabits crevices or holes in boulders (Lemos-Espinal et al., 2004, 2012). The type locality is at 1 348 m of elevation and corresponds to an oak forest. The other localities in which the species has been collected (localities of paratypes) correspond to either oak forest (localities at relatively higher elevations) or to a transitional zone between oak forest and subperennial tropical forest (localities at relatively lower elevations). The dominant tree species are Quercus aristata, Q. castanea, Q. crassipes, Q. laeta, Q. laurina, and Q. mexicana (Zamudio et al., 1992). However, secondary vegetation associated to anthropogenic disturbance is also abundant in the region. The climate in the municipality of Landa de Matamoros is transitional between temperate and tropical with most rainfall occurring between June and October. Mean annual temperature is 20°C and mean total annual rainfall is 773mm (Reyna, 1970).

These lizards are diurnal and insectivorous, with Coleoptera, Orthoptera, and larvae of Lepidoptera being the most consumed prey types (Lemos-Espinal et al., 2004; Zamora-Abrego, 2004). Their body temperature is significantly correlated to environmental temperatures (air and substrate), indicating that these lizards are thermoconformers (González-Espinosa, 2002; Lemos-Espinal et al., 2004). A demographic study of X. mendozai suggested that future increases in environmental temperature would result in negative population trends (Zamora-Abrego et al., 2010). Given its remarkably restricted distribution, this scenario implies a high risk of extinction. Therefore, the highest conservation priority is suggested for this species.
Etymology. The specific name is a patronym for the late Fernando Mendoza Quijano, in honor of whom this species is named in recognition of his significant contributions to the knowledge of the herpetofauna of Mexico, and in particular of the species described herein.
Life history. Similar to other species of the genus, Xenosaurus mendozai is viviparous (Lemos-Espinal et al., 2012). Mean litter size is 2.1 young (range 1– 4). Mating occurs during October and November and births have been observed from June to August (Zamora-Abrego, 2004; Zamora-Abrego et al., 2007). Females are significantly larger than males; however, males have heads of larger dimensions (Lemos-Espinal et al., 2004). Males mature at 80mm SVL when they reach 2 years of age, whereas females do so at 92mm SVL when they reach 3 years of age (Zamora-Abrego et al., 2012). Monthly survival rates of these lizards are remarkably high (between 0.95 and 0.98; Molina-Zuluaga et al., 2013).
Remarks. Several authors noticed the morphological distinctness of the specimens of Xenosaurus from Querétaro, treating them as a “distinct population” of X. platyceps or as an undescribed species (see above). Some of these authors even proposed some diagnostic characters for the species (Nieto-Montes de Oca, 1999; Dixon and Lemos-Espinal, 2010); however, they did not provide data to support those proposals. Although Xenosaurus mendozai does not appear to have morphological autapomorphies, it is distinct from all of the other species and subspecies of the genus by a unique combination of characters, including the presence of 2 or more postrostral scales on each side of the midline; largest supraoculars that are not, or only barely, wider than long; postocular and zygomatic ridges separated by an intervening row of scales; labiomental rows usually extending posteriorly from the second or third chinshield; an intermediate number of subdigital scales on the fourth toe; a venter that is immaculate or with only diffuse, scattered dark specks on the sides; and a postorbital region rounded, lacking a canthus temporalis demarcated by enlarged or well-defined scales. In addition to this morphological evidence, Zamora-Abrego (2009) performed a phylogenetic analysis of Xenosaurus, based on sequences from the mitochondrial genes ND1 and ND4, which provided molecular evidence for the distinctness of X. mendozai. In the Bayesian tree resulting from the analysis, all of the 4 haplotypes of X. mendozai (the Xenosaurus sp. from the Sierra Gorda of Querétaro) formed a strongly supported clade congruent with geography and strongly divergent genetically from the haplotypes of X. newmanorum and X. platyceps (its uncorrected genetic distance from these haplotypes equal to 14.9% and 15.8%, respectively), which together were strongly supported as sister group to X. mendozai. Thus, even though X. mendozai is most similar in scalation and color pattern to X. platyceps and was regarded as a distinct population of X. platyceps by González-Espinosa (2002), Lemos-Espinal et al. (2004) and Zamora-Abrego (2004), they were not each other's sister taxon. This latter result strongly supports the distinctness of X. mendozai.
Xenosaurus mendozai is allopatric from other species in the genus. The nearest species to the type-locality of X. mendozai is X. newmanorum, which occurs near Xilitla in the southeastern extreme of San Luis Potosí approximately 32 km (straight line) to the northeast (King and Thompson, 1968; Lemos-Espinal et al., 2000), followed by X. tzacualtipantecus, from central-east Hidalgo and adjacent Veracruz (Zamora-Abrego, 2009; Woolrich-Piña and Smith, 2012) about 84km to the southeast, and X. platyceps, from several localities in Tamaulipas, the nearest of which to the type locality of X. mendozai is about 280km (straight line) to the north (King and Thompson, 1968; Smith et al., 2000). Interestingly, X. mendozai is more similar in scalation and color pattern to X. platyceps than to X. newmanorum or X. tzacualtipantecus. This suggests that the overall similarity between these 2 species may be at least in part the result of convergence or retained ancestral polymorphism.
In the phylogenetic tree of Zamora-Abrego (2009), the strongly supported clade comprising X. newmanorum, X. platyceps, and X. mendozai was the sister to all other species of Xenosaurus. These phylogenetic relationships are congruent with the geographic distributions of the taxa, because X. newmanorum, X. platyceps, and X. mendozai are the 3 species with the northernmost distribution and, except for X. tzacualtepantecus, the only ones that occur north of the Mexican Transvolcanic Belt. This suggests that the topographic features of eastern (the Sierra Madre Oriental), central (the Mexican Transvolcanic Belt) and southern (the Sierra Madre del Sur) Mexico may be responsible, at least in part, of the observed isolation and differentiation of populations of xenosaurid species, as in many other taxa (e.g., Mateos, 2005; Bryson et al., 2011; Parra-Olea et al., 2012).
Xenosaurus mendozai has only been found in northeastern Querétaro and adjacent extreme northern Hidalgo (Fig. 4), in spite of collecting efforts conducted in nearby regions. This restricted distribution is a common characteristic of other species in the genus, which can be explained by their low mobility (the average distance individuals of X. mendozai moved over a 2-year period was only 11.4m; Zamora-Abrego, 2004) and by the highly specific microhabitat requirements of these crevice-dwelling lizards (Lemos-Espinal et al., 2004, 2012). Other species in the genus, such as X. arboreus, X. newmanorum, X. phalaroanthereon, and X. tzacualtipantecus also have patchy and isolated distributions (King and Thompson, 1968; Nieto-Montes de Oca et al., 2001; Lemos-Espinal et al., 2012; Woolrich-Piña and Smith, 2012).
Three xenosaurid species are listed by the Mexican government as “under special protection” given their rarity and isolated and patchy geographic distributions (X. grandis, X. platyceps, and X. newmanorum; Nom-059-Semarnat-2010Xenosarus mendozai also deserves conservation attention and therefore we recommend that this new taxon be added to the above list. However, given that this species is clearly microendemic with low recruitment rates and with low opportunities to recover from population decline (Zamora-Abrego et al., 2010), we recommend that it be listed as “threatened”.

Support for field work was provided by grants from Conabio (numbers FB340/H-245/96 and FB284/H-250/96) and CONACyT (number 154093) to A. Nieto-Montes de Oca. We thank the following individuals and institutions for loan of specimens: A. de Queiroz and M. Leonard (University of Colorado Museum), J. A. Campbell (University of Texas at Arlington), S. Sroka (University of Illinois Museum of Natural History), A. Muñoz-Alonso (Ecosur), and G. Pérez-Higareda (Estación de Biología Tropical “Los Tuxtlas,” Universidad Nacional Autónoma de México).We thank F. Mendoza-Quijano, G. Zamora-Abrego, I. Durán-Fuentes, A. Zaldívar-Riverón, L. Canseco-Márquez, H. Rivas-García, E. Pérez-Ramos, and R. Reyes-Ávila for invaluable help in collecting the specimens of the type series and/or comparative material. We thank E. Pérez-Ramos for cataloguing specimens in the MZFC collection, J. A. Hernández-Gómez for preparing the photographs of the holotype and for sharing his photographs of the habitat of the new species with us, and Peter Heimes for providing photographs of Xenosaurus mendozai in life. We thank the reviewers for their helpful comments.
Xenosaurus agrenon:Mexico: Oaxaca: Río Sal, Lachao, Juquila (UIMNH 69375 [holotype], 69373-74, 69376 [paratypes]; UCM 41821, 44475-78); 1km N Río Sal (MZFC 9541-44); 2km E Río Sal (MZFC 9545).
Xenosaurus arboreus:Mexico: Oaxaca: North Zanatepec (UCM 39889); the summit of the Sierra Madre, above Zanatepec (UIMNH 56576, holotype); the summit of the Sierra Madre, near Zanatepec (UIMNH 56158, paratype); Sierra Madre, N of Zanatepec (UIMNH 56577-78, paratypes).
Xenosaurus grandis:Mexico: Veracruz: Cuautlapan (MZFC 5920 [3 specimens], 6889-91, 6895-98, 9510); on road from Cuautlapan to Orizaba (MZFC 6892-94).
Xenosaurus mendozai:Mexico: Querétaro: Municipality of Landa de Matamoros: Acatitlán de Zaragoza (MZFC 5367 [2 specimens], 6285); 5.5km NNE of Tilaco (MZFC 8475-79); Tilaco (MZFC 27387-90); km 10 on road Lagunilla-Tilaco (MZFC 1488-89, 8004); Cerro La Cañada, approximately 3km on road Tilaco-Santa Inés (MZFC 8005, 27385-86); approximately 7km NE of Tilaco (MZFC 9422, 9440-44, 11396-97).
Xenosaurus newmanorum:Mexico: San Luis Potosí: Xilitla (MZFC 5928 [2 specimens], 10135); about 7.2km NE Xilitla (MZFC 7484); about 5km N Xilitla, km 268 on road from Xilitla to Tamazunchale (MZFC 8451-55); km 7.2 on road from Xilitla to Tamazunchale (MZFC 6521). No specific locality (MZFC 3476).
Xenosaurus penai:Mexico: Guerrero: Cerro Pico del Aguila (16°57' N, 98°19' W), Sierra de Malinaltepec (IBH 6414, holotype); foothills on E face Cerro Pico del Aguila (MZFC 7099, paratype); on road from El Terrero to Cerro Pico del Aguila (an adult female [MZFC 8479] and its litter [MZFC 7485-87, 8456]; paratypes).
Xenosaurus phalaroanthereon:Mexico: Oaxaca: Sierra Madre del Sur, Municipality of Santa María Ecatepec; about 4km NE San Juan Acaltepec (MZFC 7093-95, 7097-98; UTACV R-46031-32; paratypes); about 6.1km SW San Juan Acaltepec (16°14.93' N, 95°57.29' W), 2005m (MZFC 12223-24 [paratypes], 12225 [holotype], 12226, 12228-31 [paratypes]; UTACV R-46033 [paratype]).
Xenosaurus platyceps:Mexico: Tamaulipas: km 25 on road from Jaumave to Ciudad Victoria (MZFC 5524 [2 specimens], 5543); about 1.1km NE from turnoff (sic), on old road to Rancho El Cielo (MZFC 8517); 3km SE Estación Canindo, Reserva de la Biósfera Rancho El Cielo (MZFC 8518); on road from Gómez Farías to Ejido Azteca (MZFC 8519-21); 18.9km SW Ciudad Victoria (MZFC 9557-60); 21.7km W Ciudad Victoria, on Mexico Hwy 101 on road to Jaumave (MZFC 10051).
Xenosaurus rackhami rackhami:Guatemala: Alta Verapaz: Finca El Volcán (UTACV R 24844-45); Mexico: Chiapas: Selvas El Ocote (ECO-SC-H 34, 100; MZFC 6886-87); km 23 on road from Ocozocoautla to Apic Pac (MZFC 6888); Predio Filadelfia, km 21.7 on road from Ocozocoautla to Apic Pac (MZFC 9552-56); NW Cintalapa (UTACV R 31031, 31066); Cerro del Sumidero, 10.7mi N Tuxtla Gutiérrez (UCM 19028).
Xenosaurus rackhami sanmartinensis:Mexico: Veracruz: Los Tuxtlas (MZFC 4683); Los Tuxtlas, Sierra de Santa Martha (MZFC 6100); El Ariete-Barrava, Bastonal, Sierra de Santa Martha (UNAM-LT 2712-13); Bastonal, Sierra de Santa Martha (MZFC 9564, UNAM-LT 2794); La Asociación, Sierra de Santa Martha (UNAM-LT 2714, 2716); Azufrera, Sierra de Santa Martha (UNAM-LT 2715); crater of Volcán San Martín (MZFC 10553-54); Volcán San Martín (MZFC 9563, 9565; UTA R-29979, 36582, UNAM-LT 2767).
Xenosaurus rectocollaris:Mexico: Puebla: 5km NE Azumbilla, 28 air km NNE Tehuacán (MZFC 113); 9km E Chapulco (MZFC 5719 [3 specimens]); 8 km E Chapulco (MZFC 5923 [2 specimens], 5924, 7554, 9511-12); about km 10 on road from Tehuacán to Orizaba (MZFC 10046).
Xenosaurus tzacualtipantecus: Mexico: Hidalgo: La Mojonera (MZFC 6284, 9513-17, 10136-37).