


Forestry plantations, and particularly those based on Eucalyptus, are known to have negative effects on native avifauna. However, abandoned plantations might provide habitat for some native birds due to the presence of a native understory. Bird diversity between native forest stands and abandoned Eucalyptus plantations with native understory, at the South American temperate rainforest, was compared. Bird richness and composition did not differ between the native vegetation and the abandoned types. We identified 21 species of birds, being 19 of those (90%) shared between vegetation types. Most frequent species were Scelorchilus rubecula, Sephanoides sephaniodes and Elaenia albiceps, whereas Campephilus magellanicus and Zonotrichia capensis occurred only in the native vegetation. The plantation bird assemblage was a subset from the species pool of the native vegetation, with occurrence more variable than species identity between vegetation types. The presence of native understory vegetation at the plantation may explain the presence of some dispersal-restricted species such as rhynocryptids. Nevertheless, abandoned Eucalyptus plantations do not replace native forests as there are no species exclusive to this vegetation type and some species seem to be unable to use Eucalyptus trees.
Las plantaciones forestales, especialmente las de eucalipto, son conocidas por tener efectos negativos sobre la avifauna nativa. Sin embargo, las plantaciones abandonadas pueden proporcionar hábitat para algunas aves nativas gracias a la presencia de sotobosque nativo. Se comparó la diversidad de aves entre bosques nativos y plantaciones abandonadas de Eucalyptus con sotobosque nativo, en el bosque templado sudamericano. La riqueza y composición de especies no difirió entre la vegetación nativa y las plantaciones abandonadas. Se identificaron 21 especies de aves, siendo 19 de ellas (90%) compartidas entre ambos tipos de vegetación. Las especies más frecuentes fueron Scelorchilus rubecula, Sephanoides sephaniodes y Elaenia albiceps, mientras que Campephilus magellanicus y Zonotrichia capensis estuvieron presentes solo en la vegetación nativa. El ensamble de aves de la plantación fue un subconjunto del ensamble de aves de la vegetación nativa, siendo más variable, entre tipos de vegetación, la cantidad de registros obtenidos para las especies que su identidad. La presencia de vegetación nativa en el sotobosque de la plantación de eucalipto podría favorecer la presencia de algunas especies dependientes del bosque y de movimientos restringidos como los rinocríptidos dentro de este tipo de vegetación. Sin embargo, las plantaciones abandonadas de Eucalyptus no reemplazan al bosque nativo ya que no hay especies exclusivas de este tipo de vegetación y algunas aves parecen no ser capaces de utilizar los árboles de Eucalyptus.
Forestry plantations around the world, and particularly those based on Eucalyptus spp. monocultures, are known to have negative effects on native avifauna, causing lower species richness and abundance, as result of the loss of suitable foraging habitats and key structures such as perches and natural cavities (Barlow, Mestre, Gardner, & Peres, 2007; Calvino-Cancela, 2013; de la Hera, Arizaga, & Galarza, 2013; Hsu, French, & Major, 2010; Law, Chidel, Brassil, Turner, & Kathuria, 2014; Loyn, McNabb, Macak, & Noble, 2007; Marsden, Whiffin, & Galetti, 2001). The establishment of large areas of exotic timber monocultures is of particular concern when they occur in avian diversity hotspots, such as the Valdivian temperate rainforests. Despite being a highly biodiverse region, the Valdivian rainforest is being replaced by exotic tree plantations (Echeverría et al., 2006), at a rate of ∼7,000ha per year, currently occupying 185,915ha (from which Eucalyptus plantations represent ca. 42%, according to data of the Chilean Forestry Institute: www.infor.cl). In that context, abandoned plantations are becoming increasingly common, but their value for native avifauna remains little understood.
Due to its biogeographic context, Chilean avifauna resembles an island biota with species-depauperate forests with many endemic species (479 bird species, 12 of them endemic, for continental Chile, island territories and the Antarctic peninsula), which usually have broad latitudinal and altitudinal distribution ranges (Vuilleumier, 1985). Despite this, there is a low species turnover (i.e., low beta diversity) at the country-size scale (Cody, 1970), habitat structure at the local scale determines avifauna assemblage differentiation among different temperate forest types in southern Chile (Fontúrbel & Jiménez, 2014). Southern Chile rainforests typically present 25 bird species (excluding raptors and scavengers). Abandoned plantations are expected to be less hostile to native avifauna due to the presence of native understory vegetation regrowth (Fontúrbel, Jordano, & Medel, 2015). However, as habitat structure largely differs between native forest stands and abandoned Eucalyptus plantations (Fontúrbel et al., 2015), we hypothesize that bird assemblages would be different, with lower species richness and occurrences in the abandoned plantations. In order to determine if abandoned Eucalyptus plantations may constitute suitable habitat for native bird species, we assessed differences in species richness and composition of bird communities from native forests and abandoned plantations.
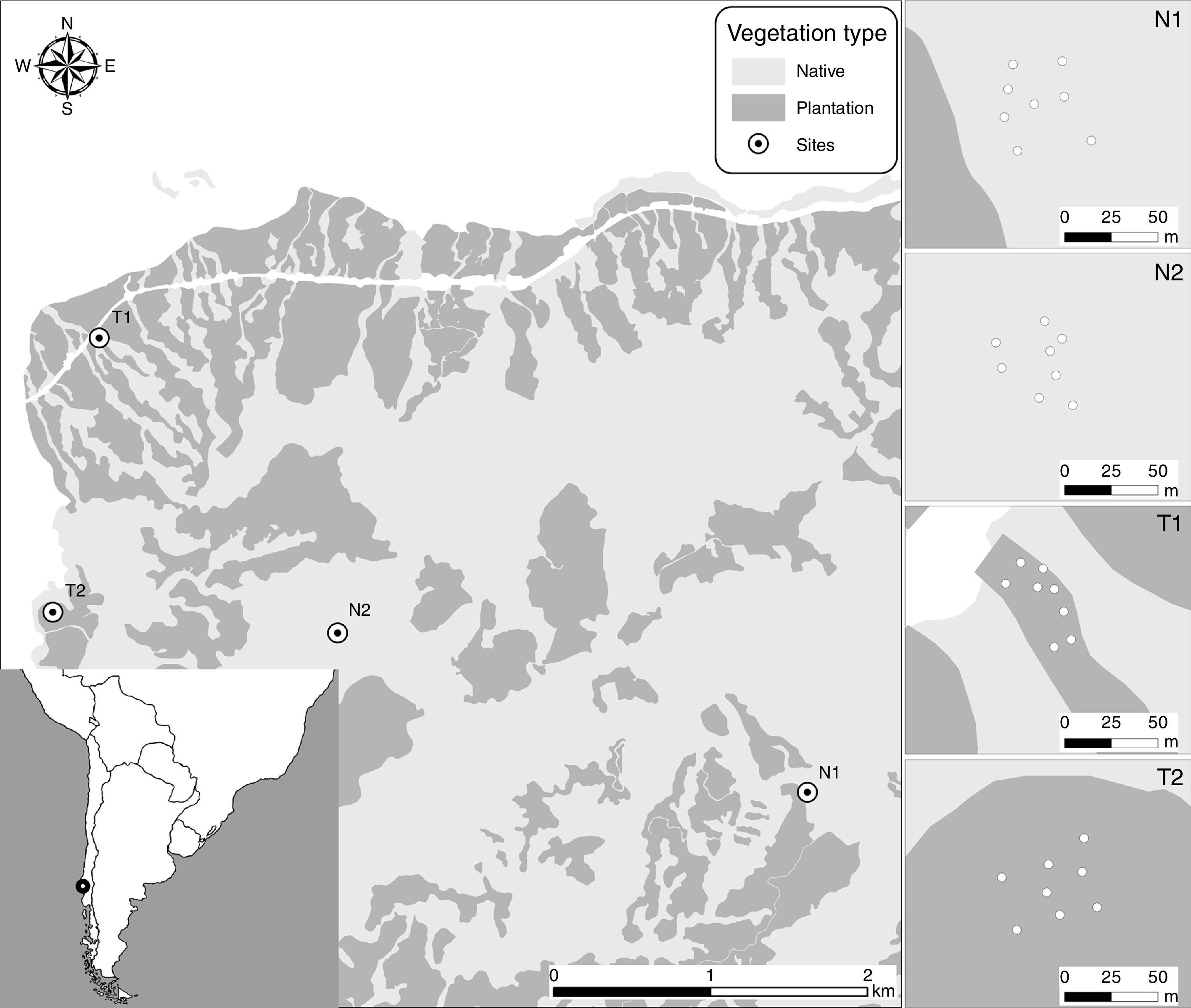
We conducted this research at the Valdivian Coastal Reserve (39°57' S, 73°34' W), a private protected area of The Nature Conservancy (Delgado, 2010), which falls within a biodiversity hotspot (Myers, Mittermier, Mittermier, da Fonseca, & Kent, 2000). This Reserve protects 50,530ha of Valdivian temperate rainforest (∼50% of this ecosystem in Chile). This highly threatened biodiversity hotspot coexists with exotic plantations (currently occupying ∼200,000ha of that ecosystem), which are continuously encroaching and replacing native habitats. Nevertheless, little is known about bird diversity in productive lands (either current or abandoned) in this ecosystem. Since part of the land that the Valdivian Coastal Reserve currently occupies was previously owned by forestry companies, it includes 2 major vegetation types: (1) native stands dominated by Nothofagus spp. trees and a diverse understory vegetation, and (2) abandoned plantations of the exotic tree Eucalyptus globulus (planted 12–20 years ago; never managed or harvested; occupying ∼12,000ha) with abundant native understory vegetation regrowth. Those vegetation types are neighboring and highly intertwined, as fragmentation is negligible; therefore, there are no non-forested areas among them. Detailed information of the study site is available on Fontúrbel et al. (2015). Native vegetation is dominated by Nothofagus dombeyi, N. pumilio and Eucryphia cordifolia as canopy species, whereas the exotic E. globulus is the only canopy species present at the plantation site. Understory vegetation in the native stands is dominated by Laurelia philippiana, Lomatia ferruginea, Drimys winteri, and Mitraria coccinea, with sparse patches of the native bamboo Chusquea quila and a few Lapageria rosea vines. At the plantation sites, understory vegetation is dominated by Aristotelia chilensis, Rhaphithamnus spinosus, Ugni molinae, Luma apiculata, thick C. quila patches, and abundant L. rosea vines climbing on the E. globulus trunks.
We located 2 stands of native vegetation (old-growth forest stands, 0.25ha) and 2 stands of abandoned plantations (0.25ha). Sampling site selection was aided by aerial images and field surveys, aiming to avoid mixing vegetation types. Sampling sites were separated at least 1.8km from each other. At each sampling site we conducted 8 bird censuses (in 8 different sampling points, resulting in 16 sampling points per vegetation type; Fig. 1) during the austral summer (December–March, 2 censuses per month), in order to quantify the avifauna present. Censuses were made from 08:00 to 10:00h, using point counts according to Bibby, Burgess, Hill, and Mustoe (2000). At each point count, we used a fixed radius of 40m for 10min, a standardized technique that allows an adequate estimation of the avifauna diversity in these forests, based on the number of individuals seen or heard, with no major detectability differences (Jiménez, 2000). As the bird species typically found on these forests could be classified as medium forest-dependent (following Rueda-Hernandez, MacGregor-Fors, & Renton, 2015), we do not expect significant inter-species detectability differences. Censuses were not conducted on windy or rainy days (Bibby et al., 2000).

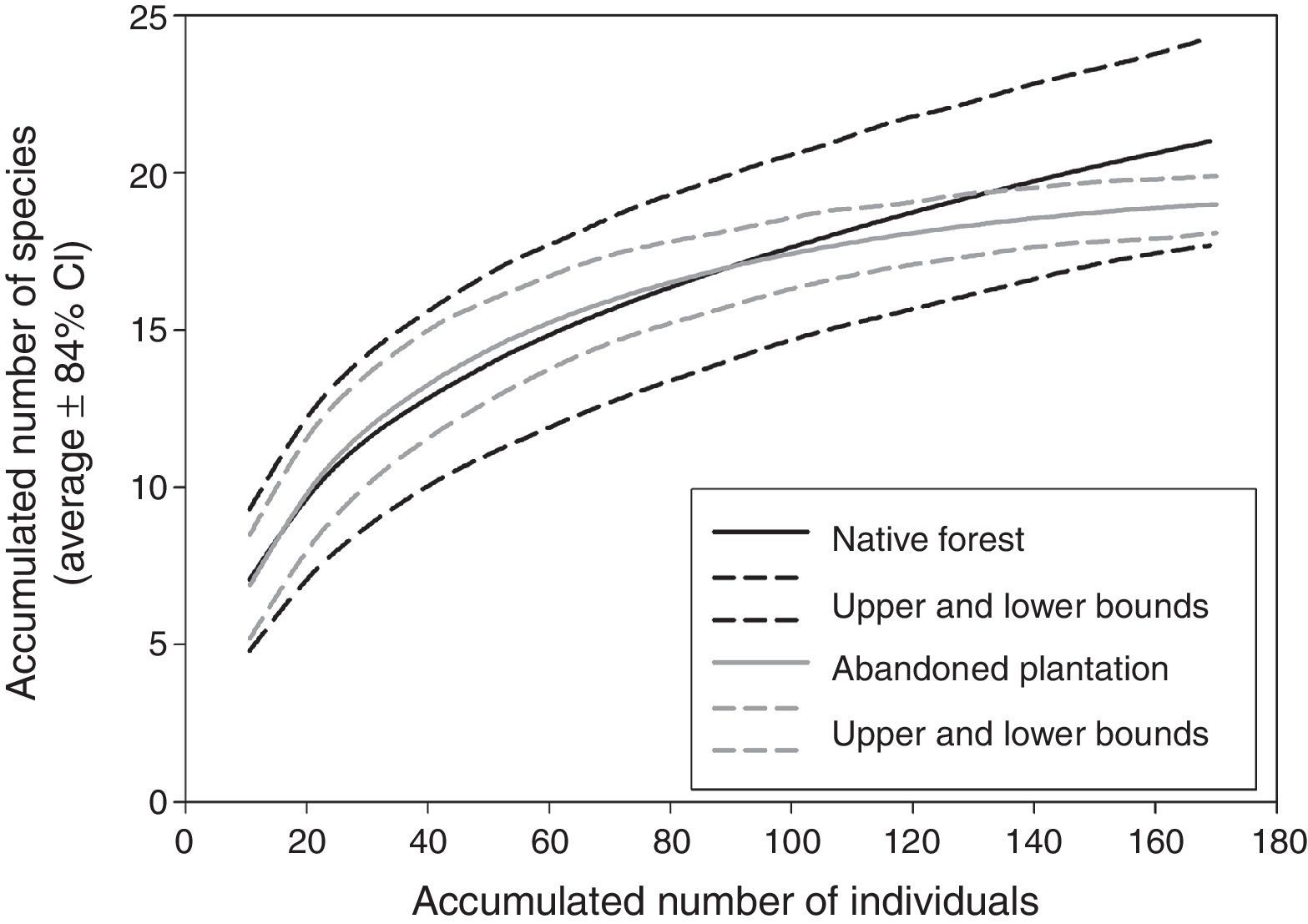
We considered each counting point as a replicate (16 per vegetation type). To compare mean species richness between native vegetation and abandoned plantations, we constructed rarefaction curves based on the number of individuals per species detected at each vegetation type using EstimateS 9.1 (Colwell, 2013). For each rarefaction curve, we depicted the estimated species richness (Sest) and the upper and lower bounds based on the 84% confidence intervals (Hanula, Horn, & O’Brien, 2015; MacGregor-Fors & Payton, 2013; Rueda-Hernandez et al., 2015). The number of individuals per plot (i.e., the abundance of all species recorded at a given plot) was compared using a one-way nested Anova, with point counts nested within each vegetation type (Castaño-Villa, Estevez, & Fontúrbel, 2014). Normality of abundance data was assessed through a Shapiro–Wilk test, and homogeneity of variances through a Levene test.
Bird assemblage composition differences between vegetation types was assessed using a one-way analysis of similarity (Anosim), which is a non-parametric test based on permutations that compares within- and between-group similarities, working similarly than an Anova test to compare species composition among groups (Clarke, 1993). Anosim was conducted using the Bray–Curtis distance, a count-based measurement widely used in comparisons of bird communities (Barlow et al., 2007; Fontúrbel & Jiménez, 2014). Complementarily, we used a similarity percentage (Simper) test to rank species in a decreasing similarity degree of the Anosim test. Anosim and Simper tests were performed using PAST 2.15 (Hammer, Harper, & Ryan, 2001).
We recorded 21 species at the study area. Twenty-one species were present at the native stands, whereas 19 species were found at the abandoned plantation (Table 1). Rarefaction curves, based on the number of individuals, show that mean species richness values between both vegetation types are not statistically different, as both rarefaction curves (and their 84% confidence intervals) overlapped at the same number of records (Fig. 2). The mean number of individuals registered did not differ between the native vegetation and the abandoned plantation (F1,28=0.27; p=0.608). Most frequent species were the Chucao Tapaculo (Scelorchilus rubecula) at both native forest (22.7% and 25.6% of the records) and abandoned plantation (18.4% and 22.2%) vegetation types; the Green-backed Firecrown hummingbird (Sephanoides sephaniodes; 9.1% and 15.9% at native vegetation stands, and 11.2% and 8.3% at plantation sites), and the White-crested Elaenia (Elaenia albiceps; 11.4% and 12.2% at native vegetation stands, and 17.3% and 18.1% at plantation sites).
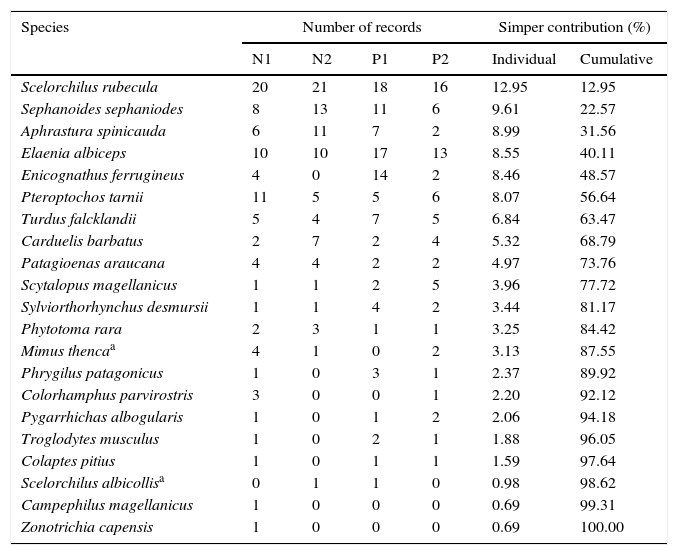
Bird species identified at native (N) and plantation (P) sampling sites (8 censuses per site). Total number of records per site, individual and cumulative similarity contribution percentages (after a Simper test) are shown. The list was arranged based on the Simper similarity values (i.e., species with similar number of records among sampling sites appear at the top of the list).
| Species | Number of records | Simper contribution (%) | ||||
|---|---|---|---|---|---|---|
| N1 | N2 | P1 | P2 | Individual | Cumulative | |
| Scelorchilus rubecula | 20 | 21 | 18 | 16 | 12.95 | 12.95 |
| Sephanoides sephaniodes | 8 | 13 | 11 | 6 | 9.61 | 22.57 |
| Aphrastura spinicauda | 6 | 11 | 7 | 2 | 8.99 | 31.56 |
| Elaenia albiceps | 10 | 10 | 17 | 13 | 8.55 | 40.11 |
| Enicognathus ferrugineus | 4 | 0 | 14 | 2 | 8.46 | 48.57 |
| Pteroptochos tarnii | 11 | 5 | 5 | 6 | 8.07 | 56.64 |
| Turdus falcklandii | 5 | 4 | 7 | 5 | 6.84 | 63.47 |
| Carduelis barbatus | 2 | 7 | 2 | 4 | 5.32 | 68.79 |
| Patagioenas araucana | 4 | 4 | 2 | 2 | 4.97 | 73.76 |
| Scytalopus magellanicus | 1 | 1 | 2 | 5 | 3.96 | 77.72 |
| Sylviorthorhynchus desmursii | 1 | 1 | 4 | 2 | 3.44 | 81.17 |
| Phytotoma rara | 2 | 3 | 1 | 1 | 3.25 | 84.42 |
| Mimus thencaa | 4 | 1 | 0 | 2 | 3.13 | 87.55 |
| Phrygilus patagonicus | 1 | 0 | 3 | 1 | 2.37 | 89.92 |
| Colorhamphus parvirostris | 3 | 0 | 0 | 1 | 2.20 | 92.12 |
| Pygarrhichas albogularis | 1 | 0 | 1 | 2 | 2.06 | 94.18 |
| Troglodytes musculus | 1 | 0 | 2 | 1 | 1.88 | 96.05 |
| Colaptes pitius | 1 | 0 | 1 | 1 | 1.59 | 97.64 |
| Scelorchilus albicollisa | 0 | 1 | 1 | 0 | 0.98 | 98.62 |
| Campephilus magellanicus | 1 | 0 | 0 | 0 | 0.69 | 99.31 |
| Zonotrichia capensis | 1 | 0 | 0 | 0 | 0.69 | 100.00 |
 and plantation (gray lines) vegetation types. Solid lines represent the estimated species richness (Sest), and dashed lines represent the upper and lower bounds based on 84% confidence intervals.")
Avifauna composition did not differ between native and plantation vegetation (Anosim R=0.061, p=0.053, N=32). The 2 species absent from the abandoned plantations were the Magellanic Woodpecker (Campephilus magellanicus) and the Rufous-collared sparrow (Zonotrichia capensis). The species that contributed the most to composition similarity were S. rubecula, S. sephaniodes, E. albiceps, Aphrastura spinicauda, and Pteroptochos tarnii, with a cumulative similarity contribution of 48.4%. Colaptes pitius, Scelorchilus albicollis, C. magellanicus, and Zonotrichia capensis were the most dissimilar species between vegetation types (Table 1).
Avifauna richness did not differ between abandoned plantations and native vegetation types in terms of species richness or composition, as both vegetation types shared 90% of their bird species, being the plantation avifauna a subset of the native vegetation species pool. Despite the limitations of our reduced number of censuses and replicates, our empirical evidence suggest that abandoned plantations are not ‘green deserts’, as they are usually considered, but they can hold many forest-dependent species. These results are based on a limited sample size and, consequently, they are open to further confirmation. Even though, our results were opposite to what was reported for similar comparisons in other temperate forests (e.g., Calvino-Cancela, 2013; de la Hera et al., 2013), in which depauperate bird communities were found at exotic tree-dominated plantations, with a few generalist species dominating the community. In this case, we found many native forest-dependent species at the abandoned plantations, pinpointing the frequent records of 2 rhynocryptid species with limited dispersal (S. rubecula and P. tarnii) and an understory insectivore (Sylviorthorhynchus desmursii) that are known to be negatively affected by habitat fragmentation (Castellón & Sieving, 2006; Díaz, Armesto, & Willson, 2006). However, a similar pattern was reported in Australia by Hsu et al. (2010), in a forest system where plantations were physically connected to native forest stands (as in our study site), stressing the role of the matrix type and the landscape configuration on bird occupation. Our results were, however, opposite to those found at Eucalyptus plantations elsewhere (Barlow et al., 2007; Loyn et al., 2007; Marsden et al., 2001), in which there were significantly lower species richness at the plantations, compared to the native forests. In those cases, the plantation understory was managed and regularly cleared. Having developed understory vegetation is known to promote enhanced avifauna (Nájera & Simonetti, 2009), because structural complexity is a key factor determining bird diversity (Castaño-Villa, Ramos-Valencia, & Fontúrbel, 2014).
The avifauna assemblage found at the abandoned plantations was a subset of the species found at the native stands, as there are no bird species exclusive to the abandoned plantations. The Magellanic Woodpecker (C. magellanicus) occurred only in the native vegetation. Its absence from the plantation vegetation might be related to the lack of native trees where this species could perch and forage (Ojeda & Chazarreta, 2014). Unlike native trees, E. globulus are devoid of natural cavities used for C. magellanicus for nesting, and provide little food (i.e., insects) supply. Therefore, as abandoned Eucalyptus plantations do not provide complementary habitat conditions that would increase beta diversity at the landscape scale, we recommend to progressively crop the plantations to allow native vegetation restoration. This approach is currently being conducted at the Valdivian Coastal Reserve as a small-scale experimental procedure (Lara, Little, González, & Lobos, 2013). This should be accompanied with further studies considering a larger sample size and covering a larger extent.
C. B. de Fontúrbel assisted in the field. Comments of 2 anonymous improved an earlier version. We thank The Nature Conservancy and the Valdivian Coastal Reserve for granting access permissions and providing field facilities. Comments of 2 anonymous reviewers improved an early version of this manuscript. Funding was provided by the Rufford Small Grants Foundation (10621-1) and the Chilean Commission for Scientific and Technological Research (CONICYT; AT-24121082). GJCV was supported by Vicerrectoría de Investigaciones y Posgrados- Universidad de Caldas. The final writing of this article was supported by FONDECYT project 3140528 (FEF).
Peer Review under the responsibility of Universidad Nacional Autónoma de México.