






Los bosques del norte de Uruguay sufren de procesos de transformación del territorio y degradación, actualmente existe escasa información sobre la estructura y composición florística de las comunidades arbóreas que sirva como base para estudiar su dinámica. Se analizó la distribución, la composición arbórea y el manejo silvícola de los bosques de la región de Rivera, Uruguay. El área de estudio comprende 33 000 ha delimitadas mediante 2 criterios, uno natural (cuenca hidrológica) y otro socioeconómico (ciudad de referencia). La información proveniente de inventarios florísticos y encuestas a productores fue ingresada a un sistema de información geográfica. Los bosques se agruparon de acuerdo al gradiente hídrico asociado con curvas de nivel. La superficie de los bosques ocupa el 13.9% del área de estudio (región de Rivera), cifra que triplica la superficie de bosques existente a nivel nacional. Se registraron 60 especies leñosas. Las familias con mayor frecuencia fueron Anacardiaceae (26.5%) y Euphorbiaceae (14.7%). Las Anacardiaceae son más frecuentes en los bosques subxerófilos y las Euphorbiaceae en los bosques hidrófilos. Todos los bosques son vulnerables debido a la mayor importancia que se concede a la ganadería, sin que exista un manejo integrado de los bosques que contemple la multifuncionalidad de usos (silvicultura y ganadería). Los bosques cercanos a la ciudad están más degradados debido a la presencia de especies exóticas.
The northern region of Uruguay undergoes land transformation processes, and degradation of native forest. The existing information on the structure and floristic composition of the forests in that region, as a basis for studies about dynamics, is scarce. Species distribution, floristic composition (wooden species) and forest management, was assessed in the Rivera Region, Uruguay. The study area (33 000 ha) was delimited by 2 simultaneous and complementary approaches, the first one natural (basin) and the other one socioeconomic (reference city). Data from floristic inventories and surveys of producers were captured into a geographic information system. Forests were grouped according to their water gradient associated to contour lines. Native forest area occupies 13.9% of the total study area in Rivera, which means 3 times the national average. Sixty woody species were found, with the Anacardiaceae (26.5%) and Euphorbiaceae (14.7%) as the most frequent families. Anacardiaceae are more common in subxerophytic woods, and Euphorbiaceae in more hydrophilic forests. All forests are vulnerable due to the weak integration between forestry and farming, resulting in no forest management. Non native species constitute a source of degradation of native forests, specifically in areas close to the city.
La transformación de la cubierta vegetal del norte de Uruguay data desde la colonización española y el establecimiento de los fazendeiros portugueses que sumaron su influencia luso-brasilera en la región (Perazza, 1998). La presión antropogénica de varios siglos de pastoreo del ganado bovino, más de una centuria de ganado ovino e introducción de especies forrajeras (Leite, 2002) se incrementó en el siglo XIX con la llegada masiva de europeos que sumaron a los impactos de la colonización primaria, impactos en la vegetación forestal de la región (Quadros y Pillar, 2002). Para el año 2004 la superficie forestada con especies introducidas de pinos y eucaliptos en la cuenca alta del río Tacuarembó era de 21% (Dell’Acqua et al., 2009). En este sentido la tendencia hacia ambientes con menor riqueza y mayor homogeneidad de especies ocurrida en el último siglo, ha sido provocada por la explotación humana, el pastoreo, la agricultura y la urbanización, factores que han sido definidos a nivel mundial (Riis y Sand-Jensen, 2001; Celesti-Grapow et al., 2006; Tassin et al., 2006). La vegetación de Uruguay pertenece al distrito uruguayense de la Provincia Pampeana, no obstante las comunidades norteñas tienen similitudes con la Provincia Paranaense y con la región del Cerrado del centro de Brasil (Cabrera y Willink, 1973; Grela, 2003) y con la del Espinal y Atlántica (Arballo, 2011). La flora está representada por numerosas especies, debido al hecho de ser una región transicional (Sganga, 1994; Arballo, 2011). Sin embargo, en Uruguay son escasos los estudios que permitan comparar el estado actual de las comunidades vegetales en relación con la situación original, así como de su dinámica tanto en su condición natural como bajo manejo silvícola (Grela, 2003). En concreto, entre los trabajos florísticos sobre bosques de la región norte del país se encuentran: el estudio en las nacientes del arroyo Lunarejo (Brussa y Escudero, 1986), la evaluación de un bosque subtropical de quebrada (Grela, 2003) y la caracterización del bosque fluvial de Minas de Corrales (Piaggio y Delfino, 2009).
Los bosques del norte del Uruguay presentan una mayor riqueza de especies debido a los aportes específicos del sur de Brasil (Brussa y Grela, 2007; Piaggio y Delfino, 2009), por ello se identificó a la naciente de la cuenca superior del río Tacuarembó localizada en el Departamento de Rivera como representativa de esa biodiversidad florística ya que incluye la vegetación de quebradas al oeste de Rivera (Grela, 2003; Brussa y Grela, 2007) y por estar enclavada en la interfase de las 2 formaciones geológicas más importantes del norte de Uruguay (Bossi, 1975).
Los ecosistemas boscosos de Uruguay han sido categorizados según criterios fisonómicos y los factores que determinan su heterogeneidad no se comprenden con claridad (Gautreau y Lezama, 2009). Las especies han sido clasificadas como subxerófilas, mesófilas e hidrófilas dado que se distribuyen de acuerdo a sus requerimientos hídricos (Piaggio y Delfino, 2009). No obstante, entre varias interrogantes, los bosques en los que se desarrollan esas especies no han sido agrupados a partir de un criterio riguroso que vincule tal categorización con el gradiente hídrico del suelo asociado a las curvas del nivel del terreno.
Se considera que los principales factores que inciden sobre la composición y la distribución arbórea del bosque son el gradiente hídrico del suelo dado por la duración anual de las inundaciones y las interferencias humanas (urbanización, tala de bosques e introducción de especies invasoras) (Lavoie et al., 2003; Wittmann et al., 2006; Toledo, 2008). Ambos factores conformarían lo que se podría denominar un modelo hidro-antrópico para la interpretación de la vegetación regional.
A través de los sistemas de información geográfica (SIG), es posible diagnosticar el estado actual de los recursos naturales y realizar evaluaciones y monitoreo sobre su dinámica de la cuenca superior del Tacuarembó, a efecto de desarrollar mecanismos efectivos para la toma de decisiones relativas al manejo, conservación y uso sostenible (MGAP, 2004; Salcedo et al., 2009). En contraparte a esta visión cuantitativa, sin una investigación etnobotánica cualitativa se pierden muchas informaciones importantes que no son reveladas por los números (Albuquerque, 2005) y que están vinculadas con el conocimiento que los propietarios de los bosques tienen sobre los recursos que aprovechan y manejan.
Con base en lo expuesto, el objetivo general es conocer la composición florística arbórea, evaluar la distribución de especies y el tipo de manejo silvícola en los bosques nativos de la cuenca superior de río Tacuarembó, el objetivo específico es definir una metodología de inventario a otras zonas del país.
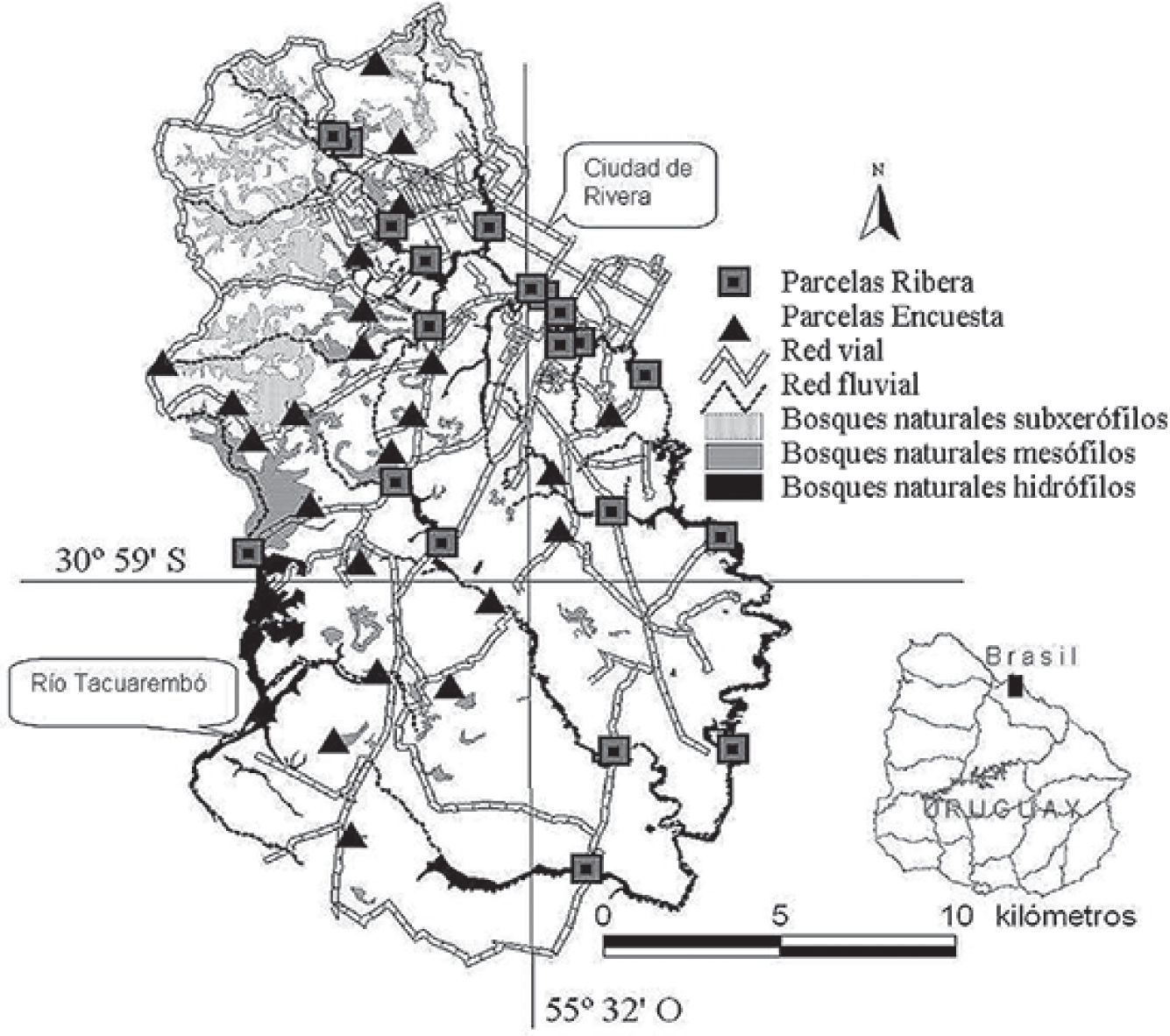
Materiales y métodosPara delimitar el área de estudio se estableció en primer lugar la cuenca hidrológica como unidad geográfica de análisis territorial (Lanna, 2000) y se eligió la cuenca del río Tacuarembó, situada en el Departamento de Rivera al norte de Uruguay. Dentro de ella se delimitó el área de trabajo a la parte superior de la cuenca, con un superficie de 33 000 ha. Este territorio se localiza en las nacientes del río Tacuarembó a una distancia de 25km de la ciudad de Rivera. El límite norte de la cuenca es la frontera con Brasil y el arroyo Cuñapirú que atraviesa la ciudad de Rivera (Fig. 1).

La región se ubica en un área de frontera con Brasil que posee características biogeográficas similares (Chebataroff, 1969). De acuerdo con la clasificación climática de Köppen (Strahler y Strahler, 1997) el clima es del tipo Cfa, templado y húmedo con precipitaciones todo el año y temperatura del mes más cálido superior a 22° C. La temperatura media anual es de 18.1° C y la precipitación anual acumulada promedio es de 1 639mm (DNM, 2009). El relieve de colinas y lomeríos desarrolla cursos de agua que pertenecen a la cuenca del río Uruguay dentro de un territorio en el que se aprecian 2 unidades geomorfológicas dominantes: la cuesta basáltica y la cuenca sedimentaria del noreste. La primera producida por el ascenso de masas de lavas de edad cretácica y la segunda compuesta por sedimentos gondwánicos que van desde el devoniano hasta el triásico (Panario, 1988). El material geológico dominante se corresponde con rocas basálticas de la formación geológica de Arapey y con rocas areniscas de la formación geológica Tacuarembó (Bossi, 1975) cuyos suelos dominantes son los litosoles, con un espesor menor a 30cm y propiedades físicoquímicas variadas y los acrisoles, que son suelos profundos de texturas livianas, lixiviadas y con un elevado nivel de aluminio (Durán, 1991).
Para el tamaño de la muestra óptimo se fijó la expresión establecida por Walpole y Myers (1998):
donde n es el tamaño de la muestra, Z el valor de tabla normal estándar para un nivel de significación, 8 es el desvío estándar y e es el error en proporción prefijado. A su vez, se consideró que la teoría de probabilidades a través de la ley de los grandes números de Bernoulli, establece que el valor medio observado en n ensayos (muestras), converge para E(X) (esperanza matemática) cuando n — x> (Infante y Zárate-de Lara, 1994). Por último, se atendió a Costa y Araújo (2001) quienes sugieren parcelas cuadradas de 400 m2. En función de lo expuesto, se llegó a la necesidad de fijar 44 parcelas (20 de ribera y 24 encuesta). Las parcelas de ribera correspondientes a lugares de fácil acceso, localizados en mapa como puntos de intercepción de cursos de aguas (red fluvial) con caminos (red vial) y las parcelas encuesta que a partir de un mapa trazado se ubicaron aleatoriamente in situ por medio de un recorrido por la zona, buscando áreas con asentamientos humanos en explotaciones agroforestales.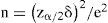
La toma de datos se realizó entre noviembre de 2006 y julio de 2008. A partir de la fotointerpretación de imágenes satelitales de Google Earth de alta resolución (2006) se localizaron los rodales de bosques nativos. Posteriormente, en el programa Arc-Gis 9.2 se generó un sistema de información geográfica (en lo sucesivo SIG), en donde se georeferenciaron las imágenes satelitales y se trazaron polígonos sobre los bosques naturales.
Posteriormente se procedió al muestreo de las parcelas de ribera. De estos puntos se eligieron de forma aleatoria 20 para proceder al inventario de los bosques, que se realizó avanzando 100 m en la dirección ascendente de la ribera para evitar el efecto antrópico de los cruces con los caminos. La parcela fue georeferenciada con sistema de posicionamiento global (GPS) en coordenadas planas Yacaré (sistema Uruguay), se registró: latitud, longitud, altitud y distancia a la ciudad de Rivera. Se censaron los árboles de todas las especies leñosas cuyos tallos superaron los 10cm de DAP (diámetro a la altura del pecho) (Blanc et al., 2000). Se registró el nombre de la especie y se tomaron muestras botánicas para su posterior determinación en el Herbario del Jardín Botánico de la Intendencia Municipal de Montevideo (Matías y Nunes, 2001). Después de identificadas las muestras, todos los nombres fueron verificados en IPNI (2011).
Para clasificar los bosques se consideró las especies hidrófilas que crecen en contacto directo con el agua o en zonas cercanas. En zonas más o menos húmedas se encuentran las mesófilas, por último hacia las afueras o en zonas de serranías con alto grado de rocosidad y en suelos poco profundos aparecen las especies subxerófilas. Asimismo, se consideró la presencia de especies bioindicadoras, de acuerdo a los criterios de Brussa y Grela (2007) y Piaggio y Delfino (2009). La información altimétrica de las curvas de nivel de 10 m creada por SGM (2000) se incluyó en el SIG. Si en una parcela la mayor frecuencia relativa se correspondía con especies hidrófilas la parcela se consideró como bosque hidrófilo. Luego, se ordenaron todas las parcelas de acuerdo a su categoría y a la cota altitudinal, para después en el SIG, unir, dividir e interpolar polígonos (bosques) en función del tipo de parcela hallada y las curvas de nivel correspondientes. Los bosques se agruparon en: bosques hidrófilos, localizados entre 160-210 m; bosques mesófilos, posicionados entre 210- 260 m y bosques subxerófilos ubicados en sitios superiores a 260 m.
Para cada grupo de bosque fue calculada la densidad de árboles, el total de especies y el índice de diversidad de Shannon-Wiener (Hastings y Beattie, 2006). La información sobre la composición de especies fue agrupada en las familias botánicas respectivas a las especies presentes y de acuerdo al origen geográfico (nativas e introducidas) (Backes y Nardino, 2001). Se calcularon el coeficiente de variación (CV), frecuencia relativa, frecuencia relativa acumulada y densidad de las especies. Para evaluar la posible significancia en la localización de las familias botánicas se efectuó el análisis de la varianza (ANOVA) (Infante y Zárate-de Lara, 1994), creándose una base de datos de presencia-ausencia de familias, con 3 tratamientos por familia (subxerófilos, mesófilos e hidrófilos). Con el fin de detectar la significancia estadística en la distribución de las especies, según su origen geográfico, se establecieron 2 categorías, nativas (N) e introducidas (I), en columnas que incluyen las distancias en metros de todos los árboles a la ciudad de Rivera.
En cada parcela se calculó el índice de Diversidad Simpson (Smith y Smith, 2001) que estima la probabilidad (valores 0-1), de que 2 individuos de una parcela, sean de especies diferentes cuando se practican 2 extracciones sucesivas sin reposición,
donde ni es el número de individuos de la especie i de la parcela y N el número total de individuos de la parcela.
Dado que el índice anterior no discrimina entre la diversidad nativa y la exótica, se propuso un nuevo índice denominado de “Naturalidad” que estima la probabilidad (valores 0-1) de que al tomar 2 individuos, sean ambos de especies nativas, cuando se practican 2 extracciones sucesivas sin reposición; para ello se usó la expresión:
donde h es el número de individuos arbóreos de las especies nativas en la parcela y N es el número total de individuos arbóreos de la parcela. Por último, para estimar la diversidad de origen natural de cada parcela, quedó justificada la proposición de un índice nuevo denominado de “Diversidad Natural” (DN’) que mide la probabilidad de que al tomar 2 individuos al azar en 2 extracciones sin reposición, sean de 2 especies nativas diferentes. Este índice surge como el producto de las 2 probabilidades anteriores (valores 0-1), bajo un supuesto estadístico de independencia de eventos, la expresión es la siguiente:

Para los 3 índices se probaron las regresiones de mejor ajuste, en el programa computacional Curve Expert.
Para las parcelas encuesta se planificó un recorrido que abarcó entrevistas a 24 productores mediante la aplicación de un cuestionario (Salcedo et al., 2009) que reveló información productiva y etnobotánica (Madiwalar et al., 2007; Camargo-de Oliveira et al., 2009). A cada productor se le preguntó por el régimen de propiedad, el tipo de sistema silvícola ejecutado: aclareos, podas, plantaciones, extracción de leña y tala de árboles. En el caso de extraer madera del bosque se le interrogó por las toneladas cosechadas por hectárea y por año. Por último, se preguntó a los productores qué especies de árboles nativos existían y reconocían en sus predios; para ésto, se registraron los nombres comunes de los árboles. Posteriormente, se escogió de forma aleatoria un rodal de bosque nativo incluido en la propiedad del encuestado, luego se caminó 40 m hacia el interior, para evitar el efecto de borde y ubicar la parcela. En las parcelas se recabó la información florística de la misma manera que en el muestreo de las parcelas ribera.
ResultadosDistribución y composición de los bosques. De acuerdo al SIG, la superficie de los bosques totalizó 4 573.5 ha lo que equivale al 13.8% de la superficie de la cuenca superior del río, el resto corresponde a campos o praderas. Los bosques se distribuyen en altitudes promedio de 224 m (± 46.7, desviación estándar). Se contabilizaron 184 rodales que tuvieron un promedio de 24.8 ha (± 63.5). Los bosques subxerófilos de las zonas altas tienen un área de 1 578.4 ha; los mesófilos, ubicados en las laderas medias, una superficie de 1 606.3 ha y los hidrófilos, localizados en las zonas más húmedas del sur, totalizaron 1 388.8 ha. Los subxerófilos están formados por árboles más bajos, espinosos o con ramas agudo espinosas, los hidrófilos presentan árboles con fuste recto, en tanto los mesófilos presentaron características intermedias.
La densidad media fue de 752 árboles/ha (CV= 0.86). En los bosques subxerófilos la cifra sube a 821 árboles/ha (CV= 1.02), en los mesófilos es de 954 árboles/ha (CV= 0.92) y en los hidrófilos se encontró la menor densidad con 567 individuos/ha (CV= 0.75). Fueron halladas 24 especies en los bosques subxerófilos, 49 en los mesófilos y 40 en los hidrófilos, los índices de diversidad Shannon- Wiener respectivos fueron: 2.65, 3.06 y 2.92 (Cuadro 1).
Total de especies leñosas, densidad total, densidad por especie e índice de Shannon-Wiener por tipo de bosque nativo de la cuenca superior del río Tacuarembó, Uruguay. nd, no determinado (especies que no determinaron valores de densidad en los análisis)
| Parámetro | Bosques hidrófilos | Bosques mesófilos | Bosques subxerófilos |
|---|---|---|---|
| Total de especies | 40 | 49 | 24 |
| Densidad (árboles/ha) | 567 | 954 | 821 |
| Índice de Shannon-Wiener | 2.92 | 3.06 | 2.65 |
| Especie | Densidad | específica | (árboles/ha) |
| Sebastiania commersoniana (Baill.) L.B. Sm. et R.J. Downs | 96 | 37 | 50 |
| Sebastiania brasiliensis Spreng. | 84 | 9 | nd |
| Gleditsia triacanthos L | 58 | 1 | nd |
| Ocotea acutifolia (Nees) Mez | 35 | 32 | 33 |
| Scutia buxifolia Reissek | 32 | 57 | 17 |
| Eugenia uniflora L. | 31 | 84 | nd |
| Allophylus edulis Niederl. | 29 | 46 | 54 |
| Salix humboldtiana Willd. | 25 | 1 | nd |
| Lithraea molleoides (Vell.) Engl. | 20 | 163 | 138 |
| Erythrina crista-galli L. | 20 | 3 | nd |
| Calliandra tweediei Benth. | 14 | 6 | nd |
| Pouteria salicifolia Radlk. | 14 | 7 | nd |
| Ligustrum lucidum W.T.Aiton | 12 | 7 | nd |
| Styrax leprosus Hook. et Arn. | 8 | 25 | 54 |
| Guettarda uruguensis Cham. et Schltdl. | 8 | 6 | 4 |
| Ligustrum sinense Lour. | 8 | nd | nd |
| Myrrhinium atropurpureum Schott | 7 | 26 | 29 |
| Blepharocalyx salicifolius (Kunth) O. Berg | 7 | 6 | 17 |
| Prunus subcoriacea Koehne | 7 | 4 | nd |
| Schinus longifolius Speg | 7 | 68 | 38 |
| Escallonia bifida Link et Otto ex Engl. | 5 | nd | nd |
| Eucalyptus grandis W. Hill | 5 | nd | nd |
| Aloysia gratissima (Gillies et Hook.) Tronc. | 4 | 26 | nd |
| Salix alba Thunb. | 4 | nd | nd |
| Celtis tala Gillies ex Planch. | 3 | 3 | 8 |
| Myrceugenia euosma (O. Berg) D. Legrand | 3 | 6 | 4 |
| Myrcianthes cisplatensis O. Berg | 2 | 3 | nd |
| Luehea divaricata Mart. | 2 | nd | nd |
| Acca sellowiana (O. Berg) Burret | 2 | 21 | 13 |
| Myrsine coriacea R. Br. | 2 | 4 | 4 |
| Zanthoxylum hyemale A. St.-Hil. | 2 | 26 | 29 |
| Myrsine laetevirens Arechav. | 1 | 2 | 4 |
| Morus alba L. | 1 | nd | nd |
| Daphnopsis racemosa Griseb. | 1 | 12 | nd |
| Acanthosyris spinescens Griseb. | 1 | nd | nd |
| Acacia caven (Molina) Molina | 1 | nd | nd |
| Platanus x acerifolia (Aiton) Willd. | 1 | nd | nd |
| Symplocos uniflora Bedd. | 1 | 1 | nd |
| Xylosma warburgii Briq. | 1 | 12 | 21 |
| Parkinsonia aculeata L. | 1 | nd | nd |
| Schinus molle Hort. ex Engl. | nd | 6 | 8 |
| Schinus lentiscifolia Marchand | nd | 117 | 179 |
| Robinia pseudoacacia L. | nd | 1 | nd |
| Quillaja brasiliensis Mart. | nd | 50 | 33 |
| Phytolacca dioica L. | nd | 6 | 4 |
| Nectandra megapotamica Mez | nd | 1 | nd |
| Myrcianthes pungens (O. Berg) D. Legrand | nd | 1 | nd |
| Melia azedarach L. | nd | 1 | nd |
| Maytenus ilicifolia Mart. Ex Reissek | nd | 6 | 4 |
| Heterothalamus alienus Kuntze | nd | 1 | nd |
| Gochnatia polymorpha (Less.) Cabrera | nd | 15 | nd |
| Eucalyptus tereticornis Sm. | nd | 9 | nd |
| Escallonia megapotamica Spreng. | nd | 6 | nd |
| Cupania vernalis Cambess. | nd | 4 | 42 |
| Citronella gongonha (Mart.) R.A. Howard | nd | 1 | nd |
| Citharexylum montevidense Moldenke | nd | nd | 33 |
| Buddleja grandiflora Cham. et Schltdl. | nd | 15 | nd |
| Berberis laurina Thunb. | nd | 1 | nd |
| Acer negundo L. | nd | 1 | nd |
| Acacia decurrens Willd. | nd | 1 | nd |
Los bosques están integrados por 60 especies principalmente nativas. Las nativas constituyen el 93% de los bosques y las introducidas el 7%. Esta riqueza arbórea se agrupa en 33 familias, de las cuales las de mayor frecuencia son Anacardiaceae (26.5%), Euphorbiaceae (14.7%) y Myrtaceae (12.9%) (Cuadro 2).
Frecuencia relativa de las especies leñosas del bosque nativo de la cuenca superior del río Tacuarembó, Uruguay. Lista ordenada de mayor a menor de acuerdo a los valores de frecuencia. f(i)= frecuencia relativa en porcentaje. *Especies introducidas a la región, cuya frecuencia relativa acumulada es de 6.9%
| Especie | Familia | f(i) |
|---|---|---|
| Lithraea molleoides (Vell.) Engl. | Anacardiaceae | 12.18 |
| Schinus lentiscifolia Marchand | Anacardiaceae | 9.27 |
| Sebastiania commersoniana (Baill.) L.B. Sm. et R.J. Downs | Euphorbiaceae | 8.93 |
| Eugenia uniflora L. | Myrtaceae | 6.31 |
| Sebastiania brasiliensis Spreng. | Euphorbiaceae | 5.76 |
| Scutia buxifolia Reissek | Rhamnaceae | 5.24 |
| Allophylus edulis Niederl. | Sapindaceae | 5.21 |
| Schinus longifolius Speg | Anacardiaceae | 4.61 |
| Ocotea acutifolia (Nees) Mez | Lauraceae | 4.52 |
| Gleditsia triacanthos L* | Fabaceae | 3.74 |
| Quillaja brasiliensis Mart. | Quillajaceae | 3.18 |
| Styrax leprosus Hook. et Arn. | Styracaceae | 2.80 |
| Myrrhinium atropurpureum Schott | Myrtaceae | 2.35 |
| Zanthoxylum hyemale A. St.-Hil. | Rutaceae | 1.97 |
| Salix humboldtiana Willd. | Salicaceae | 1.63 |
| Aloysia gratissima (Gillies et Hook.) Tronc. | Verbenaceae | 1.63 |
| Acca sellowiana (O. Berg) Burret | Myrtaceae | 1.48 |
| Erythrina crista-galli L. | Fabaceae | 1,40 |
| Pouteria salicifolia Radlk. | Sapotaceae | 1.28 |
| Calliandra tweediei Benth. | Fabaceae | 1.22 |
| Ligustrum lucidum W.T. Aiton* | Oleaceae | 1.12 |
| Blepharocalyx salicifolius (Kunth) O. Berg | Myrtaceae | 1.06 |
| Xylosma warburgii Briq. | Flacourtiaceae | 1.03 |
| Cupania vernalis Cambess. | Sapindaceae | 0.98 |
| Guettarda uruguensis Cham. et Schltdl. | Rubiaceae | 0.87 |
| Gochnatia polymorpha (Less.) Cabrera | Asteraceae | 0.76 |
| Buddleja grandiflora Cham. et Schltdl. | Buddlejaceae | 0.76 |
| Daphnopsis racemosa Griseb. | Thymelacaceae | 0.68 |
| Prunus subcoriacea Koehne | Rosaceae | 0.65 |
| Citharexylum montevidense Moldenke | Verbenaceae | 0.61 |
| Myrceugenia euosma (O.Berg) D. Legrand | Myrtaceae | 0.56 |
| Ligustrum sinense Lour.* | Oleaceae | 0.50 |
| Celtis tala Gillies ex Planch. | Ulmaceae | 0.49 |
| Schinus molle Hort. ex Engl. | Anacardiaceae | 0.45 |
| Eucalyptus tereticornis Sm.* | Myrtaceae | 0.45 |
| Myrsine coriacea R. Br. | Myrsinaceae | 0.43 |
| Maytenus ilicifolia Mart. Ex Reissek | Celastraceae | 0.38 |
| Phytolacca dioica L. | Phytolaccaceae | 0.38 |
| Escallonia bifida Link et Otto ex Engl. | Saxifragaceae | 0.35 |
| Eucalyptus grandis W. Hill* | Myrtaceae | 0.34 |
| Myrcianthes cisplatensis O. Berg | Myrtaceae | 0.32 |
| Escallonia megapotamica Spreng. | Saxifragaceae | 0.31 |
| Myrsine laetevirens Arechav. | Myrsinaceae | 0.26 |
| Salix alba Thunb.* | Salicaceae | 0.25 |
| Luehea divaricata Mart. | Tiliaceae | 0.15 |
| Symplocos uniflora Bedd. | Symplocacaceae | 0.14 |
| Acer negundo L.* | Aceraceae | 0.09 |
| Heterothalamus alienus Kuntze | Asteraceae | 0.09 |
| Berberis laurina Thunb. | Berberidaceae | 0.08 |
| Acacia caven (Molina) Molina | Fabaceae | 0.08 |
| Acacia decurrens Willd.* | Fabaceae | 0.08 |
| Robinia pseudoacacia L.* | Fabaceae | 0.08 |
| Citronella gongonha (Mart.) R.A. Howard | Icacinaceae | 0.08 |
| Nectandra megapotamica Mez | Lauraceae | 0.07 |
| Melia azedarach L.* | Meliaceae | 0.07 |
| Morus alba L.* | Moraceae | 0.07 |
| Myrcianthes pungens (O. Berg) D. Legrand | Myrtaceae | 0.06 |
| Acanthosyris spinescens Griseb. | Santalaceae | 0.06 |
| Platanus x acerifolia (Aiton) Willd.* | Platanaceae | 0.05 |
| Parkinsonia aculeata L. | Fabaceae | 0.05 |
La familia Anacardiaceae, representada por los géneros Schinus y Lithraea, tiene mayor frecuencia en los bosques subxerófilos ubicados al noroeste del área que en el resto de los bosques (F= 3.19; p= 0.044). En esos lugares se presenta mayor altitud y suelos poco profundos y de origen basáltico. Cuando esas zonas altas coincidieron con los cursos de agua fueron observadas laderas abruptas con mayor humedad y temperatura que permite el desarrollo de pteridophytas y briophytas. Por su parte, la frecuencia de la familia Myrtaceae, representada por Eugenia uniflora L. (Pitanga) y Myrrhinium atropurpureum Schott (palo de fierro), no presentó diferencias significativas entre los diferentes tipos de bosques (F= 1.65; p= 0.19). La familia Euphorbiaceae, constituida por el género Sebastiania (Blanquillos), tiene mayor frecuencia en los bosques fluviales de tipo hidrófilos (F= 3.42; p= 0.035).
Las especies Salix humboldtiana Willd. (sauce criollo) y Erythrina crista-galli L. (ceibo) crecen en bosques hidrófilos de la cuenca superior del río Tacuarembó, mientras que al inicio de los bosques mesófilos (cota de 222 m snm), se encontró un representante de cada especie y ninguno en los bosques subxerófilos. En ambientes hidrófilos también crecen: Pouteria salicifolia Radlk. (mataojo) y Escallonia bifida Link et Otto ex Engl. (árbol del pito), las cuales no superan las cotas de 212 y 210 m snm.
En áreas cercanas a la ciudad de Rivera se han establecido especies introducidas de carácter invasor, entre las que se destacan: Gleditsia triacanthos L, Ligustrum lucidum W. T. Aiton y Ligustrum sinense Lour. Si se escoge un árbol exótico al azar, éste se ubica a una distancia de 5.72 (± 3.95) m de la ciudad de Rivera, mientras que un árbol nativo se encuentra a 10.11 (± 4.55) m de la misma ciudad. El ANOVA reflejó que las diferencias fueron altamente significativas (F= 29.82; p< 0.0001). Las introducidas se diseminaron, porque en algunos casos las fronteras de fragmentación de los bosques fueron difusas, debido a las interconexiones que los rodales tuvieron en los corredores fluviales más allá de los límites prediales.
Las regresiones de la funciones de densidad de probabilidad de diversidad Simpson, índice de naturalidad y diversidad natural en función de la distancia (m) a la ciudad de Rivera son presentadas en la figura 2. De todos los modelos matemáticos posibles, los obtenidos a continuación fueron los que proyectaron valores de probabilidad entre 0 y 1 para todo el recorrido de cada una de las 3 variables ecológicas. Además, fueron los más simples por presentar: un menor número de coeficientes, un alto coeficiente de correlación y un bajo error estándar. Para la diversidad Simpson, de 7 modelos posibles, la mejor regresión obtenida fue la propuesta por el modelo sigmoidal:


(a = -2.24E-02, b = 3.29E+04, c = 8.80E-01 y d = 1.48E-00), el cual presenta un coeficiente de correlación 0.78 y un desvío estándar de 0.11. Para el índice de naturalidad, de 8 modelos ensayados, el mejor modelo de regresión es de asociación exponencial:

(a = 1.08E+00 y b = 2.03E-04) con un coeficiente de correlación de 0.82 y un desvío estándar de 0.19. Por último, entre 8 modelos posibles, la mejor regresión del índice de diversidad naturalidad es:
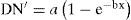
(a = 8.82E-01 y b = 2.23E-04) determinada también por el modelo de asociación exponencial, con un coeficiente de correlación de 0.76 y un desvío estándar de 0.20. Manejo de los bosques. En el 95% de los casos la tenencia de la tierra de la población entrevistada estuvo en manos de propietarios familiares. Los productores con proyectos de manejo silvícola elaborado por profesionales fueron el 18%. Dado que la riqueza derivada del muestreo de encuesta fue de 10.0 (± 3.0) especies, se comprobó un conocimiento dendrológico restringido por parte de los propietarios, quienes sólo reconocieron en su predio 4.3 (± 2.3) especies. En cuanto a tipo de prácticas silvícolas la figura 3 muestra las frecuencias relativas.

El corte de madera de los bosques tuvo como objetivo el uso para combustible o leña por la población local. Para tal motivo se cosechan 12.3 (±26.5) toneladas de leña por ha y por año. A su vez, el promedio de madera vendida por predio es de 5.5 (±15.0) toneladas por ha al año; sin embargo, sólo el 59% de los productores utiliza la madera del bosque natural y sólo el 9% vendió la leña cosechada.
Por último, en el área se observó la fabricación de ladrillos, actividad que implica la remoción del suelo y ocasiona el impacto a los bosques y árboles, en especial en las zonas aledañas a la ciudad de Rivera, los cuales son enviados como leña a la misma ciudad. Si bien hasta el momento ese disturbio abarca sólo 10 ha, es posible que en el corto plazo se incremente esta actividad y ocasione un mayor daño a los bosques. De igual forma se observa que los asentamientos humanos de Rivera han reducido y fragmentado el área boscosa. Según Sganga (1994) en el bosque fluvial, los árboles eran fustales (pies adultos con un sólo fuste y en general procedentes de regeneración por semilla), pero en la actualidad el talado los ha llevado al régimen tallar (árboles en general con más de un fuste, resultado de su regeneración por brotes de cepa posteriores a la corta).
DiscusiónLos bosques nativos que actualmente abarcan el 3% de la superficie de Uruguay, tal vez no cubrieron más del 5% del país (Chebataroff, 1969; Alonso-Paz y Bassagoda, 2002), pero al menos la cuenca de la laguna Merín (van Dillewijn, 1968) y el valle del río Uruguay (Sganga y Panario, 1974) estuvieron cubiertos por una importante vegetación boscosa que debe haber superado esas cifras. Sin embargo, en la cuenca superior izquierda del río Tacuarembó (33 000 ha) los bosques ocupan 4 574 ha, lo que supone el 13.9% del territorio, triplicando así el porcentaje nacional.
Arballo y Cravino (1999) diferencian 4 asociaciones vegetales para los distintos tipos de bosque en Uruguay, que pueden asimilarse a las distintas formaciones encontradas en la cuenca del Tacuarembó. En primer lugar, el bosque ribereño, comparable a los bosques hidrófilos de este estudio; la selva de quebrada subtropical, que se ha encontrado en áreas más reducidas de la zona estudiada; la sabana esteparia serrana, compuesta por una cubierta arbórea que puede equipararse a los bosques mesófilos, de este trabajo; y la asociación arbustiva semixerófila, que se inventarió en zonas de campos o pradera manchadas de bosquetes.
Para el bosque fluvial 13 géneros reportados por Sganga (1994) son coincidentes con éste estudio. Brussa y Escudero (1986) en sus trabajos florísticos en el arroyo Lunarejo, afluente del Tacuarembó, diferenciaron bosques de valles, bosques de quebrada del cauce, bosques de quebrada de la cumbre y bosque de parque. Esa clasificación implica un muestreo de la transición desde las zonas hidrófilas hasta las subxerófilas como la propuesta en ésta investigación. En cuanto a la riqueza florística se han recogido en los inventarios de campo 28 especies que coinciden con las reportadas por Brussa y Escudero (1986). También, 18 especies leñosas inventariadas en nuestro trabajo coinciden con las citadas por Grela (2003). De ellas, 8 se presentan en los 3 tipos de bosques, 5 en los bosques hidrófilos y mesófilos, 2 en los mesófilos, 2 en los mesófilos y subxerófilos y solamente 1 en los mesófilos. La palmera Syagrus romanzoffiana (Cham.) Glassman reportada por Grela (2003), también se halla en el área de éste estudio; sin embargo, no forma parte del listado pues no se trata de una leñosa. Por último, son compartidas 22 especies con el estudio florístico del arroyo Corrales (afluente indirecto del río Tacuarembó) realizado por Piaggio y Delfino (2009), al igual que en este trabajo, la máxima riqueza específica de nativas encontrada es para la familia Myrtaceae (7).
Los géneros Schinus y Lithraea, son más abundantes en los bosques subxerófilos de las zonas altas, donde el suelo es poco profundo y de naturaleza basáltica, aunque dentro de la familia esta tendencia es más marcada para Schinus lentiscifolia y Lithraea molleoides cuyas densidades son mayores en esos bosques. Este hallazgo coincide con lo observado por Brussa y Escudero (1986) en las zonas más elevadas de la cuenca del arroyo Lunarejo. Por su parte, los registros de Eugenia uniflora L. (Pitanga) datan del año 1817, cuando el naturalista Dámaso Antonio Larrañaga la reporta como una planta de fruto rojo común, dulce y con una baya de 4 surcos (Instituto Histórico y Geográfico del Uruguay, 1992). En nuestro trabajo, esta especie se considera mesófila por observarse a una distancia media del curso de agua del bosque fluvial (Brussa y Escudero, 1986; Piaggio y Delfino, 2009) y porque sus densidades respectivas fueron 31, 84, y 0 árboles/ha para bosques hidrófilos, mesófilos y subxerófilos, respectivamente.
Las especies del género Sebastiania (blanquillos) que tienen mayor frecuencia en los bosques fluviales hidrófilos son también consideradas hidrófilas por Brussa y Escudero (1986) por estar en las zonas más húmedas del valle del arroyo Lunarejo, aunque Piaggio y Delfino (2009) las consideran como mesófilas debido a que crecen en las zonas intermedias del bosque fluvial del arroyo Corrales. Con estos últimos autores se comparte el hecho de estar entre las 3 primeras en importancia por su frecuencia relativa.
Las especies Salix humboltiana, Erythrina crista-galli, Pouteria salicifolia y Escallonia bifida en nuestro trabajo son consideradas hidrófilas porque apenas se hallan algunos individuos de las 3 primeras especies en las zonas más bajas de los bosques mesófilos o directamente no se hallan individuos ni en los bosques mesófilos ni en los subxerófilos; como es el caso de la cuarta. Brussa y Escudero (1986) también consideran hidrófilas las 3 primeras especies,
Alonso-Paz y Bassagoda (2002) consideran hidrófilas a Salix y Erythrina; y Piaggio y Delfino (2009)Salix y Pouteria. Las densidades (pies/ha) para los 3 tipos de bosque (hidrófilo, mesófilo y subxerófilo) fueron de 25/1/0 pies/ ha para Salix; 20/3/0 pies/ha para Erythrina; 14/7/0 pies/ha para Pouteria, y 5/0/0 pies/ha para Escallonia.
Por las propiedades de conservar el suelo, el agua y de producir materias leñosas (MGAP 1987, Ley Forestal de Uruguay, 15,939/art. N° 8), el mantenimiento de la biodiversidad de los bosques naturales de Uruguay deberá integrarse en la gestión de los sistemas productivos (Torres et al., 1993). Aunque producto de la integración se observa que en la cuenca del río Tacuarembó, la vegetación arbórea de baja densidad que se inserta en un tapiz graminoide, ha sido impactada por la ganadería de igual modo que en la zona vecina del sur del estado de Rio Grande do Sul, con la que son compartidas 5 especies arbóreas: Scutia buxifolia, Sebastiania commersoniana, Lithraea brasiliensis, Schinus lentiscifolius y Allophyllus edulis (Leite, 2000). Sin embargo, esa baja integración observada entre la silvicultura y la ganadería es relativizada por Senyszyn (1969), quien sugiere que los bosques de Uruguay tienen el cometido de proteger la actividad ganadera, aunque no está en condiciones de producir ni volúmenes, ni calidad importante de madera. Esos conceptos son cuestionados por Reitz et al. (1988), quienes basándose en las potencialidades de las propiedades físico-mecánicas de la madera y combustibles, en los servicios ecológicos y los productos medicinales e industriales de las cuales se puede obte ner; destacan géneros y especies halladas en los bosques hidrófilos, mesófilos y subxerófilos del presente estudio, como: Pouteria salicifolia, Erythrina sp., Sebastiania commersoniana, Ocotea sp., Rapanea sp., Eugenia uniflora, Allophyllus edulis, Myrcianthes pungens, Quillaja brasiliensis, Styrax leprosus, Phytolacca dioica y Schinus molle. En conclusión, deberían potenciarse tanto la integración silvicultura-ganadería como la conservación del germoplasma de buenas propiedades maderables.
La urbanización de la región ha degradado el bosque fluvial (King y Buckney, 2002; Cayuela et al., 2006), aspecto observado en las zona aledañas a la ciudad de Rivera. Con el crecimiento urbano, esos disturbios serían transmitidos a las áreas de interfase entre el medio urbano y el rural (Toledo, 2008). El potencial invasor de las especies introducidas está vinculado con la población inicial y el tiempo de residencia (Rejmánek, 2000) y a factores históricos de una introducción no intencionada (Lavoie et al., 2003).
La presencia de especies introducidas puede provocar la supresión de la regeneración nativa (Blanc et al., 2000), lo cual se observa en los lugares donde crece Ligustrum lucidum, especie que ocasionó un impacto similar en las islas del océano Índico (Tassin et al., 2006). De acuerdo con Nebel y Porcile (2009) esta especie se ha diseminado a través de los bosques nativos de Uruguay. A la luz de los resultados que demuestran diferencias altamente significativas entre las especies nativas y las introducidas, ya que las primeras se encuentran en promedio casi al doble de distancia de la ciudad de Rivera que las segundas; se sugiere la implementación de programas de manejo de las especies invasoras (Nagler et al., 2005; van Wilgen et al., 2007). Los programas deben focalizarse en los bosques hidrófilos pues ninguna especie exótica se encuentra en los bosques subxerófilos, aspecto que reafirma la influencia humana de las zonas bajas. Sumado a ello, las exóticas como grupo tienen densidades más elevadas en los bosques hidrófilos que en los mesófilos. Para el caso de las 3 especies exóticas más agresivas, las densidades para los 3 tipos de bosque hidrófilo, mesófilo y subxerófilo fueron, respectivamente: 50/1/0 pies/ha para Gleditsia triacanthos; 12/7/0 pies/ha para Ligustrum lucidum y 8/0/0 pies /ha para Ligustrum sinense. Como se trata de bosques fragmentados, su vulnerabilidad es importante y su conservación se hace difícil. La vulnerabilidad debe ser atendida en los bosques mesófilos que son los de mayor densidad de árboles y los que presentaron mayor índice de diversidad Shannon-Wiener. El índice de Simpson tiene el inconveniente de sobreestimar la biodiversidad en las zonas cercanas al centro urbano, a causa de computar la diversidad aportada por especies introducidas; la probabilidad inicial (diversidad) es de 0.63 a 2 042 m del centro urbano. A su vez, el índice de naturalidad tiene el inconveniente de llegar rápidamente al máximo de 1, a 19 132 m de la ciudad de Rivera, por el hecho de sólo atender al origen de las especies y no a la riqueza y a la equitatividad de las mismas. Por el contrario, con el nuevo índice de diversidad natural que se ha propuesto se evitan esos 2 inconvenientes, dado que este índice es de sólo 0.32 a 2 049 m y de 0.85 a 20 242 m de Rivera. Ello refleja que se trata de un bioindicador válido, que podría identificar comunidades alteradas por el hombre (Sganga, 1994) en otras regiones de Uruguay y en países con disturbios en la vegetación natural. Los bosques son hábitat de una elevada riqueza de aves silvestres (Reitz et al., 1988; Harvey y Haber, 1999). La conectividad provista por una serie de cercas vivas afecta el desplazamiento de la fauna silvestre entre hábitats naturales remanentes, facilitan la dispersión de semillas (Burel, 1996) y la regeneración de la vegetación nativa (Saunders y Hobbs, 1991). Para la obtención de certificados de sostenibilidad, la industria forestal desarrollada a partir de la forestación de praderas con especies introducidas debe evaluar los problemas ambientales, del uso de plaguicidas, el uso del agua, la flora y la fauna, los cambios de uso del suelo y los impactos sociales y económicos (Geary, 2001).
En función de lo discutido es posible establecer las siguientes conclusiones: i), la cuenca superior izquierda del río Tacuarembó tiene una proporción de bosques nativos frente al territorio total considerado, 3 veces superior a la proporción de bosques nativos existentes en Uruguay referidos a todo el territorio nacional. Sin embargo, los bosques del área estudiada son vulnerables debido a la falta de integración de la ganadería y la silvicultura en el manejo, principalmente orientado a la ganadería y con insuficiente gestión silvícola, al reducido conocimiento de los productores sobre sus bosques, a la comercialización ilegal de madera, al avance de especies invasoras, y a la remoción de suelo y la utilización de la madera como combustible. Por ello, es necesario restaurar áreas degradadas, tomar medidas que minimicen las perturbaciones y realizar nuevas evaluaciones desde las parcelas establecidas en el SIG. ii), a partir de los inventarios y los datos altimétricos se han definido 3 tipos de bosque: hidrófilo, mesófilo y subxerófilo, en los cuales el gradiente hídrico del suelo y la interferencia humana son factores determinantes de la composición y la distribución arbórea. iii), si disminuye la altitud y aumenta la disponibilidad de agua, se reduce la frecuencia de Anacardiaceae y se inicia el predominio de Euphorbiaceae. En tal sentido, Lithraea molleoides y Schinus lentiscifolia dominantes en los boques mesófilos y subxerófilos, comienzan a ser substituidas por Sebastiania commersoniana y Sebastiania brasiliensis de los bosques hidrófilos. Además, desde las zonas urbanas el grupo de las especies introducidas Gleditsia triacanthos y Ligustrum sp. desplazan a las leñosas nativas y degradan el bosque natural.
A los departamentos de Física Aplicada y de Ecología y Salud Ambiental de la Universidad de Huelva, España, en la persona de los Drs. Juan Pedro Bolívar Raya y Eloy Castellanos Verdugo por sus aportes teóricos y metodológicos para el desarrollo de ésta investigación. Al Programa de Desarrollo Tecnológico (PDT) del Ministerio de Educación y Cultura (MEC) de Uruguay, en la persona del profesor José Carrera por proporcionar fondos para el financiamiento de este estudio; al Ing. Miguel Gavirondo por sus aportes y a todos los productores visitados por su receptividad.