



Se estudiaron el ciclo de vida y la distribución temporal de Stenomacra marginella (Herrich-Schaeffer). Adultos y ninfas fueron recolectados como parte de un estudio sobre la biodiversidad de hemípteros en ambientes contrastantes de la cuenca de Cuitzeo, Michoacán. Se describen e ilustran en detalle todos los estadios de vida, incluyendo notas sobre su biología, fenología y plantas huéspedes.
The life cycle and temporal distribution of Stenomacra marginella (Herrich-Schaeffer) were studied. Adults and nymphs were collected as part of a project to study the Hemiptera diversity in contrasting environments in the Cuitzeo Basin, Michoacán. All life history stages are illustrated and described in detail, and notes on biology, phenology, and host plants are included.
El género Stenomacra Stål fue revisado por Brailovsky y Mayorga (1997). Se reconocen 8 especies, de las cuales S. marginella (Herrich-Schaeffer) es la única registrada de México. No se conoce nada acerca de los estados inmaduros de las especies del género y muy poco de los de la familia Largidae. Las ninfas de Largus californicus (Van Duzee) y L. rufipennis Laporte son aquellas que han sido estudiadas hasta el presente (Booth, 1990; Coscarón et al., 2004). Se sabe que algunas especies se alimentan de las semillas y savia de las plantas de diversas familias de dicotiledóneas (Ahmad y Schaefer, 1987). Para L. californicus se cita una gran variedad de plantas huéspedes (Booth, 1990) y para L. rufipennis,Coscarón y colaboradores (2004) reportan a Lantana entrerriensis Tronc. (Verbenaceae). Se han estudiado varios aspectos de la biología y comportamiento de S. marginella, como el tracto digestivo (Nava et al., 2007) y los mecanismos de cópula (Moreno y Cordero, 2008; Oliver y Cordero, 2009). A esta especie se le conoce como chinche roja o brujita, y dado que forman grandes agregaciones, se le ha considerado desagradable o plaga en ciertas épocas del año en algunas regiones de México.
En el presente trabajo se describen e ilustran en detalle todos los estadios de S. marginella, incluyendo notas acerca de su biología, plantas huéspedes y su distribución en México.
Materiales y métodosLos sitios de estudio están localizados dentro de la cuenca de Cuitzeo, municipio de Morelia, en la localidad cerro El Águila, ubicada en las coordenadas 19°38'43” N, 101°20'45” O, a 2 239m snm. La vegetación de la zona es principalmente de bosque tropical caducifolio, predominando especies arbóreas de Acacia, Bursera y Quercus.
Los muestreos se realizaron mensualmente entre abril del 2011 y marzo del 2012, estableciendo tanto muestreos diurnos como nocturnos, con diferentes métodos de recolecta, capturándose un total de 175 individuos: Recolecta directa en cuadrantes: se hizo un transecto de 100m de longitud, sobre el cual se ubicaron de forma aleatoria 2 cuadrantes de 400m2. Dentro de éstos, se recolectó buscando debajo de piedras, en cortezas de árboles muertos, en el suelo y en la vegetación. El esfuerzo de muestreo fue de 2 horas/hombre en cada cuadrante (143 individuos).
Recolecta con red de golpeo: dentro de los cuadrantes establecidos se obtuvieron muestras para cada estrato de la vegetación (árboles, arbustos y herbáceas), realizando 5 redazos (1 redazo= 5 golpes en la vegetación) con 4 repeticiones en cada estrato, esto dentro de los cuadrantes que ya habían sido establecidos (27 individuos).
Trampas de caída: En el transecto de 100m se colocaron 20 trampas de caída en forma de zigzag con 5m de separación entre cada una (4 individuos).
Reolecta nocturnas: se colocaron 2 pantallas con luz blanca fluorescente, estableciendo una distancia de 30m entre ellas; el horario de recolecta fue de las 21:00 a 23:00 (1 individuo).
Las plantas huéspedes registradas durante este estudio son Condalia velutina I. M. Johnst. (Rhamnaceae), que en el área se conoce comúnmente como acebuche, granjeno rojo, granjeno o pico de pájaro; es un arbusto de 2 a 7m de alto. Esta especie se encuentra principalmente en pastizales junto con elementos arbustivos de Acacia; también se le observa como componente del bosque de encinos, el bosque tropical caducifolio y el matorral xerófilo que se desarrolla sobre suelos derivados de roca ígnea, en altitudes entre 1 800-2 400m snm. El periodo de floración es muy amplio, de diciembre a septiembre (Fernández, 1996). En la zona de estudio se le observó fructificando durante abril. Otra planta húesped es Forestiera pubescens Nutt. (Oleaceae), conocida también en la zona con nombres comunes de C. velutina. Es un arbusto de hasta 6m de alto, facultativamente caducifolio, propio de matorrales xerófilos, pastizales, bosques tropicales caducifolios, así como de algunos encinares y pinares contiguos, a menudo en la vegetación secundaria, en altitudes de 1 700-2 400m snm; florece de octubre a marzo y se ha recolectado con fruto de marzo a octubre (Rzedowski y Rzedowski, 2004).
Los insectos estudiados fueron depositados en la Colección Entomológica de la Universidad Michoacana de San Nicolás de Hidalgo, Facultad de Biología, Laboratorio de Entomología “Biólogo Sócrates Cisneros Paz” (CEFBUM), Colección Entomológica del Instituto de Ecología, A. C. (IEXA) y en la Colección Nacional de Insectos del Instituto de Biología, UNAM (CNIN). Las ilustraciones se realizaron con ayuda de un microscopio Leica MZ8 adaptado a un tubo de dibujo. Las medidas se expresan en milímetros ± una desviación estándar. El número de individuos observados fue variado y se indica en cada etapa.
DescripciónStenomacra marginella (Herrich-Schaeffer, 1850) (Figs. 1–8)
. 1, vista dorsal del huevo; 2, vista lateral del huevo (la escala es la misma para ambas vistas); 3, primer estadio; 4, segundo estadio; 5, tercer estadio; 6, cuarto estadio; 7, quinto estadio; 8, adulto macho.")
Huevo (Figs. 1–2): Ovalado, bastante alargado, de color amarillento al ser depositado y tornándose rosado a rojo al madurar el embrión. Opérculo rodeado por alrededor de 11 procesos micropilares. En el campo se encontró una masa de 15 huevos dispuestos de manera irregular. De 1.21±0.8 de largo por 0.71±0.03 de ancho (n=10).
Primer estadio (Fig. 3): ligeramente piriforme, con anchura máxima en el segmento abdominal IV. Cabeza, antenas, rostro, tórax, patas y abdomen de color pardo oscuro, casi negro; uniones entre los segmentos antenales, coxas de todas las patas, cuarto basal de los fémures medios y posteriores, línea de dehiscencia del tórax, márgenes laterales del pro, meso, metanoto y del abdomen, amarillo pálido. Ojos prominentes y de color rojo oscuro. Vientre de la cabeza, la pro, meso y metapleura de color pardo oscuro, casi negro. Parte media ventral del tórax de color rojo anaranjado. Abdomen ventralmente con 3 bandas pardo oscuro a negro o gris; situadas una sobre cada margen lateral y una media, resto del abdomen anaranjado rojizo. Cuerpo cubierto dorsal y ventralmente por numerosas sedas finas blanquecinas; dorsalmente, además, con sedas gruesas negras, siendo más abundantes sobre los márgenes laterales de todo el cuerpo. Tylus más largo que los juga. Rostrum llegando hasta las metacoxas. Fémures anteriores con 3 espinas robustas subapicales sobre su margen dorsal. Orificios de las glándulas senescentes no aparentes. Medidas (n=1). Longitud total del cuerpo: 2.7. Cabeza. Longitud total: 0.5; anchura entre los ojos: 0.75; distancia interocular: 0.45. Longitud de los artejos antenales: I, 0.4; II, 0.35; III, 0.27; IV, 0.57. Artejos rostrales: I, 0.35; II, 0.32; III, 0.25; IV, 0.3. Pronoto. Longitud total: 0.35; anchura entre los ángulos frontales: 0.6; anchura entre los ángulos humerales: 0.87. Pata anterior: fémur: 0.5; tibia: 0.57; tarso I: 0.1; tarso II: 0.2.
Segundo estadio (Fig. 4): bastante piriforme, con anchura máxima en el segmento abdominal IV. Muy similar al primer estadio, sin embargo, aumenta en tamaño y el abdomen dorsalmente adquiere una coloración rojiza, la cual puede abarcar sólo la parte media del segmento abdominal II, o bien, puede llegar hasta el segmento IV. La región del cuello y de la línea de dehiscencia de la cabeza adquieren coloración blanca rojiza. El rostro alcanza sólo las mesocoxas. Orificios de glándulas senescentes aparecen como pequeños óvalos amarillos situados medianamente entre los segmentos III-IV, IV-V y V-VI. Espiráculos como puntos negros, situados ventralmente sobre el margen medio de cada segmento. Medidas (n=10). Longitud total del cuerpo: 3.16±0.12. Cabeza. Longitud total: 0.65±0.04; anchura entre los ojos: 0.78±0.03; distancia interocular: 0.47±0.1. Longitud de los artejos antenales: I, 0.5±0.09; II, 0.42±0.03; III, 0.32±0.03; IV, 0.55±0.07. Artejos rostrales: I, 0.36±0.05; II, 0.31±0.05; III, 0.29±0.04; IV, 0.28±0.02. Pronoto. Longitud total: 0.38±0.03; anchura entre los ángulos frontales: 0.6±0.04; anchura entre los ángulos humerales: 0.8±0.07. Pata anterior: fémur: 0.69±0.05; tibia: 0.64±0.03; tarso I: 0.15±0.02; tarso II: 0.26±0.02.
Tercer estadio (Fig. 5): alargado, con anchura máxima en el segmento abdominal III. Muy similar al segundo estadio, pero la coloración parda se hace todavía más oscura. Mancha rojiza del abdomen se hace más definida aunque varía en el área cubierta. Ventralmente, el abdomen se vuelve también más rojizo, ya que la banda parda media desaparece en su parte anterior, cubriendo sólo la mitad posterior del esternito VI a la parte terminal del abdomen. Pubescencia negra gruesa del cuerpo desaparece casi completamente. Medidas (n=10). Longitud total del cuerpo: 4.42±0.26. Cabeza. Longitud total: 0.81±0.05; anchura entre los ojos: 1.01±0.05; distancia interocular: 0.67±0.07. Longitud de los artejos antenales: I, 0.8±0.05; II, 0.63±0.04; III, 0.45±0.07; IV, 0.76±0.05. Artejos rostrales: I, 0.46±0.03; II, 0.37±0.07; III, 0.36±0.03; IV, 0.36±0.03. Pronoto. Longitud total: 0.58±0.02; anchura entre los ángulos frontales: 0.77±0.08; anchura entre los ángulos humerales: 1.06±0.08. Pata anterior: fémur: 0.96±0.03; tibia: 0.9±0.06; tarso I: 0.25±0.02; tarso II: 0.35±0.02.
Cuarto estadio (Fig. 6): muy alargado, siendo todavía ligeramente más ancho en el segmento abdominal III. Muy similar al estadio III. Cabeza y tórax adquieren coloración más oscura, los márgenes laterales del pro, meso y metanoto son oscuros. Abdomen también adquiere coloración más oscura, sin embargo, el margen lateral sigue siendo amarillo pálido. Mancha roja del abdomen sigue siendo de tamaño variado, desde casi circular hasta romboidal. Almohadillas alares mesotoráxicas cubren casi toda la parte lateral del metanoto. Medidas (n=9). Longitud total del cuerpo: 6.24±0.42. Cabeza. Longitud total: 0.68±0.17; anchura entre los ojos: 1.23±0.07; distancia interocular: 0.8±0.03. Longitud de los artejos antenales: I, 1.22±0.19; II, 0.62±0.11; III, 0.56±0.07; IV, 0.87±0.14. Artejos rostrales: I, 0.59±0.1; II, 0.55±0.04; III, 0.48±0.04; IV, 0.38±0.05. Pronoto. Longitud total: 0.76±0.11; anchura entre los ángulos frontales: 0.76±0.11; anchura entre los ángulos humerales: 1.52±0.14. Pata anterior: fémur: 1.13±0.11; tibia: 1.27±0.2; tarso I: 0.37±0.08; tarso II: 0.32±0.07.
Quinto estadio (Fig. 7): Cuerpo muy alargado, sin que exista diferencia en la anchura del mesonoto, metanoto y abdomen. Coloración igual que en el cuarto estadio; sin embargo, la mancha roja del abdomen es menos variada y se sitúa casi exclusivamente sobre la parte media visible del segmento abdominal II y en la parte media anterior del segmento III, en un área bien definida por las almohadillas alares que alcanzan poco más de la parte media del segmento abdominal III. Parte media ventral de casi todo el cuerpo amarillenta, con excepción de los últimos esternitos abdominales que conservan la banda oscura. Medidas (n=10). Longitud total del cuerpo: 9.03±0.6. Cabeza. Longitud total: 0.97±0.13; anchura entre los ojos: 1.55±0.04; distancia interocular: 0.98±0.06. Longitud de los artejos antenales: I, 1.85±0.15; II, 1.27±0.11; III, 0.8±0.06; IV, 1.36±0.09. Artejos rostrales: I, 0.88±0.09; II, 0.73±0.05; III, 0.73±0.04; IV, 0.53±0.04. Pronoto. Longitud total: 1.22±0.09; anchura entre los ángulos frontales: 1.22±0.09; anchura entre los ángulos humerales: 2.21±0.16. Pata anterior: fémur: 2.06±0.26; tibia: 2.02±0.12; tarso I: 0.54±0.06; tarso II: 0.65±0.03.
Adulto (Fig. 8): esta especie se distingue fácilmente de las otras especies del género por tener los segmentos antenales I a IV, los segmentos rostrales I a IV, las tibias y tarsos completamente negros a pardo rojizos y por tener las siguientes áreas siempre anaranjadas a amarillo anaranjadas: tubérculos anteníferos, búcula, márgenes del pronoto, incluyendo el margen costal del corium, sutura anal y comisura claval, ápice del escutelo, margen superior de los segmentos conexivales, acetábulo, coxas, y trocánteres. Esta especie es extremadamente variada en coloración, y varias formas pueden ser encontradas dentro de una población: 1), cabeza dorsalmente negra con tonos anaranjados o sólo con la región entre los ojos negra. 2), lóbulo anterior del pronoto anaranjado o amarillo y lóbulo posterior negro, con o sin una línea media longitudinal, o lóbulos anterior y posterior negros con una línea media longitudinal amarilla, o ambos lóbulos con una difusa línea amarilla sobre un fondo pardo. 3), escutelo negro con el ápice anaranjado a amarillo, o escutelo anaranjado a amarillo con el tercio basal negro. 4), fémures completamente negros o pardo rojizos, o completamente anaranjados, o bien, con el tercio basal anaranjado y el resto negro, o anaranjado con sólo la cara dorsal del tercio apical negro (Brailovsky y Mayorga, 1997). Medidas macho (n=10). Longitud total del cuerpo: 11.15±0.27. Cabeza. Longitud total: 1.17±0.05; anchura entre los ojos: 1.77±0.09; distancia interocular: 0.97±0.07. Longitud de los artejos antenales: I, 2.73±0.45; II, 1.85±0.17; III, 1.11±0.07; IV, 1.67±0.1. Artejos rostrales: I, 1.02±0.07; II, 0.95±0.06; III, 0.87±0.09; IV, 0.6±0.05. Pronoto. Longitud total: 1.99±0.13; anchura entre los ángulos frontales: 1.5±0.15; anchura entre los ángulos humerales: 2.68±0.15. Escutelo. Longitud total 1.26±0.09; anchura máxima 1.42±0.15. Pata anterior: fémur: 2.93±0.14; tibia: 2.85±0.16; tarso I: 0.8±0.05; tarso II: 0.31±0.04; tarso III: 0.46±0.06. Medidas hembra (n=10). Longitud total del cuerpo: 12.81±0.79. Cabeza. Longitud total: 1.21±0.02; anchura entre los ojos: 1.83±0.08; distancia interocular: 1.14±0.05. Longitud de los artejos antenales: I, 2.89±0.31; II, 1.74±0.23; III, 1.17±0.15; IV, 1.52±0.16. Artejos rostrales: I, 1.02±0.07; II, 1.07±0.1; III, 1±0.09; IV, 0.55±0.11. Pronoto. Longitud total: 2.25±0.22; anchura entre los ángulos frontales: 1.58±0.11; anchura entre los ángulos humerales: 3.11±0.13. Escutelo. Longitud total 1.43±0.01; anchura máxima 1.71±0.06. Pata anterior: fémur: 3.08±0.18; tibia: 2.82±0.16; tarso I: 0.87±0.1; tarso II: 0.27±0.04; tarso III: 0.44±0.05.
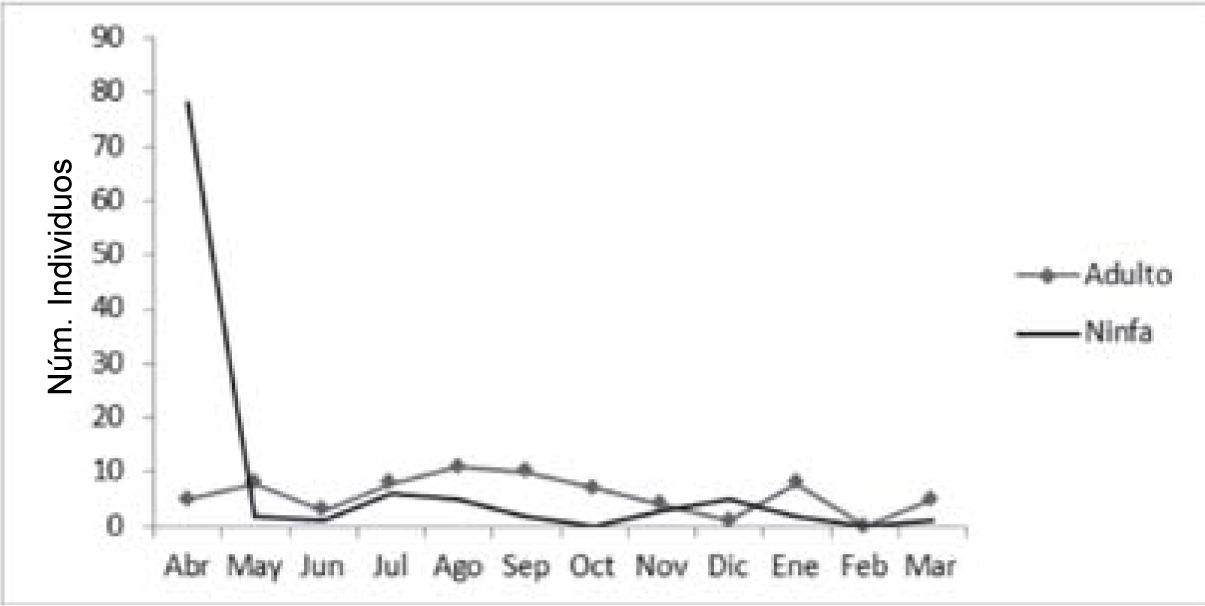
Resumen taxonómicoBiología. Se observaron agrupaciones de esta chinche sobre C. velutina (Rhamnaceae), encontrándose tanto ninfas como adultos. Estos últimos llevaban a cabo la cópula entre abril y mayo, refugiándose entre las espinas y follaje de la planta. En F. pubescens (Oleaceae) también se observó a S. marginella, sin embargo, sobre esta planta húesped no se encontraron de forma agregada; se halló una masa de huevos en el envés de las hojas, formando una masa, sin arreglo simétrico. Tanto ninfas como adultos se alimentan principalmente de la savia de los frutos, pero también del follaje. La mayor abundancia de individuos, principalmente ninfas de segundo a cuarto estadio, se registró en abril, coincidiendo con la época de fructificación de C. velutina y F. pubescens, temporada que también coincide con un abundante follaje, favoreciendo así la disponibilidad de alimento y refugio. Durante los siguientes meses del año se registraron muy pocas ninfas y casi exclusivamente de los últimos estadios. La abundancia de adultos a lo largo del año se mantuvo más o menos constante, en el mes de febrero no se registró ningún individuo (Fig. 9). En la ciudad de México, se le ha encontrado formando grandes agregaciones sobre eucaliptos (Eucalyptus spp.) y tepozán (Buddleja cordata Kunth), en el área del Pedregal de San Ángel.

Distribución. Estados Unidos de América, México, Guatemala, Nicaragua, Costa Rica, Panamá, Colombia, Venezuela y Brasil. México: Coahuila, Colima, Michoacán, Guanajuato, Jalisco, Querétaro, Morelos, Guerrero, Hidalgo, Estado de México, Puebla y Distrito Federal, Oaxaca, Veracruz y Chiapas (Brailovsky y Mayorga, 1997).
Comentarios taxonómicos. Aunque esta especie forma grandes agregaciones, y se le ha considerado desagradable o plaga en ciertas épocas del año y en algunas regiones de México, es importante resaltar que no causa ningún daño directo o indirecto al hombre. En ciertas ocasiones y debido al gran número de individuos que pueden existir en una sola área, S. marginella puede ser molesta.
Se documenta por primera vez el ciclo biológico de esta especie de amplia distribución en México. En comparación con los ciclos de vida conocidos de otros lárgidos, los estadios de S. marginella son muy similares a los de las especies de Largus conocidas. Casi todos sus estadios son bastante más alargados que los de las especies de Largus. La coloración en general es menos brillante en S. marginella; además, los fémures anteriores de todos los estadios presentan 3 espinas subdistales características del género.
Es probable que esta especie en otras regiones del país tenga plantas huéspedes que estén relacionadas con los géneros Condalia y Forestiera o al menos con miembros de las Rhamnaceae y Oleaceae. Estas familias son diferentes de las registradas para otras especies de Largidae (Ahmad y Schaefer, 1987; Booth, 1990; Coscarón et al., 2004), o al menos para miembros de las familias mencionadas. Cuando esta especie forma grandes agregaciones suele encontrarse sobre otras plantas o sobre el suelo, sin embargo, no se consideran plantas huéspedes, ya que no se alimentan y sólo se encuentran reposando sobre ellas.
Esta especie parece tener una sola generación al año en esta zona y su ciclo de vida está muy relacionado con la floración y fructificación de sus plantas huéspedes, que comienzan en marzo, aunque es necesario realizar estudios durante periodos más largos. El saber que tiene una sola generación al año, el conocer el ciclo de vida, el que su etapa ninfal es relativamente corta y que su ciclo de vida está sincronizado con la fenología de sus plantas huéspedes, nos ayudan a considerar medidas de control que en algún momento podrían aplicarse cuando las agregaciones de esta especie resulten molestas. Quizá resulte más fácil el control durante el periodo de oviposición y sólo enfocado sobre las plantas huéspedes.
A Cristina Mayorga, del Instituto de Biología (UNAM), por las facilidades otorgadas para revisar el material depositado en la Colección Entomológica. Al Conacyt por la beca otorgada a la primera autora para la realización de estudios de Maestría, así como a la Coordinación de la Investigación Científica de la UMSNH por el financiamiento parcial para la realización de este estudio.