


La medida de biodiversidad más frecuentemente utilizada es la riqueza de especies y resulta especialmente importante detectar cuántas veces es mayor la riqueza de una comunidad con respecto a otra, es decir, la proporción de riqueza o diferencia relativa en el número de especies entre comunidades. Para calcular esta proporción de riqueza es necesario estandarizar las muestras. En este trabajo se utilizan 2 métodos de estandarización para calcular la proporción de riqueza: la rarefacción tradicional basada en el número de individuos y la rarefacción basada en el grado de completitud del muestreo (cobertura de la muestra). Con ambos métodos se calculó la proporción de riqueza de murciélagos entre selvas y hábitats modificados del neotrópico. La información se obtuvo de 53 estudios de caso reportados en 34 publicaciones. Con el método de rarefacción tradicional, 35 casos presentaron mayor riqueza en la selva, mientras que con la rarefacción basada en la cobertura de la muestra, 38 casos resultaron con mayor riqueza en la selva que en el hábitat modificado. La proporción de riqueza calculada con el nuevo método de rarefacción resultó significativamente mayor que la calculada con la rarefacción tradicional. Esto significa que con la rarefacción tradicional la diferencia en la riqueza entre selvas y hábitats modificados es menor que la diferencia real. Para evitar esta reducción en la proporción de riqueza y sus posibles consecuencias en biología de la conservación, se recomienda el uso de la rarefacción por cobertura de la muestra. Estos resultados pueden ser aplicables ampliamente, más allá del grupo biológico y de la región.
The most frequently used measure of biodiversity is species richness, and it is especially important to detect how many times is richer one community regarding another one, i.e., the species richness ratio or relative difference in the number of species between communities. To assess this richness ratio, it is necessary to standardize samples. In this work we use 2 standardizing methods to assess the richness ratio: traditional rarefaction based on the number of individuals, and rarefaction based on the degree of sample completeness (sample coverage). We used both methods to analyze bat richness ratio between tropical forests and modified habitats in the Neotropics. We used information from 53 study cases reported in 34 published papers. With traditional rarefaction, 35 studies showed higher species richness in the forest, while rarefaction based on sample coverage led to 38 studies with higher richness in the forest than in the modified habitat. Richness ratios were higher with the new rarefaction method than with the traditional rarefaction. This means that traditional rarefaction gives a richness difference between forest and modified habitats lower than the real difference. To avoid this reduction in richness ratios, and its possible consequences in conservation biology, it would be better to use sample coverage based rarefaction. These results can be widely applicable, beyond the study group and the region.
El número de especies (riqueza específica) es el atributo más frecuentemente utilizado para describir una comunidad, porque es una medida rápida y sencilla de la diversidad (Gaston, 1996; Gotelli y Colwell, 2011). Sin embargo, a pesar de su sencillez, lograr una evaluación adecuada de la riqueza tiene varios retos que requieren superarse.
El primer reto se origina por el hecho de que los muestreos rara vez son suficientes para registrar el número total de especies de una comunidad. El reto consiste en predecir ese número total, para lo cual se utilizan múltiples métodos que modelan las curvas de acumulación de especies conforme se incrementa el esfuerzo de muestreo. Por ejemplo, Thompson, Withers, Pianka y Thompson (2003) describen 12 modelos paramétricos de regresión no lineal, mientras que el programa EstimateS (Colwell, 2013) permite calcular por lo menos 10 métodos no paramétricos. Estos métodos estiman valores máximos de riqueza y se suelen utilizar para evaluar la completitud de los inventarios.
Un segundo reto se origina por el hecho de que la riqueza de especies depende del tamaño de la muestra que se tome de la comunidad objetivo, por lo que, para poder hacer comparaciones, las muestras deben tener igual número de individuos (Colwell, Mao y Chang, 2005). Si esto se ignora, las comparaciones de la riqueza de especies entre comunidades estarán sesgadas (Gotelli y Colwell, 2011). El reto consiste en resolver la dependencia del tamaño de muestra y comparar la riqueza cuando el número de individuos no es igual, y para lograr esto existen 2 soluciones: interpolar o extrapolar. La interpolación de la riqueza de especies se realiza mediante la técnica de rarefacción, basada en la forma de la curva de especies-abundancia (Sanders, 1968). Esta rarefacción tradicional calcula el número esperado de especies reduciendo las muestras a un tamaño estándar, es decir, interpolando a un mismo número de individuos: el de la comunidad que presente la menor abundancia (Jiménez-Valverde y Hortal, 2003; Moreno, 2001). También es posible realizar una rarefacción interpolando el número de muestras en lugar del número de individuos (Gotelli y Colwell, 2011). No obstante, en ambos casos, la interpolación desaprovecha mucha información, dejando a un lado los datos adicionales obtenidos en las comunidades con mayor esfuerzo de muestreo (Ludwig y Reynolds, 1988; Moreno, 2001).
Para evitar esta pérdida de información, la segunda solución que controla la dependencia del tamaño de muestra consiste en extrapolar la riqueza de especies. La importancia de la extrapolación radica en la necesidad de tener muestras más grandes en el sentido estadístico, incrementando el tamaño de los conjuntos de datos pequeños para su comparación con conjuntos más grandes, conservando la información completa de los inventarios. Este procedimiento se vuelve menos confiable conforme se incrementa el tamaño de la muestra (Colwell et al., 2005).
Aun cuando la riqueza de especies haya sido estandarizada a un mismo tamaño de muestra, sea por interpolación o por extrapolación, surge un tercer reto: cómo ordenar 2 o más comunidades de acuerdo con las diferencias estadísticas en el número de especies entre comunidades. Dichas diferencias estadísticas se evalúan mediante intervalos de confianza calculados alrededor de las curvas de acumulación (Pineda y Moreno, 2015). Para ello, la opción más común es evaluar la sobreposición en los valores de los intervalos de confianza al 95%, sin embargo, se pueden utilizar como alternativa intervalos de confianza al 84%, para mimetizar pruebas estadi¿sticas con α=0.05 (MacGregor-Fors y Payton, 2013).
Ya sea que hayamos detectado o no las diferencias estadísticas en la riqueza, queda aún pendiente un cuarto reto: detectar cuántas veces es mayor la riqueza de una comunidad con respecto a otra. Dos comunidades, por ejemplo con 5 y 7 especies, podrían tener una diferencia estadísticamente significativa en su riqueza (por la separación en los intervalos de confianza alrededor de sus curvas de acumulación de especies), mientras que 2 comunidades con 5 y 10 especies podrían no diferir estadísticamente en su riqueza (por sobrelapamiento en los intervalos de confianza). En el primer caso, la riqueza de una comunidad es 1.4 veces mayor que la otra, mientras que en el segundo caso una comunidad es 2 veces más rica que la otra. Esta diferencia relativa en el número de especies entre comunidades se conoce como proporción de riqueza.
Para el cuarto reto de evaluar la proporción de riqueza, sin la pérdida de información inherente de la rarefacción tradicional basada en el número de individuos (Sanders, 1968), recientemente se propuso un método de rarefacción basado en el nivel de completitud del inventario (Chao y Jost, 2012). La completitud del inventario se evalúa mediante la «cobertura de la muestra», que mide la proporción que representan los individuos de cada especie en la muestra, con respecto al número total de individuos. Este método analítico estandariza comunidades igualmente muestreadas, de manera que aprovecha la mayor parte de la información obtenida en campo, y se propone como un método más adecuado que la rarefacción tradicional para evaluar la proporción de riqueza (Chao y Jost, 2012). La proporción de riqueza permite responder preguntas de gran importancia en el ámbito de la conservación, por ejemplo, ¿Qué tanto más rica en especies es una comunidad con respecto a la otra? (Jost, 2006), o bien, ¿Qué proporción de la riqueza cambia entre 2 comunidades? Como podría ser el caso entre áreas con diferente grado de alteración antropogénica.
En este trabajo se abordan los métodos disponibles únicamente para el cuarto reto. El objetivo es poner a prueba con datos empíricos los 2 métodos de rarefacción (basada en número de individuos y basada en la completitud del inventario) para medir la proporción de riqueza entre comunidades. Para ello, se evalúa la proporción de riqueza de murciélagos del neotrópico, comparando entre zonas con vegetación nativa (selva tropical) y hábitats modificados por el hombre (monocultivos, pastizales, sistemas agroforestales y vegetación secundaria). Los murciélagos neotropicales han sido un grupo ampliamente estudiado, principalmente por su facilidad de muestreo y taxonomía estable, y constituyen un grupo focal para evaluar cómo cambia la estructura de una comunidad en cuanto a su riqueza y composición de especies al transformarse los ambientes prístinos en ambientes modificados (García-Morales, Badano y Moreno, 2013; Meyer, 2014; Meyer et al., 2015; Montaño-Centellas et al., 2015). Considerando que existen numerosos trabajos sobre las comunidades de murciélagos neotropicales en los cuales se reporta el esfuerzo de muestreo y la abundancia de las especies, resulta un grupo ideal para analizar el desempeño de estos 2 métodos de rarefacción. Debido a que la comparación de la riqueza de especies entre las comunidades es uno de los objetivos más comunes en ecología, biogeografía y biología de la conservación, los resultados de esta comparación de métodos podrían ser de amplio interés, más allá del grupo biológico y de la región.
Materiales y métodosLos datos se tomaron de una selección de publicaciones de estudios realizados en el neotrópico recopiladas por García-Morales et al. (2013). Estos trabajos comparan las comunidades de murciélagos en sitios de selva y con hábitats modificado por las actividades humanas. En todos los trabajos el muestreo se realizó únicamente con redes de niebla, por lo que los datos no corresponden a toda la comunidad, sino a los murciélagos que vuelan cerca del suelo. Se realizó una depuración de las publicaciones, eliminando aquellas que no describieran con claridad el tipo de vegetación del área de estudio, así como la ubicación y los métodos de muestreo. Solo se consideraron las publicaciones en las que se contrastara la selva tropical, contra alguno de 5 tipos de hábitats modificados por el hombre: monocultivos, pastizales, sistemas agroforestales, vegetación secundaria joven y vegetación secundaria madura. De cada publicación se registró el número de individuos capturados por especie en cada hábitat y se evaluó la completitud de los muestreos mediante la cobertura de la muestra (Chao y Jost, 2012). En el análisis se consideraron únicamente las comunidades que estuvieran bien muestreadas (mínimo un 90% de completitud).
El análisis de los datos se realizó entre pares de hábitats: la vegetación nativa (selva tropical) y alguno de los diferentes tipos de vegetación de cada artículo. De esta manera, cada par es un caso de estudio que incluye una comunidad de murciélagos de la selva tropical y una comunidad de murciélagos en alguno de los hábitats modificados.
Para la rarefacción tradicional basada en el número de individuos, se utilizó el programa PAST 3.0 (Hammer, Harper y Ryan, 2013) disponible en la página http://folk.uio.no/ohammer/past/. Con este método se obtuvo la riqueza de especies esperada para cada caso, estandarizando el tamaño de muestra al número de individuos de la comunidad con menor abundancia.
Para el análisis de la rarefacción por cobertura de la muestra se utilizó el procedimiento de Chao y Jost (2012) en el programa iNEXT (Hsieh, Ma y Chao, 2013; disponible en: http://chao.stat.nthu.edu.tw/wordpress/software_download/inext-online/). Los análisis se realizaron con 100 aleatorizaciones y extrapolando al doble del número de individuos de la comunidad con la menor cobertura de muestra (Chao y Jost, 2012).
Con el valor esperado de riqueza obtenido con cada método de rarefacción se calculó la proporción de la riqueza entre la selva y el hábitat modificado. La proporción de riqueza permite comparar de forma relativa la magnitud de las diferencias en la riqueza de especies entre 2 comunidades, en función de la comunidad más rica, de la siguiente manera: SA/SB, si una comunidad A tiene una riqueza SA mayor que la riqueza SB de la comunidad B, entonces la comunidad A será SA/SB veces más rica en especies que la comunidad B.
Se realizó una correlación entre las proporciones de riqueza obtenidas mediante los 2 tipos de rarefacción para evaluar si los resultados obtenidos con ambos métodos siguen la misma tendencia, y se realizó una prueba de signos de Wilcoxon (t pareada para datos no paramétricos) para probar si las diferencias observadas en la magnitud estimada entre los 2 métodos eran estadísticamente significativas.
Mediante un análisis de varianza se comprobó si existían diferencias significativas en la proporción de riqueza encontrada entre los 5 tipos de hábitats modificados. Para la rarefacción por tamaño de muestra utilizamos un Anova de una vía (datos con distribución normal) y para la rarefacción por cobertura de muestra usamos una prueba de Kruskal-Wallis (datos sin distribución normal).
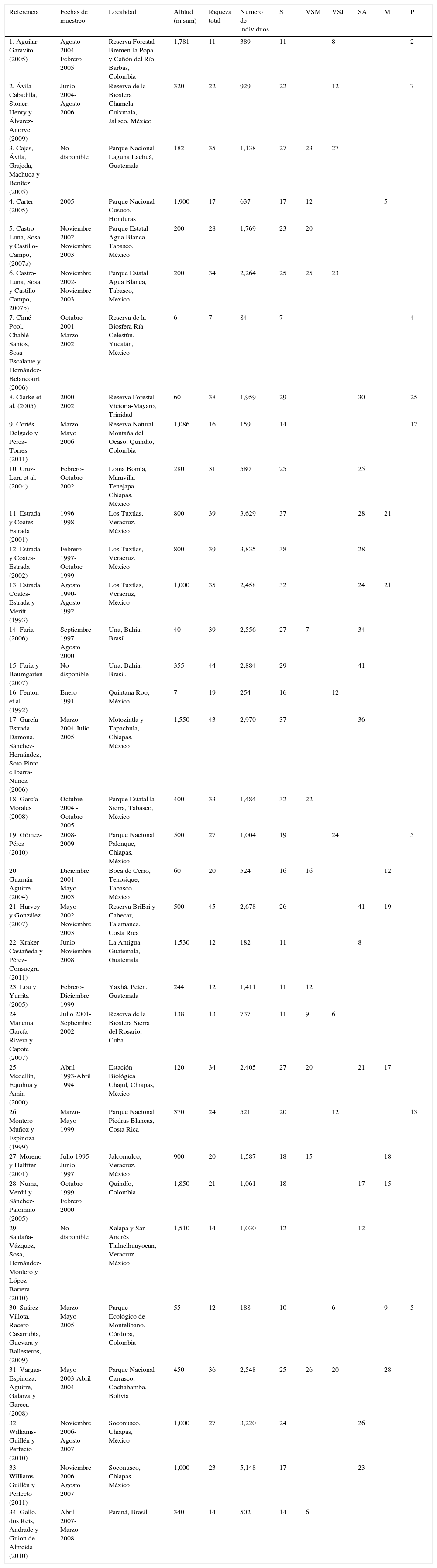
ResultadosSe encontró que 34 publicaciones cumplían los requisitos para ser incorporadas a los análisis (tabla 1). Los trabajos fueron realizados en 10 países, desde México hasta Brasil. Para las comparaciones de riqueza de especies, se trabajó con 53 casos. La completitud del inventario en la mayoría de los casos estuvo por arriba del 90%, y en algunos alcanzó el 100% (tabla 2).
Lista de trabajos seleccionados para la realización del presente estudio. En todos ellos se muestrearon con redes de niebla las comunidades de murciélagos en sitios de selva y por lo menos en un sitio con hábitat modificado por las actividades humanas en el neotrópico. Se muestra la riqueza de especies en cada tipo de vegetación.
| Referencia | Fechas de muestreo | Localidad | Altitud (m snm) | Riqueza total | Número de individuos | S | VSM | VSJ | SA | M | P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Aguilar-Garavito (2005) | Agosto 2004-Febrero 2005 | Reserva Forestal Bremen-la Popa y Cañón del Río Barbas, Colombia | 1,781 | 11 | 389 | 11 | 8 | 2 | |||
| 2. Ávila-Cabadilla, Stoner, Henry y Álvarez-Añorve (2009) | Junio 2004-Agosto 2006 | Reserva de la Biosfera Chamela-Cuixmala, Jalisco, México | 320 | 22 | 929 | 22 | 12 | 7 | |||
| 3. Cajas, Ávila, Grajeda, Machuca y Benítez (2005) | No disponible | Parque Nacional Laguna Lachuá, Guatemala | 182 | 35 | 1,138 | 27 | 23 | 27 | |||
| 4. Carter (2005) | 2005 | Parque Nacional Cusuco, Honduras | 1,900 | 17 | 637 | 17 | 12 | 5 | |||
| 5. Castro-Luna, Sosa y Castillo-Campo, (2007a) | Noviembre 2002-Noviembre 2003 | Parque Estatal Agua Blanca, Tabasco, México | 200 | 28 | 1,769 | 23 | 20 | ||||
| 6. Castro-Luna, Sosa y Castillo-Campo, 2007b) | Noviembre 2002-Noviembre 2003 | Parque Estatal Agua Blanca, Tabasco, México | 200 | 34 | 2,264 | 25 | 25 | 23 | |||
| 7. Cimé-Pool, Chablé-Santos, Sosa-Escalante y Hernández-Betancourt (2006) | Octubre 2001-Marzo 2002 | Reserva de la Biosfera Ría Celestún, Yucatán, México | 6 | 7 | 84 | 7 | 4 | ||||
| 8. Clarke et al. (2005) | 2000-2002 | Reserva Forestal Victoria-Mayaro, Trinidad | 60 | 38 | 1,959 | 29 | 30 | 25 | |||
| 9. Cortés-Delgado y Pérez-Torres (2011) | Marzo-Mayo 2006 | Reserva Natural Montaña del Ocaso, Quindío, Colombia | 1,086 | 16 | 159 | 14 | 12 | ||||
| 10. Cruz-Lara et al. (2004) | Febrero-Octubre 2002 | Loma Bonita, Maravilla Tenejapa, Chiapas, México | 280 | 31 | 580 | 25 | 25 | ||||
| 11. Estrada y Coates-Estrada (2001) | 1996-1998 | Los Tuxtlas, Veracruz, México | 800 | 39 | 3,629 | 37 | 28 | 21 | |||
| 12. Estrada y Coates-Estrada (2002) | Febrero 1997-Octubre 1999 | Los Tuxtlas, Veracruz, México | 800 | 39 | 3,835 | 38 | 28 | ||||
| 13. Estrada, Coates-Estrada y Meritt (1993) | Agosto 1990-Agosto 1992 | Los Tuxtlas, Veracruz, México | 1,000 | 35 | 2,458 | 32 | 24 | 21 | |||
| 14. Faria (2006) | Septiembre 1997-Agosto 2000 | Una, Bahia, Brasil | 40 | 39 | 2,556 | 27 | 7 | 34 | |||
| 15. Faria y Baumgarten (2007) | No disponible | Una, Bahia, Brasil. | 355 | 44 | 2,884 | 29 | 41 | ||||
| 16. Fenton et al. (1992) | Enero 1991 | Quintana Roo, México | 7 | 19 | 254 | 16 | 12 | ||||
| 17. García-Estrada, Damona, Sánchez-Hernández, Soto-Pinto e Ibarra-Núñez (2006) | Marzo 2004-Julio 2005 | Motozintla y Tapachula, Chiapas, México | 1,550 | 43 | 2,970 | 37 | 36 | ||||
| 18. García-Morales (2008) | Octubre 2004 -Octubre 2005 | Parque Estatal la Sierra, Tabasco, México | 400 | 33 | 1,484 | 32 | 22 | ||||
| 19. Gómez-Pérez (2010) | 2008-2009 | Parque Nacional Palenque, Chiapas, México | 500 | 27 | 1,004 | 19 | 24 | 5 | |||
| 20. Guzmán-Aguirre (2004) | Diciembre 2001-Mayo 2003 | Boca de Cerro, Tenosique, Tabasco, México | 60 | 20 | 524 | 16 | 16 | 12 | |||
| 21. Harvey y González (2007) | Mayo 2002-Noviembre 2003 | Reserva BriBri y Cabecar, Talamanca, Costa Rica | 500 | 45 | 2,678 | 26 | 41 | 19 | |||
| 22. Kraker-Castañeda y Pérez-Consuegra (2011) | Junio-Noviembre 2008 | La Antigua Guatemala, Guatemala | 1,530 | 12 | 182 | 11 | 8 | ||||
| 23. Lou y Yurrita (2005) | Febrero-Diciembre 1999 | Yaxhá, Petén, Guatemala | 244 | 12 | 1,411 | 11 | 12 | ||||
| 24. Mancina, García-Rivera y Capote (2007) | Julio 2001-Septiembre 2002 | Reserva de la Biosfera Sierra del Rosario, Cuba | 138 | 13 | 737 | 11 | 9 | 6 | |||
| 25. Medellín, Equihua y Amin (2000) | Abril 1993-Abril 1994 | Estación Biológica Chajul, Chiapas, México | 120 | 34 | 2,405 | 27 | 20 | 21 | 17 | ||
| 26. Montero-Muñoz y Espinoza (1999) | Marzo-Mayo 1999 | Parque Nacional Piedras Blancas, Costa Rica | 370 | 24 | 521 | 20 | 12 | 13 | |||
| 27. Moreno y Halffter (2001) | Julio 1995-Junio 1997 | Jalcomulco, Veracruz, México | 900 | 20 | 1,587 | 18 | 15 | 18 | |||
| 28. Numa, Verdú y Sánchez-Palomino (2005) | Octubre 1999-Febrero 2000 | Quindío, Colombia | 1,850 | 21 | 1,061 | 18 | 17 | 15 | |||
| 29. Saldaña-Vázquez, Sosa, Hernández-Montero y López-Barrera (2010) | No disponible | Xalapa y San Andrés Tlalnelhuayocan, Veracruz, México | 1,510 | 14 | 1,030 | 12 | 12 | ||||
| 30. Suárez-Villota, Racero-Casarrubia, Guevara y Ballesteros, (2009) | Marzo-Mayo 2005 | Parque Ecológico de Montelíbano, Córdoba, Colombia | 55 | 12 | 188 | 10 | 6 | 9 | 5 | ||
| 31. Vargas-Espinoza, Aguirre, Galarza y Gareca (2008) | Mayo 2003-Abril 2004 | Parque Nacional Carrasco, Cochabamba, Bolivia | 450 | 36 | 2,548 | 25 | 26 | 20 | 28 | ||
| 32. Williams-Guillén y Perfecto (2010) | Noviembre 2006-Agosto 2007 | Soconusco, Chiapas, México | 1,000 | 27 | 3,220 | 24 | 26 | ||||
| 33. Williams-Guillén y Perfecto (2011) | Noviembre 2006-Agosto 2007 | Soconusco, Chiapas, México | 1,000 | 23 | 5,148 | 17 | 23 | ||||
| 34. Gallo, dos Reis, Andrade y Guion de Almeida (2010) | Abril 2007-Marzo 2008 | Paraná, Brasil | 340 | 14 | 502 | 14 | 6 |
M: monocultivo; P: pastizal; S: selva; SA: sistema agroforestal; VSJ: vegetación secundaria joven; VSM: vegetación secundaria madura.
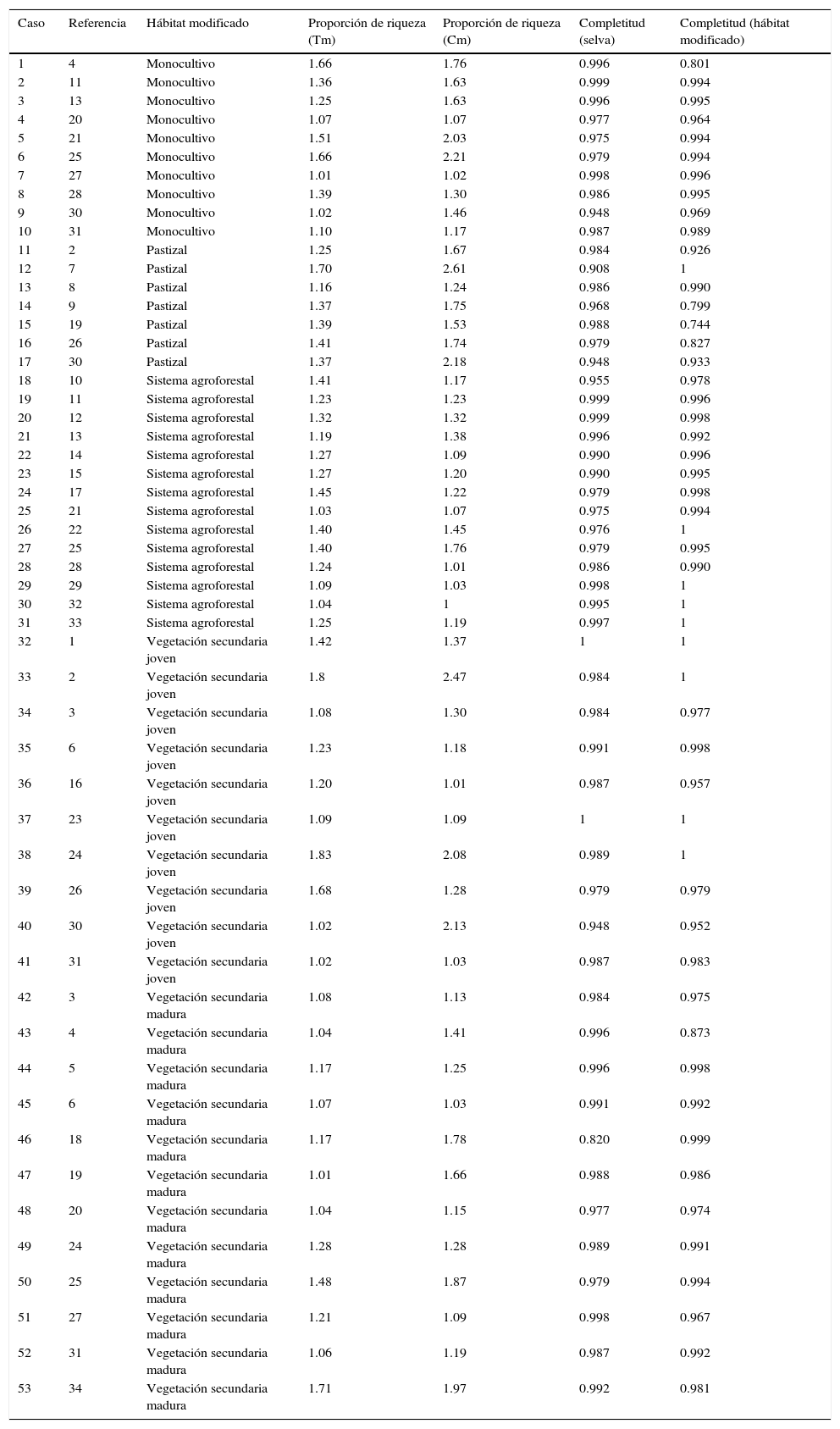
Casos de estudio utilizados para comparar 2 métodos de rarefacción: por tamaño de muestra (Tm) y por cobertura de muestra (Cm). La referencia bibliográfica de donde provienen los datos se indica en la tabla 1. Se indica el tipo de ambiente modificado con el que se comparó a la selva, la proporción de la riqueza calculada con ambos tipos de rarefacción, así como la completitud del inventario para cada tipo de vegetación (proporción de cobertura de muestra).
| Caso | Referencia | Hábitat modificado | Proporción de riqueza (Tm) | Proporción de riqueza (Cm) | Completitud (selva) | Completitud (hábitat modificado) |
|---|---|---|---|---|---|---|
| 1 | 4 | Monocultivo | 1.66 | 1.76 | 0.996 | 0.801 |
| 2 | 11 | Monocultivo | 1.36 | 1.63 | 0.999 | 0.994 |
| 3 | 13 | Monocultivo | 1.25 | 1.63 | 0.996 | 0.995 |
| 4 | 20 | Monocultivo | 1.07 | 1.07 | 0.977 | 0.964 |
| 5 | 21 | Monocultivo | 1.51 | 2.03 | 0.975 | 0.994 |
| 6 | 25 | Monocultivo | 1.66 | 2.21 | 0.979 | 0.994 |
| 7 | 27 | Monocultivo | 1.01 | 1.02 | 0.998 | 0.996 |
| 8 | 28 | Monocultivo | 1.39 | 1.30 | 0.986 | 0.995 |
| 9 | 30 | Monocultivo | 1.02 | 1.46 | 0.948 | 0.969 |
| 10 | 31 | Monocultivo | 1.10 | 1.17 | 0.987 | 0.989 |
| 11 | 2 | Pastizal | 1.25 | 1.67 | 0.984 | 0.926 |
| 12 | 7 | Pastizal | 1.70 | 2.61 | 0.908 | 1 |
| 13 | 8 | Pastizal | 1.16 | 1.24 | 0.986 | 0.990 |
| 14 | 9 | Pastizal | 1.37 | 1.75 | 0.968 | 0.799 |
| 15 | 19 | Pastizal | 1.39 | 1.53 | 0.988 | 0.744 |
| 16 | 26 | Pastizal | 1.41 | 1.74 | 0.979 | 0.827 |
| 17 | 30 | Pastizal | 1.37 | 2.18 | 0.948 | 0.933 |
| 18 | 10 | Sistema agroforestal | 1.41 | 1.17 | 0.955 | 0.978 |
| 19 | 11 | Sistema agroforestal | 1.23 | 1.23 | 0.999 | 0.996 |
| 20 | 12 | Sistema agroforestal | 1.32 | 1.32 | 0.999 | 0.998 |
| 21 | 13 | Sistema agroforestal | 1.19 | 1.38 | 0.996 | 0.992 |
| 22 | 14 | Sistema agroforestal | 1.27 | 1.09 | 0.990 | 0.996 |
| 23 | 15 | Sistema agroforestal | 1.27 | 1.20 | 0.990 | 0.995 |
| 24 | 17 | Sistema agroforestal | 1.45 | 1.22 | 0.979 | 0.998 |
| 25 | 21 | Sistema agroforestal | 1.03 | 1.07 | 0.975 | 0.994 |
| 26 | 22 | Sistema agroforestal | 1.40 | 1.45 | 0.976 | 1 |
| 27 | 25 | Sistema agroforestal | 1.40 | 1.76 | 0.979 | 0.995 |
| 28 | 28 | Sistema agroforestal | 1.24 | 1.01 | 0.986 | 0.990 |
| 29 | 29 | Sistema agroforestal | 1.09 | 1.03 | 0.998 | 1 |
| 30 | 32 | Sistema agroforestal | 1.04 | 1 | 0.995 | 1 |
| 31 | 33 | Sistema agroforestal | 1.25 | 1.19 | 0.997 | 1 |
| 32 | 1 | Vegetación secundaria joven | 1.42 | 1.37 | 1 | 1 |
| 33 | 2 | Vegetación secundaria joven | 1.8 | 2.47 | 0.984 | 1 |
| 34 | 3 | Vegetación secundaria joven | 1.08 | 1.30 | 0.984 | 0.977 |
| 35 | 6 | Vegetación secundaria joven | 1.23 | 1.18 | 0.991 | 0.998 |
| 36 | 16 | Vegetación secundaria joven | 1.20 | 1.01 | 0.987 | 0.957 |
| 37 | 23 | Vegetación secundaria joven | 1.09 | 1.09 | 1 | 1 |
| 38 | 24 | Vegetación secundaria joven | 1.83 | 2.08 | 0.989 | 1 |
| 39 | 26 | Vegetación secundaria joven | 1.68 | 1.28 | 0.979 | 0.979 |
| 40 | 30 | Vegetación secundaria joven | 1.02 | 2.13 | 0.948 | 0.952 |
| 41 | 31 | Vegetación secundaria joven | 1.02 | 1.03 | 0.987 | 0.983 |
| 42 | 3 | Vegetación secundaria madura | 1.08 | 1.13 | 0.984 | 0.975 |
| 43 | 4 | Vegetación secundaria madura | 1.04 | 1.41 | 0.996 | 0.873 |
| 44 | 5 | Vegetación secundaria madura | 1.17 | 1.25 | 0.996 | 0.998 |
| 45 | 6 | Vegetación secundaria madura | 1.07 | 1.03 | 0.991 | 0.992 |
| 46 | 18 | Vegetación secundaria madura | 1.17 | 1.78 | 0.820 | 0.999 |
| 47 | 19 | Vegetación secundaria madura | 1.01 | 1.66 | 0.988 | 0.986 |
| 48 | 20 | Vegetación secundaria madura | 1.04 | 1.15 | 0.977 | 0.974 |
| 49 | 24 | Vegetación secundaria madura | 1.28 | 1.28 | 0.989 | 0.991 |
| 50 | 25 | Vegetación secundaria madura | 1.48 | 1.87 | 0.979 | 0.994 |
| 51 | 27 | Vegetación secundaria madura | 1.21 | 1.09 | 0.998 | 0.967 |
| 52 | 31 | Vegetación secundaria madura | 1.06 | 1.19 | 0.987 | 0.992 |
| 53 | 34 | Vegetación secundaria madura | 1.71 | 1.97 | 0.992 | 0.981 |
Con la rarefacción basada en el número de individuos se estandarizaron 25 de los casos analizados por la abundancia de murciélagos en la selva, por ser este el sitio con menor número de individuos. En 35 casos se encontró una proporción de riqueza mayor para la selva y en 18 casos la selva tuvo una riqueza esperada menor a la del hábitat modificado.
Con la rarefacción por cobertura de la muestra se estandarizaron 29 casos por la selva tropical, por presentar la menor completitud del muestreo. En 38 casos, el número de especies estimado fue mayor para la selva y 15 casos presentaron una riqueza mayor en el hábitat modificado.
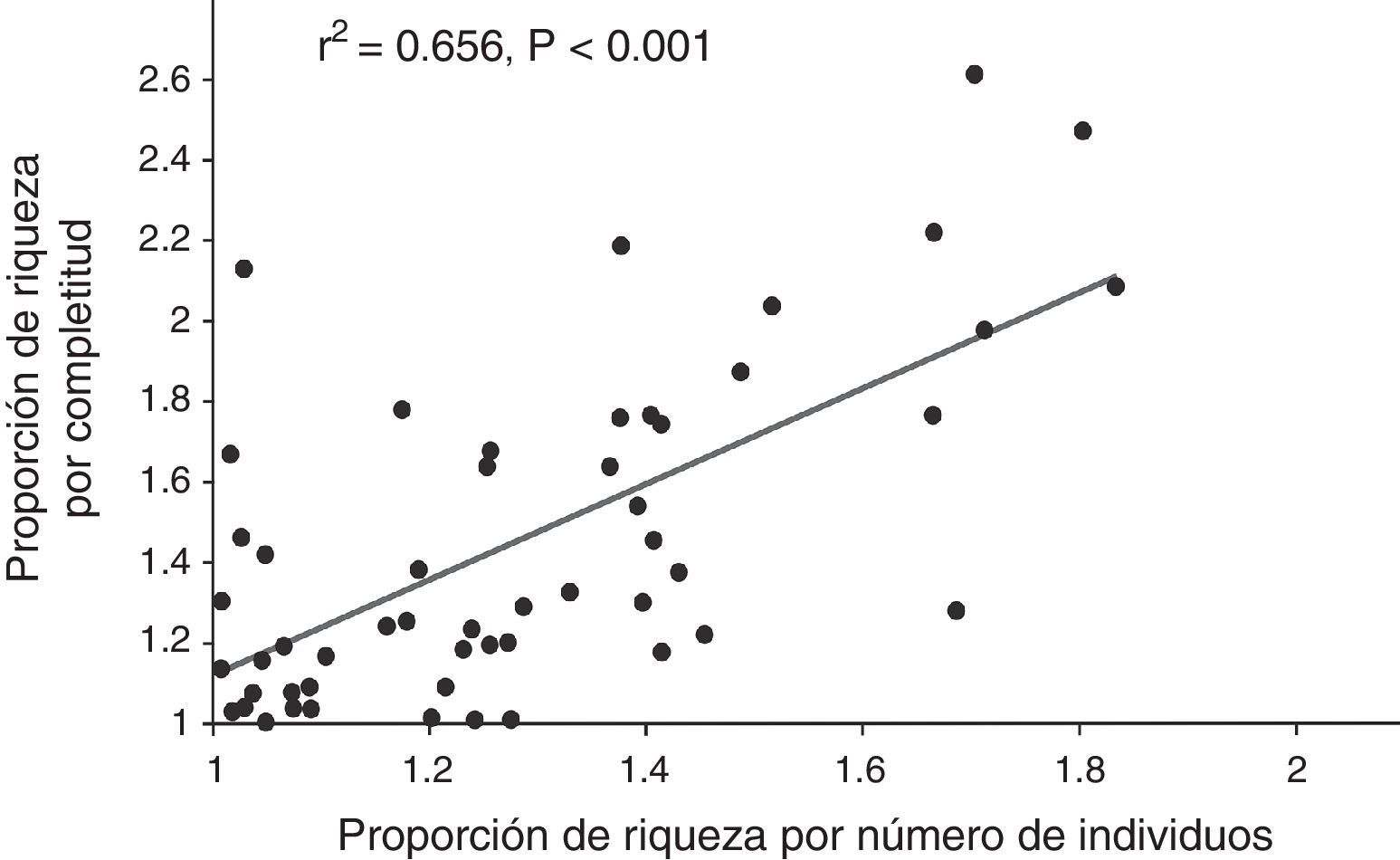
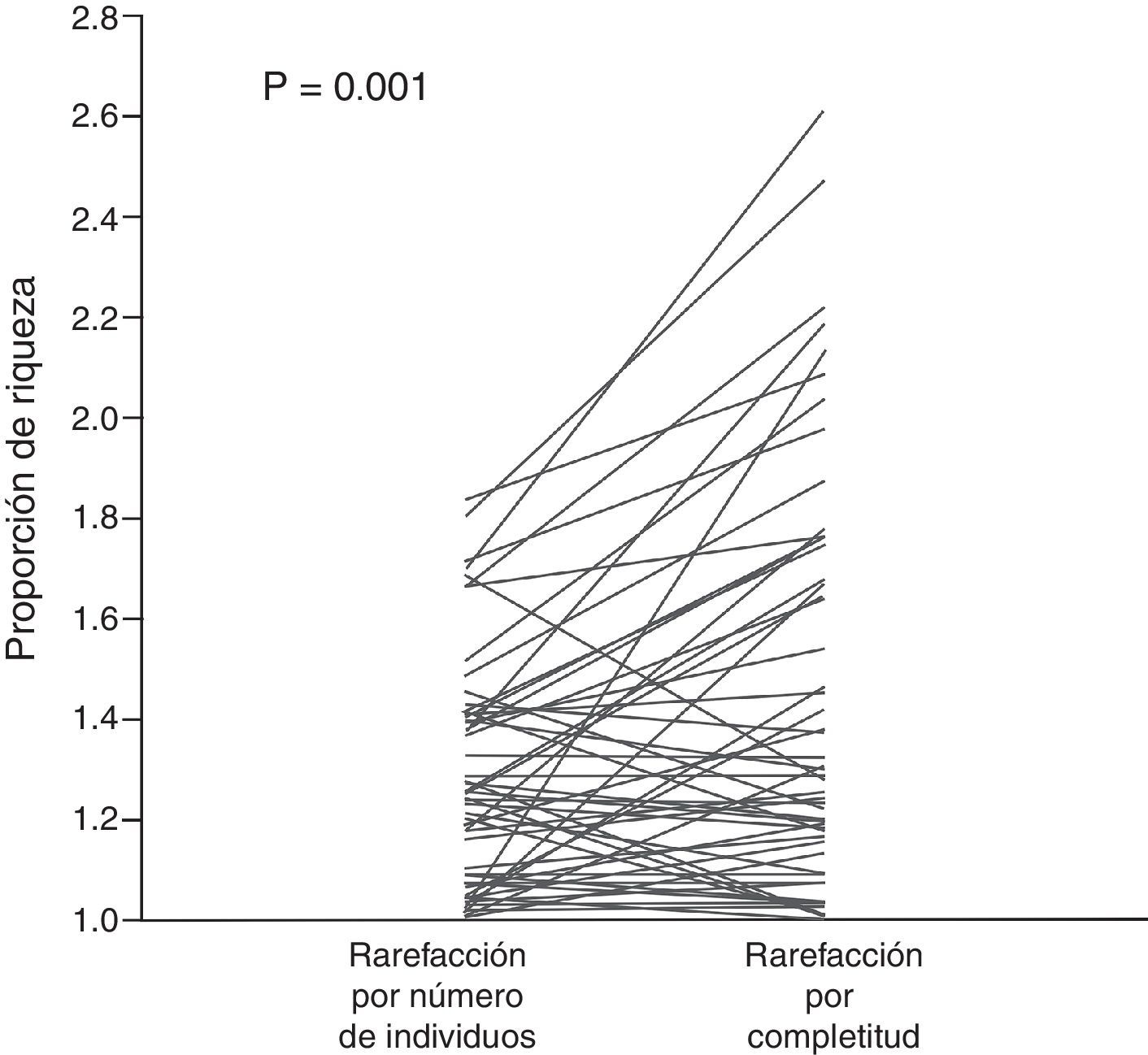
La correlación entre las proporciones de riqueza calculadas con ambos métodos de rarefacción fue positiva y significativa (fig. 1), es decir, ambos métodos mostraron resultados similares en términos de tendencias. Sin embargo, la prueba de signos muestra que la proporción de riqueza calculada por cobertura de muestra (promedio: 1.45, mínimo: 1.01, máximo: 2.61) es significativamente mayor (p<0.05, fig. 2) que la obtenida con el método de rarefacción tradicional (promedio: 1.28, mínimo: 1.01, máximo: 1.83).
 en 53 casos de estudio con murciélagos en el neotrópico.")
 en 53 casos de estudio con murciélagos en el neotrópico.")
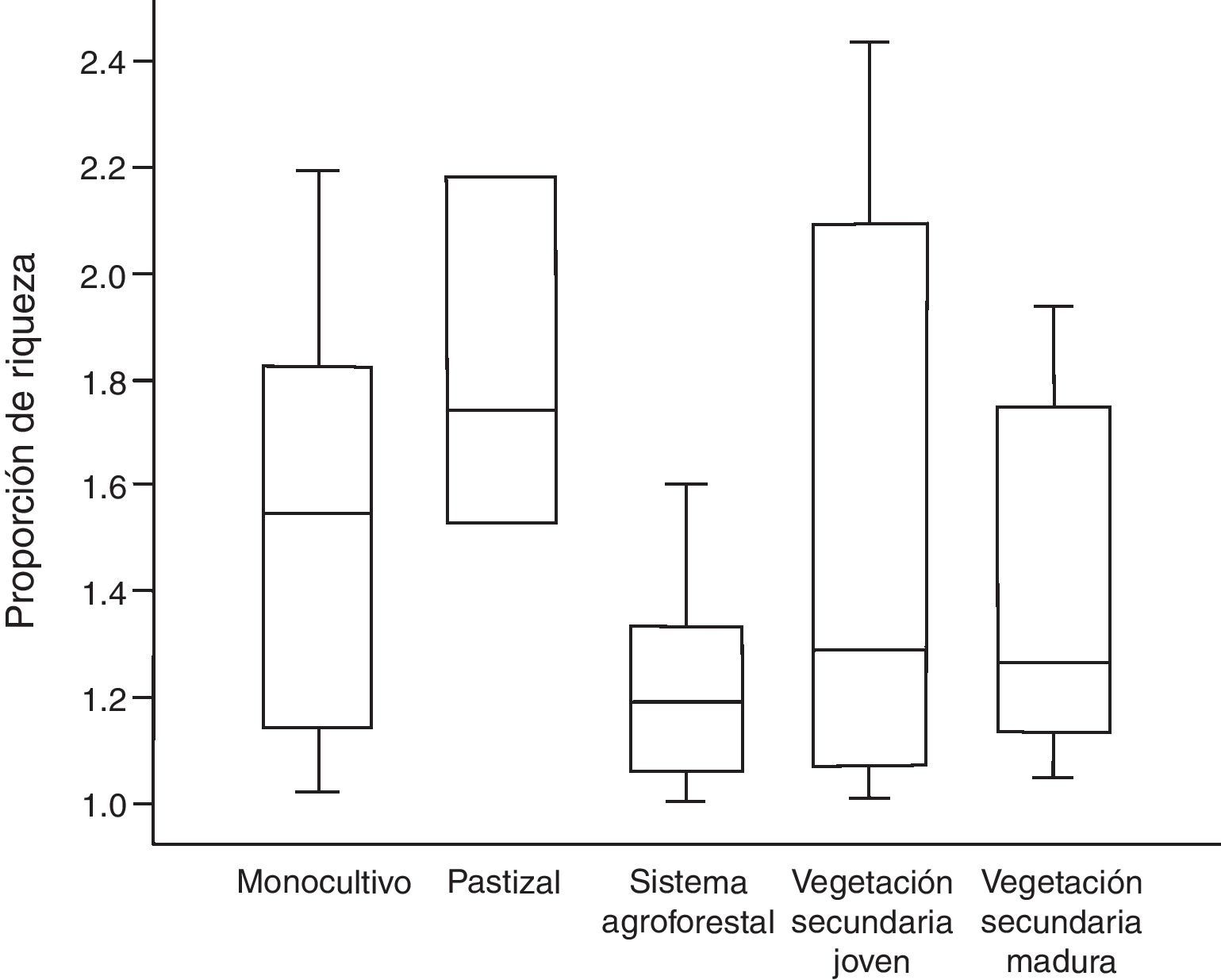
La proporción de riqueza calculada por el método de rarefacción tradicional por número de individuos (tabla 2) no es estadísticamente diferente entre los 5 tipos de hábitat modificado (F4,48=1.015, p=0.409). Sin embargo, la proporción de riqueza con el método de rarefacción por cobertura de la muestra sí es distinta entre los hábitats modificados (H=9.615, g.l.=4, p=0.047, fig. 3). La proporción de riqueza entre selva y los pastizales es mayor (mediana=1.74) que la proporción de riqueza entre selva y sistemas agroforestales (mediana=1.19, fig. 3).
Discusión
Distintos trabajos han utilizado ya el método de rarefacción por cobertura de la muestra propuesto por Chao y Jost (2012), en lugar de la rarefacción tradicional (e.g., Carneiro, Mielke, Casagrande y Fiedler, 2014; Della Roca, Stefanelli, Pasquaretta, Campanaro y Bogliani, 2014; Hernández-Ruedas et al., 2014; Müller, Jarzabek-Muller y Bussler, 2013; Pinzón, Spence y Langor, 2013). No obstante, estos trabajos se limitan a estimar los valores de riqueza, sin utilizar una de las ventajas más importantes de este método: que expresa de manera menos sesgada la magnitud de las diferencias en la riqueza entre las comunidades (Chao y Jost, 2012). Esto ocurre porque la riqueza estandarizada por cobertura de la muestra cumple con el principio de replicación: si se tienen N comunidades con el mismo nu¿mero de especies, sin ninguna especie compartida entre ellas, entonces la riqueza del conjunto de estas comunidades debe ser N veces la riqueza de una sola comunidad (Jost, 2007; Chao y Jost, 2012). La riqueza calculada por el método de rarefacción tradicional no mantiene el principio de replicación, y por lo tanto, comprime la proporción de riqueza entre comunidades, es decir, la diferencia en los valores de riqueza es menor que la real (véase la demostración matemática en Chao y Jost, 2012).
Nuestros resultados muestran que con ambos métodos se obtienen tendencias semejantes en la riqueza estimada. Sin embargo, cuando evaluamos la magnitud del cambio en la riqueza estimada, encontramos que la proporción de riqueza entre comunidades es mayor cuando las muestras se estandarizan con el método de rarefacción basado en la cobertura de la muestra. Esto concuerda con la descripción matemática propuesta por Chao y Jost (2012), quienes aseguran que al estandarizar por cobertura de la muestra se obtiene una proporción de riqueza con menos compresión, o sin reducción alguna, aun para muestras pequeñas, lo que no puede lograrse con la rarefacción tradicional.
El método de rarefacción tradicional basado en el número de individuos ha representado por décadas una herramienta popular para comparar la riqueza de especies, a pesar de tener muchos sesgos (Chiarucci, Bacaro, Rochini y Fattorini, 2008). Por ejemplo, la rarefacción subestima la riqueza cuando las muestras tienen diferencias considerables en la distribución de abundancias entre las especies (Kraker-Castañeda y Cóbar-Carranza, 2011). Es el caso de los trópicos, donde la diversidad de especies es alta y hay muchas especies raras (Gotelli y Colwell, 2011). Por otro lado, la rarefacción sobreestima la riqueza cuando la muestra proviene de una comunidad con distribución agregada, debido a que se obtienen muestras pequeñas con más frecuencia (Colwell y Coddington, 1994). Sin embargo, con la rarefacción por cobertura de la muestra el grado de completitud de las muestras se establece y se estandariza a un mismo nivel para todas las comunidades, evitando los sesgos por la distribución de las abundancias (Chao y Jost, 2012).
Nuestros resultados corroboran que la riqueza de especies en las comunidades de murciélagos no tiene una respuesta consistente al cambio de uso de suelo, como se ha visto en trabajos recientes (Clarke, Pio y Racey, 2005; Cruz-Lara, Lorenzo, Soto, Naranjo y Ramírez-Marcial, 2004; García-Morales et al., 2013; Montaño-Centellas et al., 2015). En el 66% de los casos analizados en este trabajo la riqueza fue menor en los hábitats modificados, especialmente en ambientes deforestados como los pastizales ganaderos. Es posible que las diferencias entre los ambientes se deba a que tradicionalmente se utiliza como unidad de observación el hábitat (bosque, cultivo, etc.), aunque la composición y la configuración del paisaje puede explicar mucho mejor la riqueza que se detecta (Arroyo-Rodríguez, Moreno y Galán-Acedo, en prensa).
La estandarización de la riqueza utilizando el método de rarefacción con base en la cobertura de la muestra, como se muestra en este trabajo, permite evaluar con mayor precisión los cambios que sufren las comunidades en términos de riqueza, especialmente en respuesta a los cambios derivados de las actividades humanas. Además, al priorizar el nivel de completitud es posible diseñar muestreos flexibles más eficientes, sin la necesidad de igualar el esfuerzo de muestreo entre comunidades, sobre todo cuando la asignación del tiempo y los recursos son limitados. Finalmente, la combinación de interpolación y extrapolación resulta de gran utilidad para grupos hiperdiversos, y con potencial de aplicación en patrones ecológicos y biogeográficos de la riqueza de especies, así como en biología de la conservación.
Este trabajo es parte de la tesis de Maricela López-Mejía, estudiante de la Maestría en Ciencias en Biodiversidad y Conservación, por lo que se agradece al Consejo Nacional de Ciencia y Tecnología (Conacyt) la beca otorgada (367362) y al proyecto 161702 de apoyo a la infraestructura de posgrado. Este estudio forma parte del proyecto SEP-Conacyt de Ciencia Básica 222632 «Evaluación de la diversidad de especies mediante el análisis e integración de elementos ecológicos, funcionales y evolutivos».
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.