






La nochebuena (Euphorbia pulcherrima) es una de las plantas de ornato más importantes en el mundo, tanto económica como culturalmente, por ser el símbolo floral de la Navidad. Sin embargo, su domesticación, diversidad genética e historia han sido poco estudiadas. En este trabajo se analizó la diversidad genética y las relaciones de parentesco de nochebuenas de parques y jardines del Distrito Federal, México, a fin de evaluar la diversidad genética e identificar la procedencia de las nochebuenas sembradas en esta zona urbana. Las plantas del Distrito Federal representan 2 variantes genéticas o haplotipos. Una de estas variantes proviene del estado de Guerrero, mientras que la otra pertenece a cultivares mexicanos cercanamente emparentados a las poblaciones silvestres de Guerrero y Morelos. Documentos históricos señalan que desde la época prehispánica las nochebuenas se cultivaban en el centro de México, y en un manuscrito de 1801 se ilustran nochebuenas en los jardines de la Nueva España con modificaciones con respecto a las silvestres. El reconocimiento de la diversidad genética de plantas cultivadas, así como la inferencia de su posible origen geográfico, permitirá la protección, manejo sostenible, conservación y mejoramiento genético del germoplasma mexicano de nochebuena.
Poinsettia (Euphorbia pulcherrima) is one of the most economically and culturally important ornamental plants in the world because it is the floral symbol of Christmas. However, little is known about its domestication, genetic diversity and history. This paper analyses the genetic diversity of plants grown in gardens and parks of the Distrito Federal, Mexico, and we compare it with that found in wild populations to document the genetic diversity of poinsettia represented in the Distrito Federal and its relationship to the plant's origin. The plants found in the Distrito Federal are 2 genetic variants or haplotypes. One of these variants belongs to the state of Guerrero and the other belongs to a variant closely related to wild populations of Guerrero and Morelos. Historical documents show that since the Pre-Hispanic era, poinsettias were grown in central Mexico and in a manuscript from 1801 poinsettias are illustrated in the gardens of New Spain with modifications as compared to wild plants. The recognition of the genetic diversity of cultivated plants will allow the protection, sustainable management, conservation and genetic improvement of Mexican poinsettia germplasm.
La mayoría de los estudios de domesticación se han realizado en plantas con valor alimenticio (e. g.Burger, Chapman y Burke, 2008; Morrell, Buckler y Ross-Ibarra, 2012; Zeder, Emshwiller, Smith y Bradley, 2006), pero muy poco se ha estudiado la domesticación de plantas ornamentales (Blackman et al., 2011; Malécot, Marcussen, Munzinger, Yockteng y Henry, 2007). Entre los pocos estudios al respecto con plantas ornamentales están los casos Dahlia (Hansen y Hjerting, 2000; Mera y Bye, 2006) y Tagetes (Serrato-Cruz, 2004; Serrato-Cruz, Núñez-Colín, Santos-Cortés, Luna-Morales, Martínez-Solís y Cuevas-Sánchez, 2011), en los que se documentan datos históricos y morfológicos. Sin embargo, para estos 2 géneros los estudios de diversidad genética y de relaciones de parentesco entre las plantas silvestres y cultivadas son escasos. Otro claro ejemplo es la nochebuena, una de las plantas más emblemáticas de México.
La nochebuena (Euphobia pulcherrima [E. pulcherrima] Willd. ex Klotzsch) es una de las plantas de ornato de mayor importancia económica en el mundo. Sus ventas anuales superan los 100 millones de dólares en los Estados Unidos, debido a que es el símbolo floral de la Navidad (Taylor, López, Currey y Janick, 2011USDA y NASS, 2011). En 2012, la actividad económica vinculada con esta especie produjo más de 3,000 empleos directos y condujo a la venta de 20 millones de plantas, equivalente a 700 millones de pesos. En México, Morelos es el estado con la mayor producción de nochebuenas, generando el 41.1% de la producción nacional total (Sagarpa, 2012). A pesar de la gran relevancia de la nochebuena, muy poco se ha investigado sobre su domesticación (Fry, 1994; Taylor et al., 2011), variación genética (Ling, Sauve y Gawel, 1997; Su, Jiang, Feng, Li y Tao, 2010; Trejo et al., 2012), distribución (Mayfield, 1997; Trejo et al., 2012), estado de conservación y origen de sus cultivares –variantes de plantas genéticamente estables modificadas fuera de su ambiente natural– (Fry, 1994; Trejo et al., 2012) (Ecke, Matkin y Hartley, 1990).
Las plantas silvestres de nochebuena crecen de manera natural en los bosques tropicales subcaducifolios de la costa del Pacífico, desde Sinaloa, en México, hasta Guatemala (fig. 1A). También se distribuyen en bosques tropicales secos de la cuenca del Balsas, particularmente en el norte de Guerrero y en Morelos (Mayfield, 1997; Trejo et al., 2012). La nochebuena era cultivada desde la época prehispánica en lo que actualmente es la Ciudad de México (Navarro, 1992; Sahagún, 1979; fig. 1B), teniendo una larga tradición hortícola que se remonta a los tiempos del imperio Azteca (Hernández, 1946; Navarro, 1992; Sahagún, 1979). Esta especie era llamada cuetlaxochitl, que en náhuatl significa «flor que se marchita» (Hernández, 1946; Sahagún, 1979). La flor se consideraba símbolo de la pureza (McGinty, 1980) y se utilizaba para tratar algunas afecciones de la piel, empleando las hojas como cataplasmas (Argueta, Cano y Rodarte, 1994; Hernández, 1946). La nochebuena era cultivada en los jardines de Nezahualcóyotl y Moctezuma (McGinty, 1980). Fue incorporada por los frailes franciscanos de Taxco a la celebración del nacimiento de Jesús, debido a que su época de floración coincide con dicha festividad, y a sus colores brillantes (Ecke et al., 1990; McGinty, 1980). En la actualidad, la nochebuena, que es la planta ornamental de interior de mayor demanda en el invierno, es considerada la flor de la amistad en algunos países, como España (Ecke, 2011; Taylor et al., 2011).
 analizadas en este estudio. A, recolectas de poblaciones silvestres ubicadas sobre la distribución potencial de E. pulcherrima (Trejo et al., 2012); B, recolectas de plantas ubicadas en el Distrito Federal; C, fenotipo de inflorescencia de planta silvestre nombrada como «simple»; D, fenotipo de inflorescencia de planta cultivada nombrada como «doble».")
Recolectas de plantas silvestres y cultivadas de Euphorbia pulcherrima (E. pulcherrima) analizadas en este estudio. A, recolectas de poblaciones silvestres ubicadas sobre la distribución potencial de E. pulcherrima (Trejo et al., 2012); B, recolectas de plantas ubicadas en el Distrito Federal; C, fenotipo de inflorescencia de planta silvestre nombrada como «simple»; D, fenotipo de inflorescencia de planta cultivada nombrada como «doble».
La introducción de la nochebuena a la horticultura internacional ha sido tradicionalmente atribuida a Joel Roberts Poinsett, Primer Ministro Plenipotenciario de los Estados Unidos en México, de 1825-1830 (Ecke et al., 1990; Fry, 1994; Graham, 1836a; McGinty, 1980; Moon, 1956; Rafinesque, 1833). Se dice que Poinsett encontró la nochebuena en barrancas cercanas a Taxco, Guerrero, en 1828 (Ecke et al., 1990; McGinty, 1980). Poinsett en su calidad de miembro de la Sociedad Filosófica Americana, cuya sede se ubicaba en Filadelfia, realizó una expedición a distintos lugares de México, acompañado por 3 miembros de la Sociedad Histórica de Pensilvania: William Maclure, Thomas Say y William H. Keating, en 1828 y 1829. Poinsett y sus acompañantes enviaron desde México al Jardín Botánico de Bartram en Filadelfia al menos 4 cargamentos de plantas en el curso de esos años, entre las cuales pudieron haberse encontrado los ejemplares de nochebuena (Fry, 1994; Ronaldson, 1828; Say, 1828). Debido a que dichos ejemplares no estaban bien etiquetados, se desconoce quiénes los recolectaron. En una carta, James Ronaldson le escribió a Poinsett que desconoce quién mandó las cajas que recibieron en Bartram, pero que suponía que eran de él (Poinsett) y William H. Keating (Ronaldson, 1828). Por consiguiente, los sitios de los cuales procedían las nochebuenas que posiblemente llegaron a Bartram y quién o quiénes las recolectaron siguen sin conocerse.
Una vez que la nochebuena llegó a los Estados Unidos se comenzó a escribir sobre la peculiar planta de flores rojas proveniente de México. Graham (1836a) menciona que en 1828 Poinsett envió plantas y semillas de nochebuena al Jardín Botánico de Bartram en Filadelfia (Rafinesque, 1833) y a la casa de Poinsett en Charleston, Carolina del Sur. Fry (1994) indica que la aclimatación de las plantas y su cultivo se dio en el Jardín Botánico de Bartram y estuvo a cargo de la nieta de John Bartram, Ann Bartram, y del esposo de la misma, Robert Carr. En 1929, los Bartram presentaron plantas de nochebuena, a la cual llamaron Poinsettia pulcherrima en reconocimiento a Poinsett, en la primera exposición semianual de frutos, flores y plantas de la Sociedad de Horticultura de Pensilvania (Fry, 1994), evento que se conoce hoy en día como «Feria de Flores de Filadelfia» (www.bartramsgarden.org). Pocos años después de la introducción de la nochebuena en Filadelfia, las casas cercanas a Bartram comenzaron a presentar nochebuenas en sus invernaderos (Rafinesque, 1833). En 1833, Robert Buist introdujo las nochebuenas cultivadas por los Bartram al Real Jardín Botánico de Edimburgo (Graham, 1836a). Lamentablemente, en la actualidad no hay plantas vivas ni ejemplares de herbario de las nochebuenas enviadas por Poinsett en colección alguna del Jardín Botánico de Bartram (Fry, com. pers.) o en el Real Jardín Botánico de Edimburgo.
Las nochebuenas introducidas a los Estados Unidos y al resto del mundo han sido modificadas por manejo local, mejoramiento genético y biotecnología, lo que ha dado origen a más de 300 cultivares (Ecke, 2011; Ecke et al., 1990; Fry, 1994; Graham, 1836a; Taylor et al., 2011). ¿Cuál es la historia de los cultivares de nochebuena en México? ¿La nochebuena fue modificada en México antes de ser introducida a los Estados Unidos? El análisis de las relaciones de parentesco entre las poblaciones silvestres de nochebuena y los cultivares sembrados en México podría aportar indicios para responder a estas preguntas.
En una investigación previa se encontró que los cultivares comerciales de nochebuena presentan únicamente 2 variantes genéticas de cloroplasto, el haplotipo 5 y 7 (Trejo et al., 2012). Una de estas variantes (haplotipo 5) está presente también en los cultivares extranjeros y en poblaciones silvestres del occidente y del norte de Guerrero. La segunda variante (haplotipo 7) está presente en los cultivares mexicanos, pero no se ha encontrado en población silvestre alguna (Trejo et al., 2012). Sin embargo, la segunda variante podría ser del centro de México, puesto que está cercanamente relacionada con otras variantes de la misma región (Trejo et al., 2012). En una pequeña muestra de plantas sembradas en jardines y parques del Distrito Federal, México, fue encontrado el haplotipo 7 en plantas con apariencia silvestre y en cultivares (Trejo y Olson, en prensa). Por lo tanto, el haplotipo 7 podría ser distintivo de poblaciones silvestres aún no encontradas del centro de México y plantas modificadas conscientemente por el hombre. Desde la época prehispánica, la horticultura se practicaba en los jardines y traspatios de las casas o en los jardines botánicos de los emperadores (Granziera, 2001; McGinty, 1980). Dada esta tradición centenaria, podríamos encontrar evidencias del manejo prehispánico de la nochebuena.
Para analizar la diversidad genética de la nochebuena en zonas urbanas, así como para inferir la probable procedencia de las plantas cultivadas y del haplotipo 7, realizamos un muestreo extenso de plantas de nochebuenas sembradas en parques y jardines del Distrito Federal. El análisis de la diversidad genética de E. pulcherrima y la determinación de la procedencia geográfica de los cultivares permitirá impulsar el desarrollo de nuevos cultivares mexicanos, conservar el germoplasma mexicano de nochebuena y avanzar en el conocimiento del manejo y domesticación de plantas ornamentales.
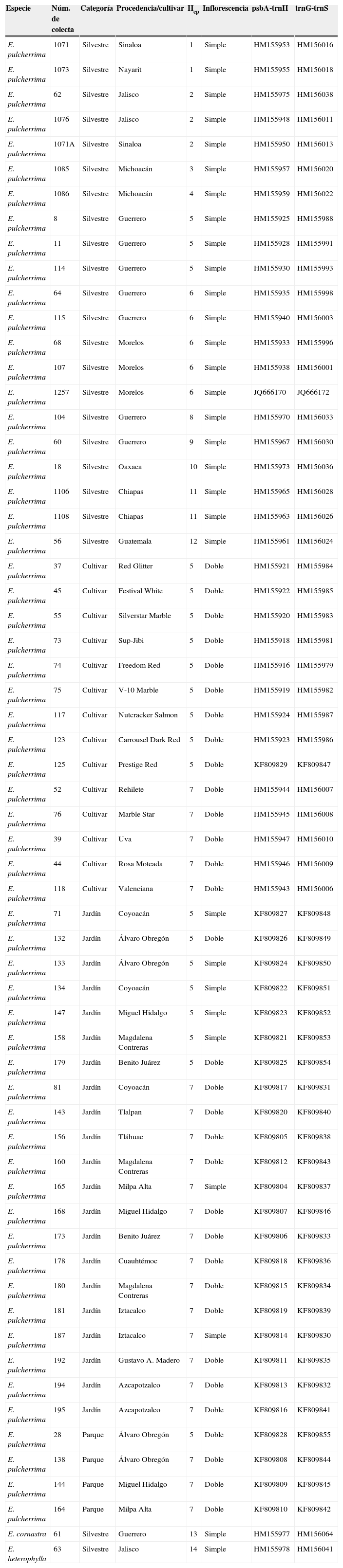
Materiales y métodosPlantas analizadas. Uno de los principales efectos de la selección artificial en E. pulcherrima se observa en la inflorescencia. Al mismo tiempo que las plantas silvestres y algunos cultivares presentan un conjunto de flores asociadas a un solo verticilo de brácteas –conjunto de hojas de colores que brotan a un mismo nivel– (fig. 1C), otros cultivares exhiben 2 o más (fig. 1D). En este estudio fueron analizadas 21 plantas provenientes de poblaciones silvestres de E. pulcherrima de México y Guatemala (fig. 1A), 14 cultivares de México y los Estados Unidos, y 25 plantas del Distrito Federal (fig. 1B) con un solo verticilo de brácteas «simples», o con 2 «dobles» (fig. 1C-D). Debido a que en estudios previos (Trejo et al., 2012; Trejo y Olson, en prensa) no se encontró más de una variante genética dentro de las poblaciones silvestres o cultivar, solo se analizó un individuo por cada grupo (tabla 1). Utilizamos como grupos externos a E. cornastra Dressler por ser la especie hermana de E. pulcherrima y E. heterophylla L. por ser otro miembro del subgénero Poinsettia, según resultados obtenidos en análisis filogenéticos previos (Mayfield, 1997; Steinmann y Porter, 2002; Zimmermann, Ritz y Hellwing, 2010).
Plantas de Euphorbia analizadas en este estudio. Se presenta la especie, el número de colecta, la procedencia en México, con excepción de Sacatepéquez que pertenece a Guatemala, el número de haplotipo de cloroplasto (Hcp), tipo de inflorescencia y número de accesión en GenBank siguiendo la numeración de Trejo et al. (2012).
| Especie | Núm. de colecta | Categoría | Procedencia/cultivar | Hcp | Inflorescencia | psbA-trnH | trnG-trnS |
|---|---|---|---|---|---|---|---|
| E. pulcherrima | 1071 | Silvestre | Sinaloa | 1 | Simple | HM155953 | HM156016 |
| E. pulcherrima | 1073 | Silvestre | Nayarit | 1 | Simple | HM155955 | HM156018 |
| E. pulcherrima | 62 | Silvestre | Jalisco | 2 | Simple | HM155975 | HM156038 |
| E. pulcherrima | 1076 | Silvestre | Jalisco | 2 | Simple | HM155948 | HM156011 |
| E. pulcherrima | 1071A | Silvestre | Sinaloa | 2 | Simple | HM155950 | HM156013 |
| E. pulcherrima | 1085 | Silvestre | Michoacán | 3 | Simple | HM155957 | HM156020 |
| E. pulcherrima | 1086 | Silvestre | Michoacán | 4 | Simple | HM155959 | HM156022 |
| E. pulcherrima | 8 | Silvestre | Guerrero | 5 | Simple | HM155925 | HM155988 |
| E. pulcherrima | 11 | Silvestre | Guerrero | 5 | Simple | HM155928 | HM155991 |
| E. pulcherrima | 114 | Silvestre | Guerrero | 5 | Simple | HM155930 | HM155993 |
| E. pulcherrima | 64 | Silvestre | Guerrero | 6 | Simple | HM155935 | HM155998 |
| E. pulcherrima | 115 | Silvestre | Guerrero | 6 | Simple | HM155940 | HM156003 |
| E. pulcherrima | 68 | Silvestre | Morelos | 6 | Simple | HM155933 | HM155996 |
| E. pulcherrima | 107 | Silvestre | Morelos | 6 | Simple | HM155938 | HM156001 |
| E. pulcherrima | 1257 | Silvestre | Morelos | 6 | Simple | JQ666170 | JQ666172 |
| E. pulcherrima | 104 | Silvestre | Guerrero | 8 | Simple | HM155970 | HM156033 |
| E. pulcherrima | 60 | Silvestre | Guerrero | 9 | Simple | HM155967 | HM156030 |
| E. pulcherrima | 18 | Silvestre | Oaxaca | 10 | Simple | HM155973 | HM156036 |
| E. pulcherrima | 1106 | Silvestre | Chiapas | 11 | Simple | HM155965 | HM156028 |
| E. pulcherrima | 1108 | Silvestre | Chiapas | 11 | Simple | HM155963 | HM156026 |
| E. pulcherrima | 56 | Silvestre | Guatemala | 12 | Simple | HM155961 | HM156024 |
| E. pulcherrima | 37 | Cultivar | Red Glitter | 5 | Doble | HM155921 | HM155984 |
| E. pulcherrima | 45 | Cultivar | Festival White | 5 | Doble | HM155922 | HM155985 |
| E. pulcherrima | 55 | Cultivar | Silverstar Marble | 5 | Doble | HM155920 | HM155983 |
| E. pulcherrima | 73 | Cultivar | Sup-Jibi | 5 | Doble | HM155918 | HM155981 |
| E. pulcherrima | 74 | Cultivar | Freedom Red | 5 | Doble | HM155916 | HM155979 |
| E. pulcherrima | 75 | Cultivar | V-10 Marble | 5 | Doble | HM155919 | HM155982 |
| E. pulcherrima | 117 | Cultivar | Nutcracker Salmon | 5 | Doble | HM155924 | HM155987 |
| E. pulcherrima | 123 | Cultivar | Carrousel Dark Red | 5 | Doble | HM155923 | HM155986 |
| E. pulcherrima | 125 | Cultivar | Prestige Red | 5 | Doble | KF809829 | KF809847 |
| E. pulcherrima | 52 | Cultivar | Rehilete | 7 | Doble | HM155944 | HM156007 |
| E. pulcherrima | 76 | Cultivar | Marble Star | 7 | Doble | HM155945 | HM156008 |
| E. pulcherrima | 39 | Cultivar | Uva | 7 | Doble | HM155947 | HM156010 |
| E. pulcherrima | 44 | Cultivar | Rosa Moteada | 7 | Doble | HM155946 | HM156009 |
| E. pulcherrima | 118 | Cultivar | Valenciana | 7 | Doble | HM155943 | HM156006 |
| E. pulcherrima | 71 | Jardín | Coyoacán | 5 | Simple | KF809827 | KF809848 |
| E. pulcherrima | 132 | Jardín | Álvaro Obregón | 5 | Doble | KF809826 | KF809849 |
| E. pulcherrima | 133 | Jardín | Álvaro Obregón | 5 | Simple | KF809824 | KF809850 |
| E. pulcherrima | 134 | Jardín | Coyoacán | 5 | Simple | KF809822 | KF809851 |
| E. pulcherrima | 147 | Jardín | Miguel Hidalgo | 5 | Simple | KF809823 | KF809852 |
| E. pulcherrima | 158 | Jardín | Magdalena Contreras | 5 | Simple | KF809821 | KF809853 |
| E. pulcherrima | 179 | Jardín | Benito Juárez | 5 | Doble | KF809825 | KF809854 |
| E. pulcherrima | 81 | Jardín | Coyoacán | 7 | Doble | KF809817 | KF809831 |
| E. pulcherrima | 143 | Jardín | Tlalpan | 7 | Doble | KF809820 | KF809840 |
| E. pulcherrima | 156 | Jardín | Tláhuac | 7 | Doble | KF809805 | KF809838 |
| E. pulcherrima | 160 | Jardín | Magdalena Contreras | 7 | Doble | KF809812 | KF809843 |
| E. pulcherrima | 165 | Jardín | Milpa Alta | 7 | Simple | KF809804 | KF809837 |
| E. pulcherrima | 168 | Jardín | Miguel Hidalgo | 7 | Doble | KF809807 | KF809846 |
| E. pulcherrima | 173 | Jardín | Benito Juárez | 7 | Doble | KF809806 | KF809833 |
| E. pulcherrima | 178 | Jardín | Cuauhtémoc | 7 | Doble | KF809818 | KF809836 |
| E. pulcherrima | 180 | Jardín | Magdalena Contreras | 7 | Doble | KF809815 | KF809834 |
| E. pulcherrima | 181 | Jardín | Iztacalco | 7 | Doble | KF809819 | KF809839 |
| E. pulcherrima | 187 | Jardín | Iztacalco | 7 | Simple | KF809814 | KF809830 |
| E. pulcherrima | 192 | Jardín | Gustavo A. Madero | 7 | Doble | KF809811 | KF809835 |
| E. pulcherrima | 194 | Jardín | Azcapotzalco | 7 | Doble | KF809813 | KF809832 |
| E. pulcherrima | 195 | Jardín | Azcapotzalco | 7 | Doble | KF809816 | KF809841 |
| E. pulcherrima | 28 | Parque | Álvaro Obregón | 5 | Doble | KF809828 | KF809855 |
| E. pulcherrima | 138 | Parque | Álvaro Obregón | 7 | Doble | KF809808 | KF809844 |
| E. pulcherrima | 144 | Parque | Miguel Hidalgo | 7 | Doble | KF809809 | KF809845 |
| E. pulcherrima | 164 | Parque | Milpa Alta | 7 | Doble | KF809810 | KF809842 |
| E. cornastra | 61 | Silvestre | Guerrero | 13 | Simple | HM155977 | HM156064 |
| E. heterophylla | 63 | Silvestre | Jalisco | 14 | Simple | HM155978 | HM156041 |
En invierno del 2011 y 2012 recolectamos 25 nochebuenas en parques y jardines privados del Distrito Federal. Se pretendió recolectar en cada una de las 16 delegaciones una nochebuena de inflorescencia simple y una con inflorescencia doble. Sin embargo, no en todas ellas encontramos plantas que exhibieran las 2 variantes y en algunas delegaciones no encontramos nochebuenas en parques o jardines (fig. 1). Por consiguiente, nuestro muestreo final consistió en 21 plantas de jardín y 4 de parque, provenientes de 12 delegaciones. Siete de las plantas de jardín (33.3%) presentaron inflorescencias simples, el resto dobles (66.6%), y las nochebuenas de los parques presentaron inflorescencias dobles (tabla 1; fig. 1).
Secuenciación y análisis de la variación genética. La extracción de ADN, las reacciones de amplificación y la secuenciación de 2 espaciadores intergénicos no codificantes de cloroplasto psbA-trnH (Sang, Crawford y Stuessy, 1997) y trnS(GCU)-trnG(UCC) (Hamilton, 1999) fueron llevadas a cabo como se describe en Trejo et al. (2012). Para efectuar los análisis de variación genética se estimó el número de haplotipos, la diversidad haplotípica (Hd) (Nei, 1987), la diversidad genética de Watterson θ (θw) (Watterson, 1975), la diversidad nucleotídica (π o pi) (Nei, 1987), el número de sitios polimórficos (S) y el número promedio de diferencias pareadas (k) (Tajima, 1983), utilizando DnaSP v. 5.0 (Librado y Rozas, 2009).
Relaciones entre plantas silvestres y cultivares. Para describir el patrón de relaciones de parentesco existente entre los individuos de E. pulcherrima, llevamos a cabo un análisis bayesiano. Seleccionamos el modelo de evolución que mejor se ajustara a nuestros datos utilizando el criterio de información de Akaike (Akaike, 1974) implementado en el program jModeltest 2.1.3 (Darriba, Taboada, Doallo y Posada, 2012). Realizamos un análisis conjunto de los 2 marcadores de cloroplasto en MrBayes v.3.1.2 (Huelsenbeck y Ronquist, 2001). En dicho análisis corrimos 4 cadenas de Markov por 10,000 generaciones, habiendo especificado una temperatura de 0.03. Una vez que concluimos el análisis, confirmamos que las búsquedas hubieran alcanzado la fase estacionaria mediante la observación de gráficas de valores log-likelihood contra generaciones. Descartamos el 25% de los árboles iniciales como burn-in y calculamos el consenso de mayoría con el resto de los árboles. Excluimos la mitad de los árboles antes de calcular las probabilidades posteriores bayesianas (PP).
ResultadosAnálisis de la variación genética. La matriz de las secuencias alineadas del espaciador intergénico de cloroplasto psbA-trnH tuvo una longitud de 716pb, mientras que la de trnS-trnG fue de 838pb. La matriz combinada de los 2 fragmentos tuvo una longitud de 1.554pb. Los números de acceso de GenBank aparecen en la tabla 1 y el alineamiento está disponible en TreeBASE (http://purl.org/phylo/treebase/phylows/study/TB:S14695).
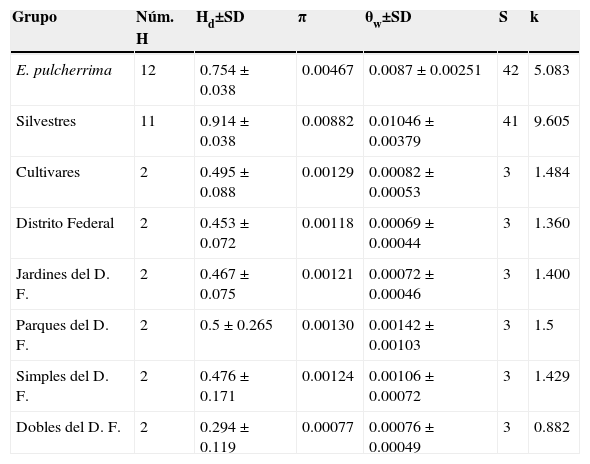
La diversidad genética presente en las plantas del Distrito Federal (Hd=0.453±0.072, π=0.00118) es casi tan elevada como la encontrada en los cultivares (Hd=0.495±0.088, π=0.00129), aunque menor que la observada en las plantas silvestres (Hd=0.914±0.038, π=0.00882). La cantidad de variación genética es similar entre las plantas de jardín (Hd=0.467±0.075, π=0.00121) y las plantas de los parques (Hd=0.5±0.265, π=0.00130). Las plantas con inflorescencia sencilla presentan valores de diversidad genética más elevados (Hd=0.476±0.171, π=0.00124) que las plantas con inflorescencias dobles (Hd=0.294±0.119, π=0.00077) (tabla 2).
Índices de diversidad genética para el análisis de fragmentos de cloroplasto psbA-trnH y trnS-trnG en plantas de Euphorbia pulcherrima.
| Grupo | Núm. H | Hd±SD | π | θw±SD | S | k |
|---|---|---|---|---|---|---|
| E. pulcherrima | 12 | 0.754±0.038 | 0.00467 | 0.0087±0.00251 | 42 | 5.083 |
| Silvestres | 11 | 0.914±0.038 | 0.00882 | 0.01046±0.00379 | 41 | 9.605 |
| Cultivares | 2 | 0.495±0.088 | 0.00129 | 0.00082±0.00053 | 3 | 1.484 |
| Distrito Federal | 2 | 0.453±0.072 | 0.00118 | 0.00069±0.00044 | 3 | 1.360 |
| Jardines del D. F. | 2 | 0.467±0.075 | 0.00121 | 0.00072±0.00046 | 3 | 1.400 |
| Parques del D. F. | 2 | 0.5±0.265 | 0.00130 | 0.00142±0.00103 | 3 | 1.5 |
| Simples del D. F. | 2 | 0.476±0.171 | 0.00124 | 0.00106±0.00072 | 3 | 1.429 |
| Dobles del D. F. | 2 | 0.294±0.119 | 0.00077 | 0.00076±0.00049 | 3 | 0.882 |
H: haplotipo; Hd: diversidad haplotípica; k: promedio de número de diferencias nucleotídicas; S: número de sitios segregantes; SD: desviación estándar; θw: diversidad genética; π: diversidad nucleotídica.
Relaciones genealógicas de las plantas del Distrito Federal. Las plantas del Distrito Federal presentan 2 haplotipos, las variantes 5 y 7. El haplotipo 5 se presenta en los cultivares extranjeros y en las poblaciones de Guerrero, mientras que el haplotipo 7 está presente en los cultivares mexicanos y un cultivar extranjero (fig. 2). No se ha encontrado ninguna población silvestre que tenga el haplotipo 7, pero dada su cercanía genética con los haplotipos de las poblaciones del centro de México podría demostrar que son de esta región geográfica.
. Las probabilidades posteriores se muestran sobre las ramas. J: planta de jardín; p: planta de parque; s: inflorescencia simple; d: inflorescencia doble.")
Cladograma de consenso de mayoría al 50% bayesiano basado en los fragmentos de cloroplasto psbA-trnH y trnG-trnS (1.554pb; K81uf+G). Las probabilidades posteriores se muestran sobre las ramas. J: planta de jardín; p: planta de parque; s: inflorescencia simple; d: inflorescencia doble.
Nochebuenas del Distrito Federal, México. Las nochebuenas del Distrito Federal representan las variantes genéticas tanto de cultivares mexicanos como de extranjeros. La mayoría de estas plantas, dos terceras partes, corresponden al haplotipo 7 y, de estas, el 86% son de inflorescencias dobles. El haplotipo 7 y las inflorescencias dobles son representativos, principalmente, de los cultivares mexicanos. También este haplotipo se observó en el cultivar extranjero MarbleStar, cultivar bicolor de brácteas color salmón y crema que apareció en el mercado en el invierno del 2011 (Ecke, 2011). Por las características de este cultivar, es poco probable que sea una de las plantas que estudiamos en el Distrito Federal, porque las plantas que analizamos en este trabajo fueron de brácteas de color rojo. Es posible que en el Distrito Federal encontremos más cultivares mexicanos debido al cultivo tradicional de la nochebuena en esta región desde hace cientos de años (Hernández, 1946; Sahagún, 1979). Datos históricos señalan que se cultivaban nochebuenas desde la época prehispánica en lo que es hoy el Distrito Federal (Sahagún, 1979), y la presencia de cultivares mexicanos en la Nueva España se observa en la ilustración de Navarro (1992), en la cual se dibuja una nochebuena con inflorescencias de más de una hilera brácteas (fig. 3). Incluso es posible que Joel Roberts Poinsett se haya llevado plantas con cierto grado de manejo, ya que se observan plantas con inflorescencias dobles en ilustraciones de Graham (1836b) (fig. 4). También es posible que existan variantes novedosas de nochebuenas en el Distrito Federal por adaptaciones a las condiciones ambientales de la región, como pudiera ser tolerancia al frío. Por lo anterior, las nochebuenas del Distrito Federal pueden ser germoplasma útil para el mejoramiento genético y la generación de nuevos cultivares mexicanos.
. Ilustración de Euphorbia pulcherrima con inflorescencias dobles.")
Nochebuena nombrada como «paño de grana» en Historia Natural o Jardín Americano del año 1801 (Navarro, 1992). Ilustración de Euphorbia pulcherrima con inflorescencias dobles.
.")
Ilustración botánica de nochebuena descrita como Poinsettia pulcherrima en Graham (1836b).
Movimiento de plantas de nochebuenas silvestres a zonas urbanas. Esquejes, semillas e incluso plantas completas de nochebuenas silvestres han sido sembradas en los jardines de las casas ubicadas en las proximidades de las poblaciones silvestres. En ocasiones, se toman esquejes de estas plantas para sembrarlos en otras casas del mismo poblado o en otros cercanos. Este movimiento de nochebuenas silvestres lo hemos observado en lugares rurales de Sinaloa, Guerrero, Oaxaca y Chiapas. En jardines del Distrito Federal hay plantas de apariencia silvestre, de inflorescencias simples y que comparten la misma variante genética de poblaciones silvestres de Guerrero (haplotipo 5). Esto podría reflejar la costumbre de los habitantes del Distrito Federal de sembrar plantas de otras partes de México.
Variantes silvestres desconocidas o extintas en su hábitat natural presentes en el Distrito Federal. El Distrito Federal podría resguardar variantes genéticas de nochebuenas silvestres que ya no existen en su hábitat natural o de poblaciones silvestres no exploradas. En el Distrito Federal también encontramos plantas de apariencia silvestre con inflorescencias simples y de una variante genética, el haplotipo 7, que no tuvo correspondencia con las poblaciones silvestres estudiadas hasta este momento (Trejo et al., 2012). El haplotipo 7 podría pertenecer al centro de México por su cercanía genética con las poblaciones silvestres de dicha región, en donde la mancha urbana ha crecido enormemente y han desaparecido grandes extensiones de bosques y poblaciones nativas de plantas y animales (Rzedowski y Rzedowski, 2001). Miller y Schaal (2005) observaron algo similar al detectar que algunos individuos de ciruelas silvestres, Spondias purpurea, sembrados en casas a lo largo de la distribución natural de esta especie contaban con variantes silvestres que no encontraron en su hábitat natural. En dado caso, las interacciones ecológicas y la biodiversidad asociadas a las plantas silvestres en su hábitat natural se pierden, y los jardines de las casas y los parques podrían ser el último refugio para algunas variantes genéticas. Finalmente, los resultados de este estudio ilustran la necesidad de estudiar detalladamente el manejo y la domesticación de la nochebuena a lo largo de su distribución natural. Lo anterior permitiría reconocer la procedencia de los cultivares e identificar los procesos de domesticación que les dieron origen, lo que contribuiría a proteger, conservar, manejar e impulsar el desarrollo de nuevos cultivares mexicanos, y evitar la biopiratería del germoplasma mexicano.
A National Geographic Society CRE concesión 870-09 y a Conacyt. Laura Trejo agradece al Posgrado en Ciencias Biológicas la beca Conacyt y la beca ICyTDF. A Joel Fry, Academy of Natural Sciences of Philadelphia, Royal Botanic Garden Edinburgh y Susana Magallón por la información histórica. A Angélica Cervantes, Leonardo Alvarado y Gabriela Montes por sus comentarios al escrito.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.