





La construcción de ciudades constituye la transformación más drástica, fundamental e irreversible de los sistemas naturales, reemplazando todo componente biótico y abiótico original del lugar. Entre los artrópodos que han podido adaptarse a los ambientes urbanos se encuentran las arañas. Se encontraron 41 especies en el interior de 12 casas de 2 niveles de la ciudad de Toluca, ubicadas en 4 ambientes con diferente grado de urbanización, durante el periodo de septiembre del 2009 a agosto del 2010. Por primera vez se empleó un método sistematizado para recolecta de arañas en el interior de las viviendas. La diversidad de arañas fue diferente en cada uno de los ambientes muestreados; el índice de Shannon (H’) mostró que la diversidad de arañas es mayor en las casas que presentan jardín en el ambiente urbano, por lo que no se apoya la hipótesis del disturbio intermedio. El número de arañas encontradas fue mayor en el primer nivel de las viviendas que en el segundo, por lo que se proponen 3 hipótesis para explicar esta diferencia. Se propone la prueba de Olmstead-Tukey para determinar los 4 niveles de sinantropismo (N. S.), ya que la prueba engloba valores utilizados en los índices de densidad e infestación (ocupación).
City-building is the most drastic, fundamental and irreversible transformation of natural environments, replacing all original biotic and abiotic components of the site. Spiders are among the arthropods that have become adapted to urban environments. We found 41 species within 12 two-story houses of the city of Toluca, located in 4 environments with different degrees of urbanization, during the period September 2009-August 2010. For the first time a systematic method was used to collect spiders inside houses with even collecting efforts. The diversity of spiders is different in each of the sampled environments, the Shannon index (H ‘) indicated that the diversity of spiders was higher in houses with gardens in the urban environment, so it does not support the hypothesis of intermediate disturbance. The number of spiders found is higher on the first level of houses than on the second, and 3 hypotheses are proposed to explain this difference. The Olmstead-Tukey test is proposed to determine the 4 levels of synanthropism (N. S.), since the test includes values used in the density and infestation rates (occupation).
La construcción de ciudades constituye la transformación de los sistemas ambientales naturales más drástica, fundamental e irreversible, reemplazando todo componente biótico y abiótico original (Romero et al., 2004). En este nuevo conjunto de componentes instaurado por el ambiente urbano, existen varios efectos que permiten el establecimiento de ciertos grupos o especies de artrópodos, a la asociación entre estos “huéspedes” y las construcciones del hombre se le denomina sinantropismo (McIntyre, 2000; Durán-Barrón et al., 2009).
Los artrópodos son las opciones lógicas para estudiar los efectos de la urbanización, por 5 razones: 1) son diversos y por lo tanto pueden ofrecer una diversidad biológica instantánea de un área en general; 2) sus tiempos de generación son relativamente cortos, lo que significa que pueden responder rápidamente a los cambios antropogénicos en el suelo y la vegetación; 3) son bastante fáciles de muestrear (y muchas personas en general no se oponen a tomar muestras de artrópodos en comparación con los vertebrados); 4) representan el espectro de los niveles tróficos y 5) pueden ser importantes componentes sociológicos, agronómicos y económicos de los hábitats que altera el ser humano (McIntyre, 2000). Entre los artrópodos que han podido adaptarse a los ambientes urbanos se encuentran las arañas, ya que algunos aspectos de su biología pueden proveerles de algunas ventajas y desventajas para realizar la transición de un ambiente natural hacia uno urbano (Durán-Barrón, 2004).
Una forma de estimar los efectos de la urbanización en la naturaleza es el estudio de la estructura y función de los sistemas ecológicos a lo largo de zonas urbanas y rurales degradadas. Los efectos de la urbanización en la abundancia y la diversidad de artrópodos han sido poco estudiados, aunque la urbanización es considerada una de las principales causas de la disminución de las poblaciones de artrópodos. Por lo tanto, el estudio de los artrópodos en el entorno urbano es una estrategia oportuna para la investigación (Magura et al., 2008).
En pocos países del mundo se han realizado trabajos respecto a las arañas sinantrópicas. Estos estudios están enfocados básicamente a 2 grupos: las arañas que se encuentran o habitan en el interior y exterior (jardín y patio) de las viviendas y/o edificaciones humanas, y las que se establecen en áreas urbanas (calles, jardines, lotes baldíos, camellones, etc.). Dentro del primer grupo de estudio, en el continente europeo se encuentran las investigaciones realizadas por Urák (2005), quien cita 2 especies de arañas asociadas con las viviendas de Rumania; además del de Kostanjšek y Celestina (2008), quienes listan 4 especies de arañas sinantrópicas para Eslovenia. En el continente americano se encuentran los trabajos realizados en los Estados Unidos por Cutler (1973), quien registra 14 especies de arañas en las viviendas de la ciudad de Minneapolis-St. Paul, Minnesota; el de Kaston (1983), quien cita 158 especies encontradas en edificaciones y casas de los Estados Unidos; mientras que Guarisco (1999) reconoce 74 especies para el estado de Kansas. En Canadá, Williams (1999) registra 37 especies de arañas; mientras que para Cuba, Armas (2003) cita 31 especies, ambos trabajos realizados en 1 sola vivienda. En México, los estudios sobre el tema los han realizado Jiménez (1998), quien registra para la ciudad de La Paz, Baja California Sur, 42 especies de arañas; Cupul-Magaña y Navarrete-Heredia (2008), quienes mencionan sólo 10 familias de arañas para viviendas de Puerto Vallarta, Jalisco y Durán-Barrón et al. (2009), quienes citan 63 especies para casas de la ciudad de México.
Tanto en la investigación de Kaston (1983) como en la de Williams (1999), se plantean diferentes grados de sinantropismo, pero no tienen un sustento matemático en el cual basarse para proponer esta clasificación. Jiménez (1998), por primera vez emplea 2 tipos de índice (infestación y densidad) con la finalidad de determinar la abundancia relativa de las especies registradas y recientemente Durán-Barrón et al. (2009) utilizan los índices modificados por Jiménez (1998) y Durán-Barrón (2004), y proponen un índice de permanencia para poder conocer la presencia de las especies a lo largo del año y así sustentar mejor los niveles de sinantropismo propuestos en su trabajo. Con excepción de los trabajos mencionados, en la mayoría de las investigaciones sólo se registran las especies de arañas encontradas en las viviendas y las afinidades que existen entre estas especies respecto a las partes de las casas; además, en ellos se carece de un método sistematizado para la recolecta de las arañas en el interior de las casas.
En cuanto a investigaciones realizadas sobre las arañas asociadas con áreas urbanas (segundo grupo); en Europa, en la ciudad de Debrecen (al este de Hungría), Magura et al. (2008) realizaron un trabajo sobre la diversidad de arañas, escarabajos e isópodos en un gradiente urbano-rural, alrededor de la reserva forestal Nagyerd, sin encontrar diferencias significativas ni en la riqueza general de escarabajos ni en la riqueza de especies de isópodos a lo largo del gradiente; sin embargo, la riqueza de especies de arañas fue significativamente mayor en el área urbana en comparación con las suburbanas y rurales. En Latinoamérica, Brasil es el país con más conocimiento de su aracnofauna sinantrópica, entre los trabajos elaborados destacan los realizados por Brescovit (2002) y Japyassú (2002). Brazil et al. (2005) citan 30 especies de arañas en 3 ciudades con diferente tiempo de urbanización, encontrando que el barrio más antiguo presenta la mayor abundancia relativa de las especies identificadas, en este trabajo la recolecta de arañas se llevó a cabo para los 2 grupos de estudio aquí mencionados. Melo et al. (2010) citan 170 especies y morfoespecies de arañas en el bosque atlántico brasileño, el cual se encuentra bajo presión urbana. En México únicamente se conoce el trabajo realizado por Hurtado-Félix et al. (2007), quienes recolectaron arañas en diferentes zonas urbanas de la ciudad de Hermosillo, Sonora, correspondiendo los organismos a 17 familias, las cuales sólo fueron determinados hasta esa categoría taxonómica. Shochat et al. (2004) evaluaron la diversidad de arañas en 6 hábitats diferentes (parques desérticos, restos urbanos, industrial, agrícola, jardines residenciales xerófilos y jardines residenciales mésicos) en Arizona, EUA, encontrando una mayor diversidad en los parques desérticos y una menor diversidad en los jardines residenciales mésicos.
Es evidente que muchas cuestiones sobre las arañas sinantrópicas no se han respondido aún; por consiguiente, los objetivos de este estudio son: implementar un método de recolecta sistematizado para evaluar y comparar la diversidad de arañas en el interior de las viviendas en diferentes grados de urbanización, indicar si existen diferencias entre las arañas de un primer nivel y un segundo nivel de las viviendas, y modificar y proponer la prueba de Olmstead-Tukey para determinar los 4 niveles de sinatropismo (N. S).
Materiales y métodosÁrea de estudio. El presente trabajo se realizó en el municipio de Toluca, el cual se ubica en el centro del Estado de México. En dicho municipio predomina un clima templado subhúmedo con lluvias en verano C (w2), con una temperatura promedio de 13.7° C. La temperatura máxima es de 30° C y la mínima de 1.7° C, las temperaturas medias mensuales más bajas se han registrado en enero, febrero y diciembre, mientras que las temperaturas máximas se presentan en mayo (Secretaria de Desarrollo Urbano del Estado de México, 2003). Con base en el mapa E2, zonificación del territorio (uso de suelo) del Plan Municipal de Desarrollo Urbano del 2003, se tomaron 3 ambientes: urbano, en proceso de urbanización y agropecuario. En cada uno de estos ambientes se eligieron 3 viviendas de 2 niveles; en el ambiente urbano se tomaron 3 viviendas más, las cuales tienen un jardín, lo que constituyó un total de 12 casas en donde se realizaron los muestreos.
Las viviendas del ambiente 1 (urbano con jardín) se ubican en: casa 1: calle Puebla 417, Col. Nueva Santa María de las Rosas, municipio de Toluca, Estado de México (19°16’16.76” N, 99°38’28.88” O; 2 666 m); casa 2: calle Manzanares # 1, Barrio de Santa María Zozoquipan, municipio de Toluca, Estado de México (19°16’39.02” N, 99°36’43.44” O; 2 631 m); casa 3: calle Nicolás Bravo Norte # 727, Col. Unión, municipio de Toluca, Estado de México (19°17’55.76” N, 99°39’31.60” O; 2 686 m). Las casas del ambiente 2 (urbano sin jardín), se localizan en: casa 4: calle José María Morelos # 14, Barrio de Santa María Zozoquipan, municipio de Toluca, Estado de México (19°16’ 56.68” N, 99°37’0.86” O; 2 642 m); casa 5: calle Musas # 223A, Col. Infonavit San Gabriel, municipio de Metepec, Estado de México (19°16’17.93” N, 99°37’4.99” O; 2 654 m); casa 6: calle Instituto Literario # 302, Col. Centro, municipio de Toluca, Estado de México (19°17’10.73” N, 99°39’21.80” O; 2 668 m). Los hogares del ambiente 3 (proceso de urbanización), se ubican en: casa 7: calle Cerrada José López Portillo s/n, San Mateo Otzacatipan, municipio de Toluca, Estado de México (19°20’8.47” N, 99°36’21.77” O; 2 603 m); casa 8: carretera al Zoológico s/n; Col. La Magdalena Ocotitlan, municipio de Metepec, Estado de México (19°14’12.50” N, 99°37’7.80” O; 2 629 m); casa 9: calle Otzolotepec #1, San Diego de los Padres Cuexcontitlan, Sección 5A, municipio de Toluca, Estado de México (19°21’50.47” N, 99°36’32.80” O; 2 538 m). Las viviendas del ambiente 4 (agropecuario) se localizan en: casa 10: calle Miguel Hidalgo # 3, el Cerrillo Piedras Blancas, municipio de Toluca, Estado de México (19°24’47.83” N, 99°40’53.59” O; 2 609 m); casa 11: Camino a San Lorenzo s/n, el Cerrillo Piedras Blancas, municipio de Toluca, Estado de México (19°24’2.15” N, 99°40’38.72” O; 2 605 m); casa 12: domicilio conocido, Mina México, municipio de Almoloya de Juárez, Estado de México (19°24’34.21” N, 99°43’35.47” O; 2 618 m). Es necesario mencionar que las casas 5, 8 y 12, que no corresponden al municipio de Toluca, se encuentran a unos 500 m del límite del mismo.
Recolecta de organismos. En cada uno de los domicilios, una persona recolectó arañas 1 vez por mes durante 1 año (septiembre del 2009 a agosto del 2010), con un esfuerzo de recolecta de 100 minutos (50 minutos en el primer nivel de la casa y 50 minutos en el segundo), buscando los organismos con ayuda de una lámpara de luz blanca, sobre el piso y las paredes, en las esquinas de los muros, debajo de muebles y detrás de los cuadros y accesorios colgados en la pared. La captura de los organismos se realizó con ayuda de frascos de plástico y pinzas entomológicas. Los muestreos se realizaron durante el día entre las 13 y las 16 hrs.
Trabajo de gabinete. Los organismos se sacrificaron en alcohol al 70% y se determinaron con ayuda de un microscopio estereoscópico Zeigen, en algunos casos se disecaron el epiginio de las hembras y el pedipalpo de los machos, aclarándolos con KOH al 10%, además se utilizaron claves y publicaciones específicas; preservándolos bajo la técnica descrita por Levi y Levi (1993).
Índices de diversidad. Los índices de diversidad alfa (índice de diversidad de Shannon [H’] e índice de dominancia Simpson [1-D]) y el análisis de diversidad, se obtuvieron con el programa Paleontological Statistics (PAST) versión 1.93. El valor de complementariedad, grado de disimilitud, entre los sitios, índice de diversidad beta, se obtuvo con la fórmula (CAB); la cual se calcula a través de la riqueza total para ambos sitios combinados, SAB= a + b - c, donde “a” es el número de especies del sitio A, “b” es el número de especies del sitio B y “c” es el número de especies en común entre los sitios A y B; el número de especies únicas en cualquiera de los 2 sitios: UAB= a + b - 2c. A partir de estos 2 valores, se calcula la complementariedad de los sitios A y B como: CAB= UAB /SAB. La complementariedad varía desde 0, cuando ambos sitios son idénticos en cuanto a composición de especies, hasta 1, cuando las especies de cada sitio son completamente distintas (Moreno, 2001).
Índices y nivel de sinantropismo (N. S.). Se obtuvieron los índices de densidad (I. D.), infestación (I. I.) y permanencia (I. P.) propuestos por Durán-Barrón et al. (2009) para otorgar el nivel de sinantropismo a cada una de las especies recolectadas (Cuadro 1).
Intervalos para otorgar el nivel de sinantropismo (N.S.) propuestos por Durán-Barrón et al. (2009)
| Nivel de sinantropismo | Intervalos I. D. | Intervalos I. I. | IntervalosI. P. |
|---|---|---|---|
| Accidentales | 0-0.9 | 0-1 | 1-10 |
| Ocasionales | 1-2.9 | 1.1-3.5 | 11-30 |
| Frecuentes | 3.0-9.9 | 3.6-9.9 | 31-50 |
| Comunes | 10- enadelante | 10- enadelante | 51- enadelante |
Índice de densidad (I. D.): número de arañas de las especies A, B o C / número de unidades domiciliarias habitadas por las especies A, B o C X 100
Índice de infestación (I. I.): número de unidades domiciliarias infestadas por las especies A, B o C / número de unidades domiciliarias examinadas X 100
Índice de permanencia (I. P.): número de meses donde aparecen las especies A, B o C / número de meses totales X 100
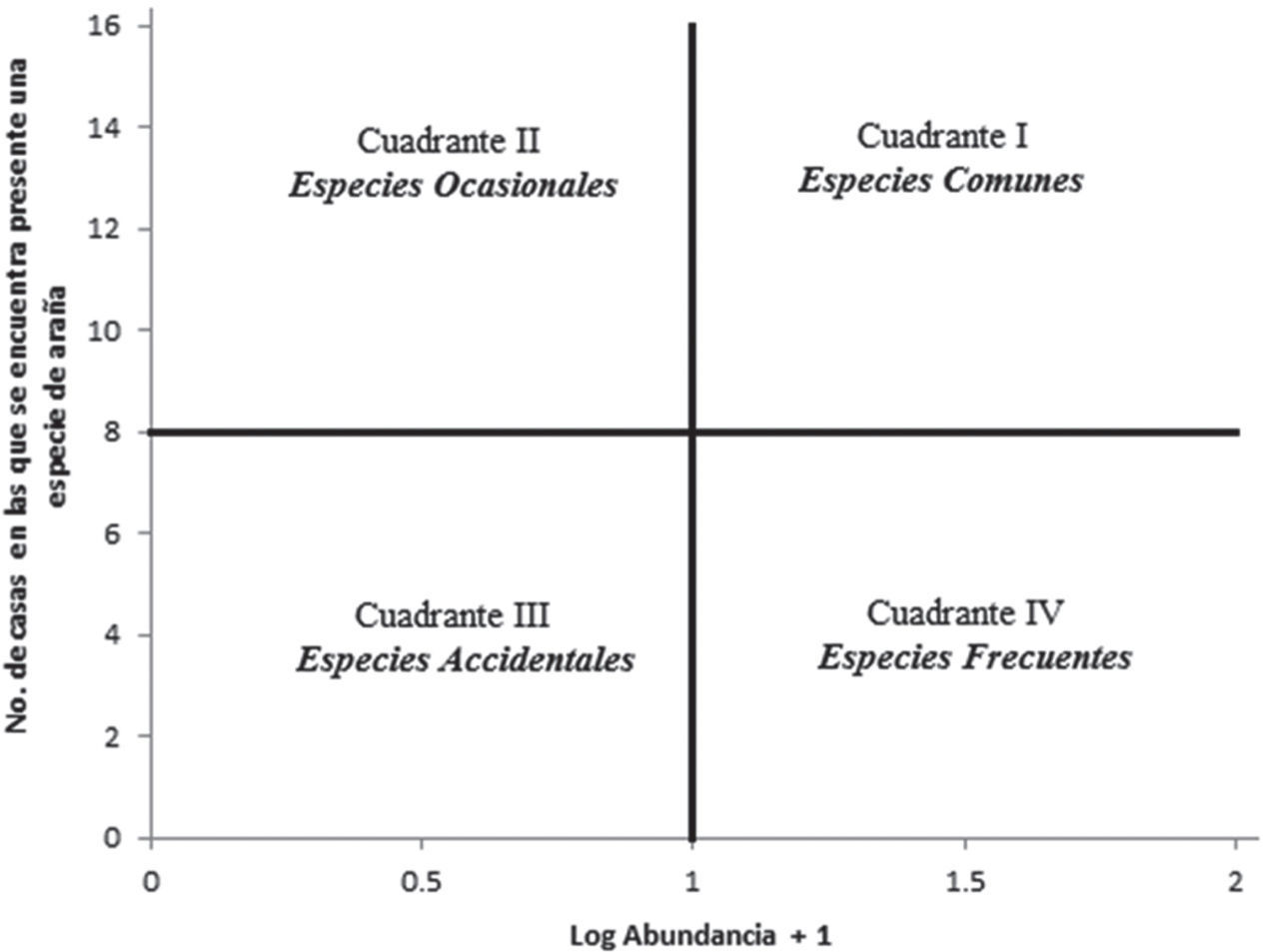
Se modificó el método de asociación no paramétrica de Olmstead-Tukey (Sokal et al., 1981) para otorgar un único nivel de sinantropismo a las arañas recolectadas en el interior de las viviendas. Una de las modificaciones de la prueba de Olmstead-Tukey consistió en ajustar las categorías que emplea la prueba antes mencionada, con los niveles de sinantropismo propuestos por Durán-Barrón et al. (2009), para emplear un único nombre en los niveles de sinantropismo para las especies de arañas (Fig. 1). Esta prueba permite categorizar a las especies arañas de forma gráfica en función de su abundancia y su prevalencia, ubicando a las especies en los siguientes cuadrantes: especies comunes (abundantes y frecuentes, ubicadas en el Cuadrante I), especies frecuentes (abundantes y poco frecuentes, ubicadas en el Cuadrante IV), especies ocasionales (poco abundantes y frecuentes, ubicadas en el Cuadrante II) y especies accidentales (poco abundantes y poco frecuentes, ubicadas en el Cuadrante III).
.")
Los valores a graficar son: en el eje de las abscisas, el logaritmo de la abundancia de cada una de las especies, mientras que en el eje de las ordenadas, el número de casas en que se encuentran presentes las especies. La segunda modificación a la prueba de Olmstead-Tukey se realizó al trazar los cuadrantes en la gráfica, en la cual para dividir el eje y formar los cuadrantes se toma el promedio de los datos graficados en cada eje; mientras que en el presente estudio, en cada uno de los ejes se tomó el valor más alto y se dividió entre 2, teniendo así una gráfica dividida en 4 cuadrantes iguales (Fig. 1).
ResultadosSe recolectaron 1 404 organismos (341 ♀♀,164 ♂♂ y 899 juveniles) en las 12 casas muestreadas. Los ejemplares recolectados correspondieron a 28 especies y 13 morfoespecies, todas ellas agrupadas en 16 familias (Cuadro 2). Las familias con mayor número de organismos fueron Pholcidae, Salticidae, Agelenidae, Filistatidae y Theridiidae, mientras que las familias con menor número fueron Zorocratidae, Thomisidae, Corinnidae, Oonopidae y Sparassidae (Cuadro 2). Las especies con mayor cantidad de organismos recolectados correspondieron a Physocyclus dugesi, Mexigonus sp. 1, Filistatinella sp. y Tegenaria sp.; mientras que el 41.5% del total de las especies recolectadas sólo estuvieron representadas por 1 organismo. Las 4 especies antes mencionadas se hallaron en más del 58% de las casas muestreadas, mientras que Mexigonus sp. 1 se encontró en 11 casas y Psilochorus sp. estuvo presente en las 12 casas muestreadas. Únicamente P. dugesi y Mexigonus sp. 1 se recolectaron todo el año (Cuadro 2).
Número de organismos, de casas y de meses en que se recolectaron las especies y morfoespecies. El número entre paréntesis indica la abundancia de cada familia
| Familia | Especie y/o morfoespecie | Abundancia | Núm. de casas | Núm. de meses |
|---|---|---|---|---|
| Agelenidae (119) | Tegenaria sp. | 24 | 7 | 9 |
| Morfo 1 | 2 | 1 | 2 | |
| Anyphaenidae (11) | Anyphaena sp. 1 | 2 | 1 | 1 |
| Anyphaena sp. 2 | 2 | 1 | 2 | |
| Anyphaena sp. 3 | 1 | 1 | 1 | |
| Anyphaena sp. 4 | 1 | 1 | 1 | |
| Anyphaena sp. 5 | 1 | 1 | 1 | |
| Araneidae (6) | Cyclosa sp. | 2 | 2 | 2 |
| Corinnidae (2) | Trachelas bulbosus | 2 | 1 | 1 |
| Filistatidae (119) | Filistatinella sp. | 64 | 9 | 12 |
| Kukulcania sp. | 1 | 1 | 1 | |
| Gnaphosidae (17) | Herpyllus sp. | 8 | 4 | 5 |
| Linyphiidae (18) | Morfo 1 | 1 | 1 | 1 |
| Morfo 2 | 1 | 1 | 1 | |
| Morfo 3 | 3 | 1 | 2 | |
| Morfo 4 | 1 | 1 | 1 | |
| Morfo 5 | 2 | 1 | 1 | |
| Lycosidae (37) | Arctosa sp. | 1 | 1 | 1 |
| Morfo 1 | 1 | 1 | 1 | |
| Morfo 2 | 1 | 1 | 1 | |
| Pardosa sp. 1 | 2 | 1 | 1 | |
| Pardosa sp. 2 | 1 | 1 | 1 | |
| Oonopidae (1) | Oonops sp. | 1 | 1 | 1 |
| Pholcidae (738) | Physocyclus dugesi | 175 | 7 | 12 |
| Physocyclus globosus | 1 | 1 | 1 | |
| Psilochorus sp. | 51 | 12 | 11 | |
| Salticidae (218) | Mexigonus sp. 1 | 95 | 11 | 12 |
| Mexigonus sp. 2 | 7 | 1 | 4 | |
| Scytodidae (10) | Scytodes longipes | 7 | 2 | 6 |
| Sparassidae (1) | Heteropoda sp. | 1 | 1 | 1 |
| Theridiidae (101) | Achaearanea sp. | 6 | 4 | 4 |
| Latrodectus mactans | 1 | 1 | 1 | |
| Morfo 1 | 2 | 2 | 2 | |
| Morfo 2 | 2 | 1 | 3 | |
| Morfo 3 | 1 | 1 | 1 | |
| Morfo 4 | 1 | 1 | 1 | |
| Morfo 5 | 3 | 2 | 3 | |
| Steatoda grossa | 11 | 4 | 7 | |
| Theridion sp. | 3 | 3 | 2 | |
| Thomisidae (2) | Misumenops sp. | 2 | 2 | 2 |
| Zorocratidae (3) | Zorocrates fuscus | 3 | 2 | 3 |
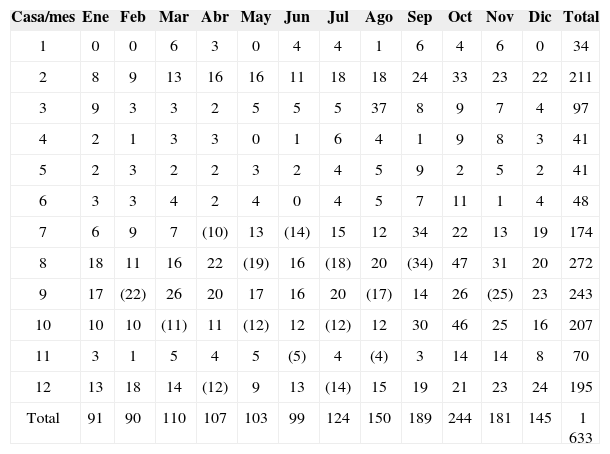
En la casa 1 no se encontraron organismos en 4 meses (diciembre-febrero y mayo), mientras que en las casas 4 y 6 no se hallaron organismos en mayo y junio, respectivamente; aun cuando se realizó la búsqueda de los organismos en el tiempo establecido. En algunos meses no se pudieron realizar las recolectas en las casas debido a circunstancias ajenas a la investigación, para obtener un valor en el mes de no recolecta, se obtuvo el promedio con los valores del mes anterior y el mes siguiente, al sumar los nuevos datos se obtuvo un aproximado de 1 633 organismos recolectados (Cuadro 3).
Número de organismos por mes, recolectados en cada casa muestreada. Los números entre paréntesis (meses donde no se pudo realizar la recolecta en la casa indicada) representan el promedio del mes anterior y del mes siguiente
| Casa/mes | Ene | Feb | Mar | Abr | May | Jun | Jul | Ago | Sep | Oct | Nov | Dic | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0 | 0 | 6 | 3 | 0 | 4 | 4 | 1 | 6 | 4 | 6 | 0 | 34 |
| 2 | 8 | 9 | 13 | 16 | 16 | 11 | 18 | 18 | 24 | 33 | 23 | 22 | 211 |
| 3 | 9 | 3 | 3 | 2 | 5 | 5 | 5 | 37 | 8 | 9 | 7 | 4 | 97 |
| 4 | 2 | 1 | 3 | 3 | 0 | 1 | 6 | 4 | 1 | 9 | 8 | 3 | 41 |
| 5 | 2 | 3 | 2 | 2 | 3 | 2 | 4 | 5 | 9 | 2 | 5 | 2 | 41 |
| 6 | 3 | 3 | 4 | 2 | 4 | 0 | 4 | 5 | 7 | 11 | 1 | 4 | 48 |
| 7 | 6 | 9 | 7 | (10) | 13 | (14) | 15 | 12 | 34 | 22 | 13 | 19 | 174 |
| 8 | 18 | 11 | 16 | 22 | (19) | 16 | (18) | 20 | (34) | 47 | 31 | 20 | 272 |
| 9 | 17 | (22) | 26 | 20 | 17 | 16 | 20 | (17) | 14 | 26 | (25) | 23 | 243 |
| 10 | 10 | 10 | (11) | 11 | (12) | 12 | (12) | 12 | 30 | 46 | 25 | 16 | 207 |
| 11 | 3 | 1 | 5 | 4 | 5 | (5) | 4 | (4) | 3 | 14 | 14 | 8 | 70 |
| 12 | 13 | 18 | 14 | (12) | 9 | 13 | (14) | 15 | 19 | 21 | 23 | 24 | 195 |
| Total | 91 | 90 | 110 | 107 | 103 | 99 | 124 | 150 | 189 | 244 | 181 | 145 | 1 633 |
Al agrupar las abundancias del cuadro 3 por estación climática (invierno
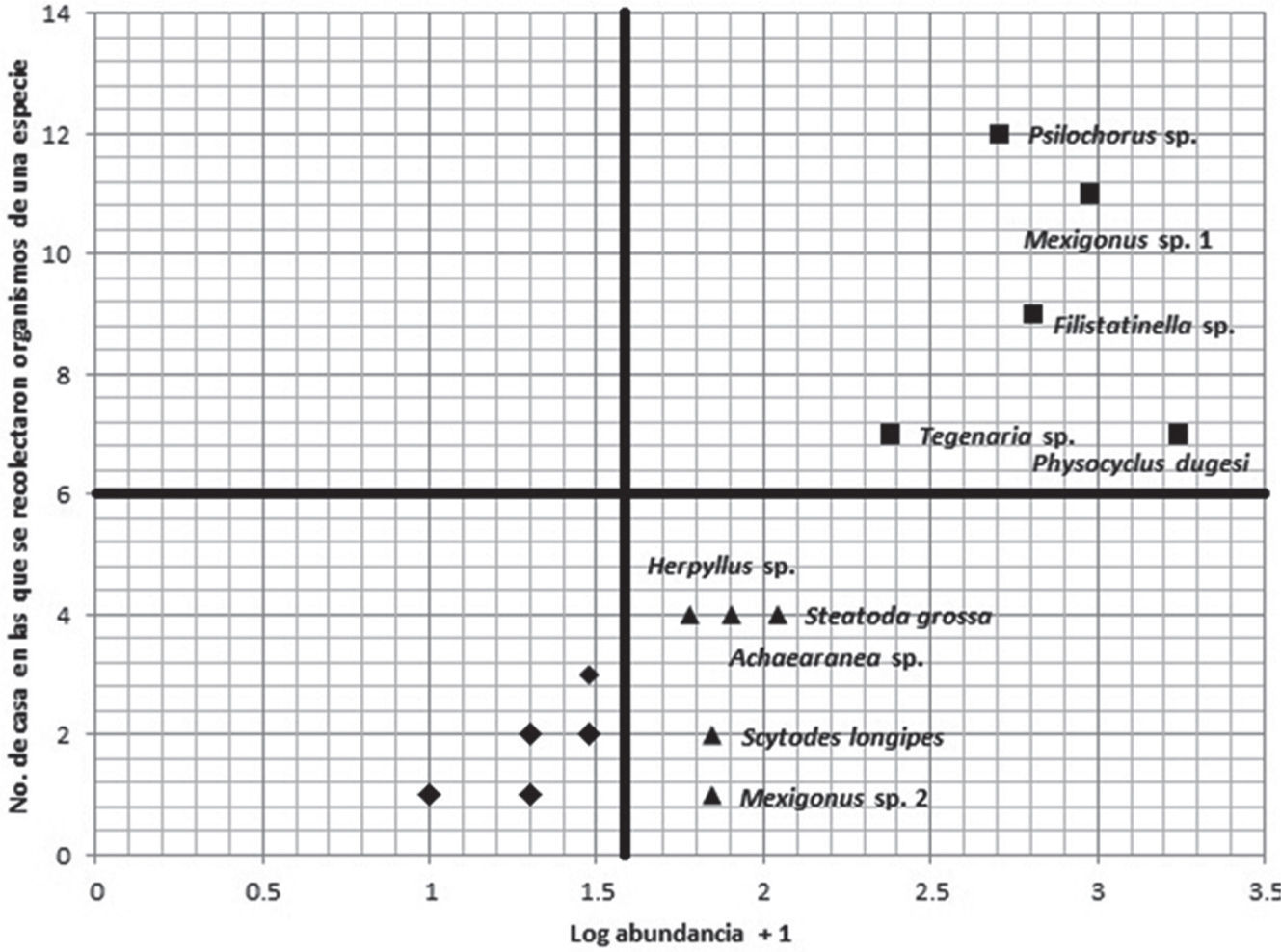
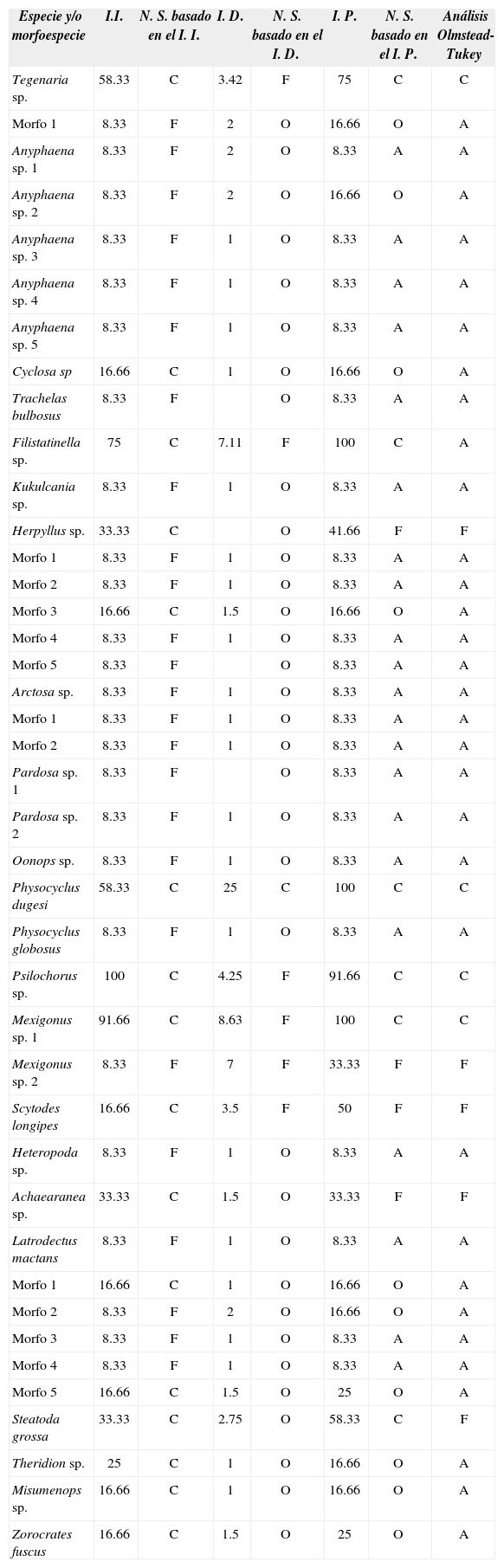
El I. D. muestra que P. dugesi es la única especie común dentro de todo el estudio, mientras que Tegenaria sp., Filistatinella sp., Psilochorus sp., Mexigonus sp.1, Mexigonus sp. 2 y Scytodes longipes son especies frecuentes en las casas muestreadas y el resto de las especies tiene un nivel de sinantropismo ocasional (Cuadro 4). El 53.6% de las especies difieren en su nivel de sinantropismo dado por los I. I., I. D. y el I. P., siendo P. dugesi la única especie que mantiene el mismo nivel de sinantropismo (común). El análisis de Olmstead-Tukey establece que Tegenaria sp., Filistatinella sp., P. dugesi, Psilochorus sp. y Mexigonus sp.1 son comunes (Fig. 2), mientras que Herpyllus sp., Mexigonus sp. 2, S. longipes, Achaearanea sp. y Steatoda grossa son frecuentes y el resto son consideradas como accidentales; ninguna especie quedó en la categoría de ocasional (Cuadro 4).
Nivel de sinantropismo (N. S.) para cada especie. Índice de infestación (I. I.), índice de densidad (I. D.), índice de permanencia (I. P.) y Olmstead-Tukey
| Especie y/o morfoespecie | I.I. | N. S. basado en el I. I. | I. D. | N. S. basado en el I. D. | I. P. | N. S. basado en el I. P. | Análisis Olmstead-Tukey |
|---|---|---|---|---|---|---|---|
| Tegenaria sp. | 58.33 | C | 3.42 | F | 75 | C | C |
| Morfo 1 | 8.33 | F | 2 | O | 16.66 | O | A |
| Anyphaena sp. 1 | 8.33 | F | 2 | O | 8.33 | A | A |
| Anyphaena sp. 2 | 8.33 | F | 2 | O | 16.66 | O | A |
| Anyphaena sp. 3 | 8.33 | F | 1 | O | 8.33 | A | A |
| Anyphaena sp. 4 | 8.33 | F | 1 | O | 8.33 | A | A |
| Anyphaena sp. 5 | 8.33 | F | 1 | O | 8.33 | A | A |
| Cyclosa sp | 16.66 | C | 1 | O | 16.66 | O | A |
| Trachelas bulbosus | 8.33 | F | O | 8.33 | A | A | |
| Filistatinella sp. | 75 | C | 7.11 | F | 100 | C | A |
| Kukulcania sp. | 8.33 | F | 1 | O | 8.33 | A | A |
| Herpyllus sp. | 33.33 | C | O | 41.66 | F | F | |
| Morfo 1 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 2 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 3 | 16.66 | C | 1.5 | O | 16.66 | O | A |
| Morfo 4 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 5 | 8.33 | F | O | 8.33 | A | A | |
| Arctosa sp. | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 1 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 2 | 8.33 | F | 1 | O | 8.33 | A | A |
| Pardosa sp. 1 | 8.33 | F | O | 8.33 | A | A | |
| Pardosa sp. 2 | 8.33 | F | 1 | O | 8.33 | A | A |
| Oonops sp. | 8.33 | F | 1 | O | 8.33 | A | A |
| Physocyclus dugesi | 58.33 | C | 25 | C | 100 | C | C |
| Physocyclus globosus | 8.33 | F | 1 | O | 8.33 | A | A |
| Psilochorus sp. | 100 | C | 4.25 | F | 91.66 | C | C |
| Mexigonus sp. 1 | 91.66 | C | 8.63 | F | 100 | C | C |
| Mexigonus sp. 2 | 8.33 | F | 7 | F | 33.33 | F | F |
| Scytodes longipes | 16.66 | C | 3.5 | F | 50 | F | F |
| Heteropoda sp. | 8.33 | F | 1 | O | 8.33 | A | A |
| Achaearanea sp. | 33.33 | C | 1.5 | O | 33.33 | F | F |
| Latrodectus mactans | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 1 | 16.66 | C | 1 | O | 16.66 | O | A |
| Morfo 2 | 8.33 | F | 2 | O | 16.66 | O | A |
| Morfo 3 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 4 | 8.33 | F | 1 | O | 8.33 | A | A |
| Morfo 5 | 16.66 | C | 1.5 | O | 25 | O | A |
| Steatoda grossa | 33.33 | C | 2.75 | O | 58.33 | C | F |
| Theridion sp. | 25 | C | 1 | O | 16.66 | O | A |
| Misumenops sp. | 16.66 | C | 1 | O | 16.66 | O | A |
| Zorocrates fuscus | 16.66 | C | 1.5 | O | 25 | O | A |
A= accidental, O= ocasional, F= frecuente, C= común.
.")
Análisis Olmstead-Tukey, en donde las especies comunes están representadas por los cuadros, las especies frecuentes están representadas por los triángulos y las especies accidentales están representadas por los rombos (32 especies; cuadro 4).
Las abundancias de cada uno de los ambientes (Cuadro 5) se analizaron por estación climática, encontrándose para el ambiente urbano con jardín una estacionalidad muy marcada del verano (121 organismos); respecto al resto de las estaciones, la prueba de X2 muestra que existe diferencia significativa (X2= 41.04, p< 0.00001, g. l. 3). En cuanto al ambiente urbano sin jardín, de igual manera la prueba de X2 muestra que existe diferencia significativa (X2= 19.7846, p< 0.0002, g. l. 3), hallando mayor número de organismos en verano y otoño (45 organismos en las 2 estaciones climáticas). En los ambientes en proceso de urbanización y agropecuario hay estacionalidad marcada del otoño (226 y 191 organismos respectivamente para la estación climática mencionada) respecto a las otras estaciones climáticas con diferencias significativas para el ambiente en proceso de urbanización (X2= 30.6806, p< 0.00001, g. l. 3) y para el ambiente agropecuario (X2= 64.9830, p< 0.00001, g. l. 3). Se analizó la abundancia de arañas encontradas en las casas que integran cada ambiente muestreado (cuadro 3), la prueba de Kruskal-Wallis indicó que existen diferencias significativas entre las abundancias de las arañas recolectadas en las casas que integran el ambiente urbano con jardín (p< 0.00002), ambiente en proceso de urbanización (p< 0.01) y ambiente agropecuario (p< 0.0003), mientras que para las casas que integran el ambiente urbano sin jardín no se encontraron diferencias significativas (p> 0.66) entre las abundancias.
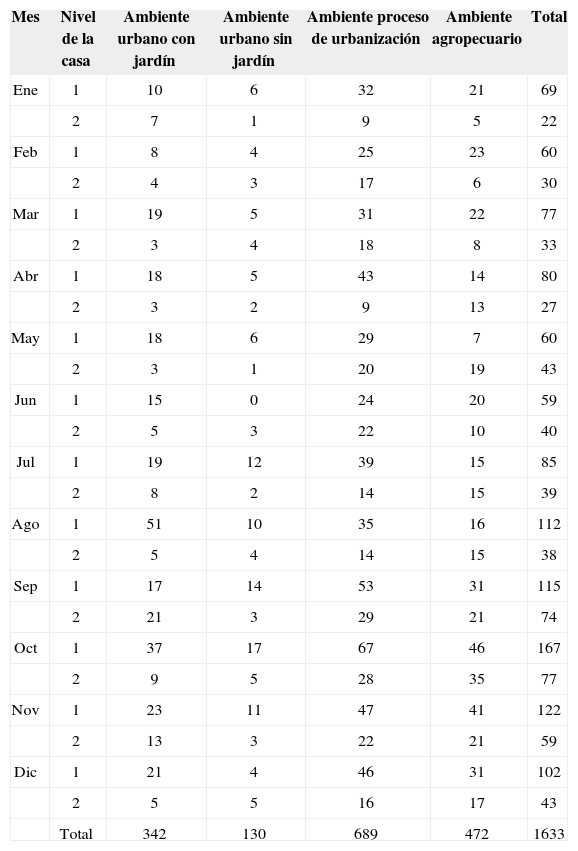
Número de organismos recolectados en cada uno de los niveles de las casas de los ambientes muestreados durante un año
| Mes | Nivel de la casa | Ambiente urbano con jardín | Ambiente urbano sin jardín | Ambiente proceso de urbanización | Ambiente agropecuario | Total |
|---|---|---|---|---|---|---|
| Ene | 1 | 10 | 6 | 32 | 21 | 69 |
| 2 | 7 | 1 | 9 | 5 | 22 | |
| Feb | 1 | 8 | 4 | 25 | 23 | 60 |
| 2 | 4 | 3 | 17 | 6 | 30 | |
| Mar | 1 | 19 | 5 | 31 | 22 | 77 |
| 2 | 3 | 4 | 18 | 8 | 33 | |
| Abr | 1 | 18 | 5 | 43 | 14 | 80 |
| 2 | 3 | 2 | 9 | 13 | 27 | |
| May | 1 | 18 | 6 | 29 | 7 | 60 |
| 2 | 3 | 1 | 20 | 19 | 43 | |
| Jun | 1 | 15 | 0 | 24 | 20 | 59 |
| 2 | 5 | 3 | 22 | 10 | 40 | |
| Jul | 1 | 19 | 12 | 39 | 15 | 85 |
| 2 | 8 | 2 | 14 | 15 | 39 | |
| Ago | 1 | 51 | 10 | 35 | 16 | 112 |
| 2 | 5 | 4 | 14 | 15 | 38 | |
| Sep | 1 | 17 | 14 | 53 | 31 | 115 |
| 2 | 21 | 3 | 29 | 21 | 74 | |
| Oct | 1 | 37 | 17 | 67 | 46 | 167 |
| 2 | 9 | 5 | 28 | 35 | 77 | |
| Nov | 1 | 23 | 11 | 47 | 41 | 122 |
| 2 | 13 | 3 | 22 | 21 | 59 | |
| Dic | 1 | 21 | 4 | 46 | 31 | 102 |
| 2 | 5 | 5 | 16 | 17 | 43 | |
| Total | 342 | 130 | 689 | 472 | 1633 |
La prueba de U de Mann-Whitney mostró que existen diferencias significativas entre las abundancias de las arañas del primero y segundo nivel de las casas (Cuadro 5), en el ambiente urbano con jardín (p< 0.002), ambiente urbano sin jardín (p< 0.002) y ambiente en proceso de urbanización (p< 0.0005); sólo en el ambiente agropecuario no se encontraron diferencias significativas. Se comparó entre ambientes la abundancia total de arañas del primer nivel mediante la prueba de Kruskal-Wallis, que mostró que existen diferencias estadísticamente significativas (p< 0.000001); así mismo, se comparó entre ambientes la abundancia total de arañas del segundo nivel mediante la misma prueba y mostró diferencias significativas (p< 0.0000007) (Cuadro 5).
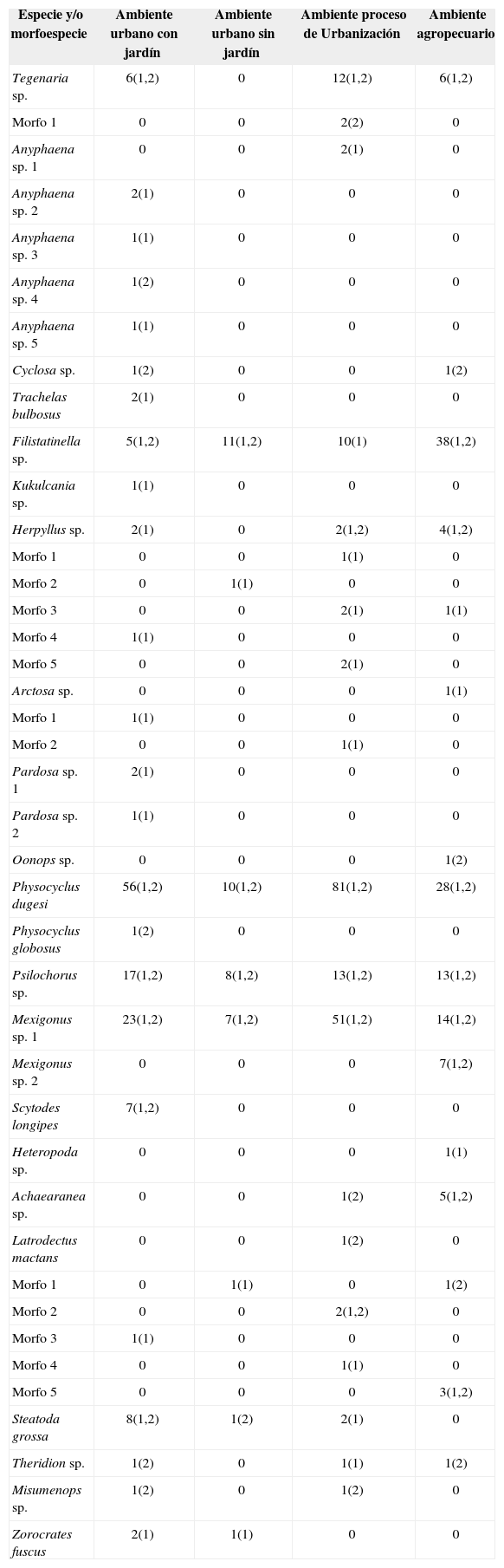
Se recolectaron diferentes especies de arañas en los distintos ambientes (Cuadro 6); sin embargo, algunas especies se hallaron en los 4, tal es el caso de Filistatinella sp., P. dugesi, Psilochorus sp. y Mexigonus sp. 1. Es importante indicar que, a excepción de que en el segundo nivel del ambiente en proceso de urbanización no se encontraron organismos de Filistatinella sp., sí se encontraron en los 2 niveles de las casas de los 4 ambientes. Asimismo, se encontraron especies de arañas en 3 ambientes, las cuales corresponden a Tegenaria sp. (excepto en el ambiente urbano sin jardín), recolectada en los 2 niveles; Herpyllus sp. fue hallada en los 2 niveles de los ambientes en proceso de urbanización y agropecuario, y en el primer nivel del ambiente urbano con jardín; S. grossa se encontró en el segundo y primer niveles de los ambientes urbano sin jardín y en proceso de urbanización, respectivamente, además de los 2 niveles del ambiente urbano con jardín. También se recolectaron especies en 2 ambientes, destacando Achaearanea sp. que tiene afinidad por ambientes en proceso de urbanización y agropecuario, en este último presente en los 2 niveles, mientras que en el ambiente en proceso de urbanización, estuvo presente en el nivel 2; Cyclosa sp. está presente en el segundo nivel de los ambientes, urbano con jardín y agropecuario; Misumenops sp. se recolectó en los ambientes urbano con jardín y en proceso de urbanización, pero sólo en el segundo nivel de ambos; por último, Zorocrates fuscus presenta afinidad por los ambientes urbanos (con jardín y sin jardín) únicamente en el primer nivel. El resto de las especies estuvieron presentes solamente en 1 ambiente, pero es necesario destacar que todas las especies de las familias Linyphiidae y Lycosidae se encontraron en el nivel 1, al igual que las especies de la familia Anyphaenidae, excluyendo aAnyphaena sp. 4, que se encontró en la parte alta (segundo nivel) de una casa.
Número de organismos por especie y/o morfoespecie recolectados en cada ambiente. El número entre paréntesis indica el nivel donde se encontraron los ejemplares (1= primer nivel, 2= segundo nivel)
| Especie y/o morfoespecie | Ambiente urbano con jardín | Ambiente urbano sin jardín | Ambiente proceso de Urbanización | Ambiente agropecuario |
|---|---|---|---|---|
| Tegenaria sp. | 6(1,2) | 0 | 12(1,2) | 6(1,2) |
| Morfo 1 | 0 | 0 | 2(2) | 0 |
| Anyphaena sp. 1 | 0 | 0 | 2(1) | 0 |
| Anyphaena sp. 2 | 2(1) | 0 | 0 | 0 |
| Anyphaena sp. 3 | 1(1) | 0 | 0 | 0 |
| Anyphaena sp. 4 | 1(2) | 0 | 0 | 0 |
| Anyphaena sp. 5 | 1(1) | 0 | 0 | 0 |
| Cyclosa sp. | 1(2) | 0 | 0 | 1(2) |
| Trachelas bulbosus | 2(1) | 0 | 0 | 0 |
| Filistatinella sp. | 5(1,2) | 11(1,2) | 10(1) | 38(1,2) |
| Kukulcania sp. | 1(1) | 0 | 0 | 0 |
| Herpyllus sp. | 2(1) | 0 | 2(1,2) | 4(1,2) |
| Morfo 1 | 0 | 0 | 1(1) | 0 |
| Morfo 2 | 0 | 1(1) | 0 | 0 |
| Morfo 3 | 0 | 0 | 2(1) | 1(1) |
| Morfo 4 | 1(1) | 0 | 0 | 0 |
| Morfo 5 | 0 | 0 | 2(1) | 0 |
| Arctosa sp. | 0 | 0 | 0 | 1(1) |
| Morfo 1 | 1(1) | 0 | 0 | 0 |
| Morfo 2 | 0 | 0 | 1(1) | 0 |
| Pardosa sp. 1 | 2(1) | 0 | 0 | 0 |
| Pardosa sp. 2 | 1(1) | 0 | 0 | 0 |
| Oonops sp. | 0 | 0 | 0 | 1(2) |
| Physocyclus dugesi | 56(1,2) | 10(1,2) | 81(1,2) | 28(1,2) |
| Physocyclus globosus | 1(2) | 0 | 0 | 0 |
| Psilochorus sp. | 17(1,2) | 8(1,2) | 13(1,2) | 13(1,2) |
| Mexigonus sp. 1 | 23(1,2) | 7(1,2) | 51(1,2) | 14(1,2) |
| Mexigonus sp. 2 | 0 | 0 | 0 | 7(1,2) |
| Scytodes longipes | 7(1,2) | 0 | 0 | 0 |
| Heteropoda sp. | 0 | 0 | 0 | 1(1) |
| Achaearanea sp. | 0 | 0 | 1(2) | 5(1,2) |
| Latrodectus mactans | 0 | 0 | 1(2) | 0 |
| Morfo 1 | 0 | 1(1) | 0 | 1(2) |
| Morfo 2 | 0 | 0 | 2(1,2) | 0 |
| Morfo 3 | 1(1) | 0 | 0 | 0 |
| Morfo 4 | 0 | 0 | 1(1) | 0 |
| Morfo 5 | 0 | 0 | 0 | 3(1,2) |
| Steatoda grossa | 8(1,2) | 1(2) | 2(1) | 0 |
| Theridion sp. | 1(2) | 0 | 1(1) | 1(2) |
| Misumenops sp. | 1(2) | 0 | 1(2) | 0 |
| Zorocrates fuscus | 2(1) | 1(1) | 0 | 0 |
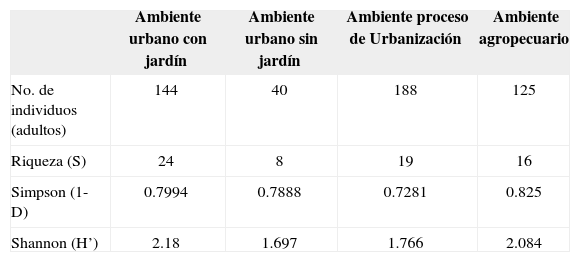
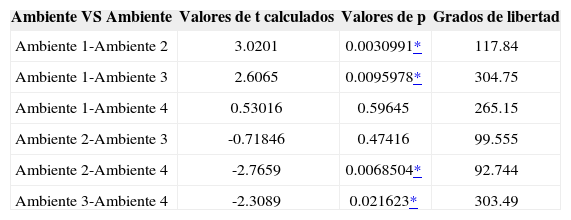
La riqueza de especies en cada uno de los ambientes fue diferente, encontrándose el valor más alto en el ambiente urbano con jardín (Cuadros 6, 7), el cual presentó el 58.5% de las especies recolectadas, contrastando drásticamente con el ambiente urbano sin jardín en donde se registró el 5.1% de las especies totales. Asimismo, el índice de diversidad mostró valores desiguales entre los ambientes, hallándose la máxima diversidad en el ambiente urbano con jardín y la diversidad mínima en el ambiente urbano sin jardín (Cuadro 7). Las especies dominantes en los ambientes no son las mismas y es por eso que los valores del índice de dominancia de Simpson muestran su valor más alto en el ambiente agropecuario y el menor en el ambiente en proceso de urbanización (Cuadro 7). La prueba estadística de t de Student aplicada a H’ en la comparación entre cada uno de los ambientes, reveló que solamente los ambientes urbano con jardín-agropecuario y urbano sin jardín-proceso de urbanización, son estadísticamente iguales (Cuadro 8).
Índices de diversidad alfa para cada uno de los ambientes muestreados
| Ambiente urbano con jardín | Ambiente urbano sin jardín | Ambiente proceso de Urbanización | Ambiente agropecuario | |
|---|---|---|---|---|
| No. de individuos (adultos) | 144 | 40 | 188 | 125 |
| Riqueza (S) | 24 | 8 | 19 | 16 |
| Simpson (1-D) | 0.7994 | 0.7888 | 0.7281 | 0.825 |
| Shannon (H’) | 2.18 | 1.697 | 1.766 | 2.084 |
Análisis de la diversidad (H’) de arañas entre cada uno de los ambientes mediante la prueba estadística de t
| Ambiente VS Ambiente | Valores de t calculados | Valores de p | Grados de libertad |
|---|---|---|---|
| Ambiente 1-Ambiente 2 | 3.0201 | 0.0030991* | 117.84 |
| Ambiente 1-Ambiente 3 | 2.6065 | 0.0095978* | 304.75 |
| Ambiente 1-Ambiente 4 | 0.53016 | 0.59645 | 265.15 |
| Ambiente 2-Ambiente 3 | -0.71846 | 0.47416 | 99.555 |
| Ambiente 2-Ambiente 4 | -2.7659 | 0.0068504* | 92.744 |
| Ambiente 3-Ambiente 4 | -2.3089 | 0.021623* | 303.49 |
Los valores de complementariedad muestran que entre cada par de ambientes analizados, la aracnofauna encontrada fue diferente, con valores superiores al 50% de diferencia en cuanto a la composición de especies de cada uno de los ambientes. El valor más alto de disimilitud estuvo en el ambiente par 3, 2 (proceso de urbanización y urbano sin jardín, respectivamente), en donde se tiene el 77% de especies distintas y sólo cuentan con 5 especies en común, mientras que el valor más bajo se presenta en el ambiente par 4, 3 (agropecuario y proceso de urbanización, respectivamente), con un 65% de especies distintas entre estos 2 ambientes, compartiendo sólo 9 especies (Cuadro 9).
Valores de complementariedad (disimilitud) y especies compartidas entre pares de ambientes muestreados. Los datos de la altura son el promedio de los metros a los que se encuentran las casas que integran cada uno de los ambientes. Ambiente 1= urbano con jardín, ambiente 2= urbano sin jardín, ambiente 3= proceso de urbanización, ambiente 4= agropecuario
| Complementariedad | ||||
|---|---|---|---|---|
| Ambientes | Ambiente 1 | Ambiente 2 | Ambiente 3 | Ambiente 4 |
| Ambiente 1 | - | 0.76 | 0.73 | 0.75 |
| Ambiente 2 | 6 | - | 0.77 | 0.73 |
| Ambiente 3 | 9 | 5 | - | 0.65 |
| Ambiente 4 | 8 | 5 | 9 | |
| Especies compartidas | ||||
Método de recolecta. El establecer un método de recolecta sistematizado permitió obtener un mayor número de organismos, comparado con el trabajo de Durán-Barrón (2004), quien sólo recolectó 322 arañas en un año. Además, se obtuvo un mayor número de especies de arañas en el interior de las viviendas, en comparación con los resultados de Cutler (1973), quien cita 14 especies y una riqueza ligeramente mayor con respecto a los estudios de Jiménez (1998) y Williams (1999), quienes citan 35 y 37 especies de arañas, respectivamente. McIntyre (2000) menciona que no existe una metodología estándar para los estudios de los artrópodos en los ambientes urbanos. Pues bien, en este trabajo se propone un método de recolecta sistematizado, el cual se espera sirva como base para futuros estudios, en donde se pretenda conocer la diversidad de arañas presentes en el interior de las casas. Además, partiendo de este método, es posible implementar las debidas modificaciones del mismo para responder otros tipos de preguntas biológicas sobre arañas sinantrópicas.
Niveles de sinantropismo. Para los índices de infestación u ocupación, densidad y permanencia, Durán-Barrón (2004) y Durán-Barrón et al. (2009) proponen y hasta cierto punto, dan sustento matemático a 4 niveles de sinantropismo (accidentales, ocasionales, frecuentes y comunes), pero los valores para separar cada uno de estos niveles fueron establecidos arbitrariamente, asignando un 25% de la muestra total a cada una de las categorías. Asímismo, para determinar el N. S. de una especie de araña, únicamente consideran los valores del índice de densidad; la pregunta es ¿para qué obtener los índices de infestación y permanencia, si sólo se otorga el nivel de sinantropismo con los valores del índice de densidad? Es cierto que cada índice proporciona información diferente, pero ésta debe ser conjunta para otorgar y darle mayor peso al N. S. que presenta cada especie; aunado a que en muchas ocasiones el N. S. dado para una especie, por cada uno de los índices, no siempre es el mismo, tal y como ocurrió en este trabajo y en algunas especies del trabajo de Durán-Barrón (2004) y Durán-Barrón et al. (2009). Por tal motivo, se propone la prueba de Olmstead-Tukey, con las modificaciones mencionadas en el método, como una herramienta para otorgar el N. S. a una especie, ya que esta prueba engloba valores utilizados en los índices de densidad e infestación (ocupación), dándole mayor soporte al otorgarle un nivel de sinantropismo a una especie, tal como se pudo observar en la presente investigación, en la que a 5 especies se les otorgó el N. S. de común, cuando con el I. D. sólo se tenía una especie dentro de ese nivel.
Estacionalidad por ambiente. La urbanización implica la modificación profunda de la temperatura (Hawkins et al., 2004); entonces, dependiendo del grado de urbanización de una área, la temperatura se modifica. Esto ayuda a entender por qué la estacionalidad de las arañas en algunos de los ambientes muestreados fue diferente. En los ambientes donde la urbanización no es tan elevada (agropecuario y en proceso de urbanización) la estacionalidad de las arañas se encontró en los meses más fríos (Cuadro 6). Ésto es porque los organismos necesitan resguardarse de las bajas temperaturas que se presentan en el exterior de los hogares, ya que en el interior se tiene una temperatura más favorable que les permite desarrollar sus actividades. Dicho incremento en la temperatura se da como resultado del uso de lámparas incandescentes (focos) y aparatos electrodomésticos. Si bien no se evaluó la temperatura en el interior de las casas, lo antes expuesto se deduce de lo expresado por Tischler (1973), quien menciona que en las zonas urbanas existe un incremento de calor por la concentración de las personas y el uso de las máquinas. El hecho de que en los ambientes más urbanizados la estacionalidad no se presente en los meses más fríos, sino en verano, puede deberse a que en estos ambientes existe una mayor reflectancia del calor y la capacidad de absorción del pavimento (aunque este último elemento, no está presente cerca de las viviendas de los ambientes agropecuario y en proceso de urbanización) (Kim, 1992), sin que la temperatura disminuya significativamente para las arañas, en estos ambientes. Además, en el interior de los hogares existe menor competencia por los recursos, ya que es común que en esta temporada se incremente el número de insectos en los jardines de las casas y la competencia por el alimento se incrementa. Con base en lo expresado con anterioridad, surgen las siguientes preguntas: ¿Cuál sería la estacionalidad en lugares urbanizados cercanos a las costas? ¿La relación urbanización-estacionalidad sería diferente? Temas sin duda, que es importante explorar.
Diversidad por ambiente. Gray (1989) acepta que la diversidad total debería disminuir en hábitats perturbados; sin embargo, en el presente estudio, el índice de Shannon reveló que la mayor diversidad se encontró en el ambiente urbano con jardín. Por consiguiente, el presente trabajo, al igual que el de Magura et al. (2008), no coincide con la hipótesis del disturbio intermedio, la cual indica que la diversidad es más alta en los niveles intermedios de perturbación (Connell, 1978). Es necesario destacar que los resultados muestran que la mayor abundancia de organismos y el segundo lugar en cuanto a riqueza de especies se presentó en el ambiente en proceso de urbanización, lo que indica que estos ambientes son de suma importancia porque podrían ser las zonas de amortiguamiento y transición en la adaptación de las especies de arañas entre diferentes grados de urbanización (Desales-Lara et al., 2011).
Es importante señalar que la riqueza y el índice de diversidad de Shannon fueron mayores en el ambiente urbano con jardín, respecto al ambiente urbano sin jardín, aunque ambos corresponden a un ambiente urbano. Esta diferencia la establece la existencia del jardín, ya que las plantas urbanas pueden soportar mayores densidades de fitófagos que las plantas rurales (McIntyre, 2000) y por consiguiente, existe mayor disponibilidad de alimento para las arañas, las cuales se pueden establecer en los jardines y de esta manera entrar fácilmente a las viviendas (Desales-Lara et al., 2011), además de que pueden adaptarse poco a poco a las condiciones de las mismas y habitar en ellas. Esta situación de los fitófagos también puede explicar el por qué en el ambiente agropecuario la riqueza de especies fue menor tomando en cuenta sólo 3 ambientes: urbano, proceso de urbanización y agropecuario.
Las especies de los ambientes pares fueron altamente distintas; esta diferencia en ambientes naturales o sin urbanización se atribuye a la variación altitudinal (Trevilla, 2008; Desales-Lara, 2009). Sin embargo, en áreas urbanizadas se puede atribuir a que en cada ambiente se presentan diferentes grados de perturbación y en cada uno de ellos interactúa un sistema complejo de factores abióticos, como temperatura, humedad, condiciones edáficas, acidez, contaminación y descomposición (McDonnell et al., 1997; Niemelä, 1999) y factores bióticos, que en la etapa de transición entre cada escala de urbanización no presentan una variación muy alta, lo que permite que algunas especies de arañas se encuentren en la escala de urbanización siguiente a la inicial. Eso también explica el por qué el valor más bajo de complementariedad se encontró en el ambiente par 4, 3 (agropecuario y en proceso de urbanización, respectivamente), siendo el ambiente agropecuario el primer nivel de perturbación y el siguiente, el ambiente en proceso de urbanización. Al cambiar drásticamente los factores antes mencionados, la diferencia en cuanto a la composición de especies de los ambientes comparados, será mayor, tal como lo indica la comparación de los ambientes en proceso de urbanización-urbano sin jardín y urbano sin jardín-urbano con jardín.
Nivel 1 de las casas versus nivel 2 de las casas. Los resultados mostraron que las arañas están en mayor abundancia en el primer nivel respecto al segundo nivel de las casas habitación estudiadas, aunque esta premisa no se cumple en el ambiente agropecuario. No se encontraron trabajos con los cuales comparar dicha situación, por lo que se proponen las siguientes hipótesis: a) el primer nivel de las casas presenta mayor cantidad de alimento para las arañas; b) el primer nivel de las viviendas ofrece mayor número de microhábitats con mejores condiciones (temperatura, humedad) para que los organismos puedan desarrollar sus actividades; c) el acceso (colonización) al primer nivel es mayor o más fácil que al segundo nivel, especialmente para las arañas activas. Asimismo, se propone que la “invasión” de las arañas generalmente se da de un primer nivel a un segundo nivel de las viviendas.
Se recolectaron especies de arañas que sólo se localizan en un nivel de la casa, lo cual obedece en gran parte a la biología de cada una de las familias encontradas, ya que éstas pueden ser clasificadas como activas y pasivas (Duran-Barrón, 2004), o cazadoras errantes y cazadoras sedentarias (Jiménez, 1998), respectivamente (aunque esta última clasificación ya no es muy aceptada). En este trabajo se encontraron arañas activas en el primer nivel (e.g. todas las especies de Lycosidae y 4 de 5 especies de Anyphaenidae) y arañas activas en el primero y segundo niveles (e.g. Agelenidae, Filistatidae, Gnaphosidae y Salticidae); estas últimas son las que presentan mayor abundancia, lo que puede atribuirse al hecho de que explotan al máximo los microhábitats ocultos que las viviendas les proporcionan (Jiménez, 1998). Tal vez lo mismo ocurriría con las arañas pasivas (P. dugesi y Psilochorus sp.) que se hallaron en el primer y segundo niveles de las viviendas, que también presentaron la mayor abundancia.
Para una araña, una vivienda es un gran ecosistema que le provee diferentes hábitats con las condiciones necesarias (alimento, temperatura, humedad) para desarrollar sus actividades, lograr su supervivencia y dejar descendencia. Dentro de una vivienda se ha establecido una “repartición” de la misma, de acuerdo con la biología de cada una de las familias como se mencionó en el párrafo anterior, pero al igual que en un ambiente natural, las especies de arañas de una misma comunidad explotan pequeñas diferencias ambientales para evadirse entre ellas (Norgaard, 1951) y para evitar la competencia tanto por alimento como por ocupar un mismo nicho ecológico (Foelix, 1996). En el interior de las viviendas ocurre lo mismo; ésto se ve reflejado al encontrar en el presente trabajo los organismos de Filistatinella sp. detrás de los cuadros o accesorios que se cuelgan en la pared, a Mexigonus sp. 1 y Mexigonus sp. 2 deambulando a media altura de los muros de las viviendas, a Psilochorus sp. en las esquinas de los muros, a P. dugesi debajo de los diferentes muebles de la casa y esquinas de los muros y a Tegenaria sp. corriendo sobre el piso.
Arañas de la ciudad de México vs. arañas de la ciudad de Toluca. La riqueza de arañas citada por Durán-Barrón (2004) en el interior de las casas de la ciudad de México (64 especies) es mayor a la encontrada en el presente trabajo (41 especies). Aun cuando entre las 2 ciudades existe una diferencia de especies del 93.9%, las diferencias se pueden atribuir a lo siguiente: a) los factores abióticos de cada ciudad son diferentes, la ciudad de México se encuentra a 2 200 m snm; su temperatura media anual varía entre 14 y 17° C., mientras que Toluca se encuentra a 2 680 m y su temperatura media anual varía entre 12° C y 13.7° C; b) la urbanización de ambas ciudades no se dio al mismo tiempo, la mancha urbana comenzó a extenderse a partir de 1950 en la ciudad de México, mientras que en la ciudad de Toluca la alta urbanización comenzó a principios de 1980. McIntyre (2000) menciona que la diversidad debería aumentar con la edad de urbanización de un área y al comparar la riqueza entre la ciudad de México y la ciudad de Toluca, la premisa anterior se cumple.
Durán-Barrón (2004) registra una estacionalidad en las arañas de las viviendas de la ciudad de México donde el mayor número de individuos se presenta en primavera y la disminución del mismo en otoño e invierno, contrastando drásticamente con la estacionalidad señalada en este trabajo, en el que la mayor abundancia se presentó en el otoño y la menor en la primavera. Sin embargo, como el esfuerzo de recolecta no fue uniforme durante el año en el estudio de Durán-Barrón (2004), los resultados podrían ser poco precisos. La estacionalidad que se describe en el trabajo de Durán-Barrón (2004) puede modificarse si se realizara un muestreo sistematizado. Sin embargo, si se mantiene la estacionalidad de las arañas en la primavera, la variación se puede atribuir a que en los meses más fríos las arañas tienden a entrar a las casas para resguardarse de las bajas temperaturas, además de que en el interior de los hogares puede existir mayor cantidad de alimento que el que se encuentra fuera de ellos (Desales-Lara et al., 2011).
A las familias que amablemente nos permitieron entrar a sus hogares a recolectar arañas, ya que sin su permiso no se hubiese podido realizar este trabajo. El primer autor agradece al Consejo Mexiquense de Ciencia y Tecnología (COMECYT) por la beca otorgada para realizar los estudios de maestría del cual se desprende este proyecto. A los revisores por sus comentarios y sugerencias para mejorar el escrito. A la memoria de Martha Desales Lara.