







En 2009 ocurrió un evento de mortandad masiva de Diadema mexicanum en La Entrega, bahías de Huatulco y mortalidad parcial en comunidades arrecifales cercanas. En este trabajo se estimó durante el periodo de mayo 2009 a octubre 2011 la densidad poblacional, mortalidad natural y distribución espacial de D. mexicanum en 2 localidades afectadas, San Agustín e isla Montosa. Además, se evaluó la relación existente entre la densidad poblacional y la temperatura, pH y precipitación pluvial. La densidad media (± DE) fue de 0.64±0.42 ind.m–2 en San Agustín y de 1.29±1.24 ind.m–2 en isla Montosa. Se calculó una tasa de mortalidad de 0.219 para San Agustín y 0.605 para isla Montosa. En ambas localidades la distribución fue agregada en todos los meses excepto en 5 de ellos, que fue aleatoria. En San Agustín la precipitación se relacionó significativamente con la densidad y en isla Montosa ninguna variable mostró una relación significativa. Isla Montosa presenta una mayor disponibilidad de recursos y sustrato más adecuado para la población de D. mexicanum que San Agustín, sin embargo, su población presentó mayor inestabilidad, probablemente debido a su cercanía con La Entrega, que ocasionó que su nivel de mortandad fuera mayor.
In 2009 there was a report on an event of mass mortality of Diadema mexicanum in La Entrega, Bahías de Huatulco, and partial mortality in adjacent reef communities. In this study we estimated population density of D. mexicanum, as well as natural mortality and spatial distribution in 2 affected localities (San Agustín and Isla Montosa) during a subsequent period (May 2009-October 2011). Additionally we evaluated the relation between population density and temperature, pH and pluvial precipitation. Mean density (± SD) was 0.64±0.42 ind.m–2 in San Agustín, and 1.29±1.24 ind.m–2 in Isla Montosa. Mortality rate was 0.219 in San Agustín, and 0.605 in Isla Montosa. For both localities spatial distribution was aggregated during the whole period except in 5 months when it was random. In San Agustín pluvial precipitation was significantly related to density, whereas in Isla Montosa there were not significant relations. Isla Montosa has a larger availability of resources and a more suitable substrate for D. mexicanum population than San Agustín, however, the sea urchin population in this locality showed more instability, probably because it is closer to La Entrega and the level of the die-off was higher.
Los erizos del género Diadema son erizos regulares que se consideran dentro de los equinodermos más ampliamente distribuidos, abundantes y ecológicamente importantes de aguas someras de los mares tropicales; encontrándose, principalmente, en hábitats de arrecifes someros, ya sea sobre arena, escombros o coral (Benítez-Villalobos, Domínguez y Gómez y López-Pérez, 2008; Muthiga y McClanahan, 2007). Los erizos del género Diadema son muy importantes, ya que sus densidades poblacionales altas y sus hábitos alimenticios son capaces de modificar directamente la distribución, abundancia relativa y composición de especies en los ecosistemas de algas y corales (Benítez-Villalobos et al., 2008; Carpenter, 1988,Carpenter, 1990Herrera-Escalante, López-Pérez y Leyte-Morales, 2005; Levitan, 1988a; Sammarco, Levington y Ogden, 1974).
Algunos autores mencionan a la especie Diadema mexicanum como un importante bioerosionador de los arrecifes y de gran relevancia en su restructuración y modelado (Alvarado y Chiriboga, 2008; Alvarado, Cortés y Reyes-Bonilla, 2012; Benítez-Villalobos et al., 2008; Herrera-Escalante et al., 2005). Se ha mencionado que una pérdida o disminución drástica de estos erizos conllevaría un desarrollo excesivo de las macroalgas que cubrirían y asfixiarían el coral, como ocurrió en el Caribe después del evento de mortandad masiva de D. antillarum a inicios de los años 80 (Lessios, 2005). Por otro lado, las altas densidades de Diadema provocan la bioerosión de los corales, por lo que es importante conocer cuál es el límite o la densidad que permita a los erizos contribuir al buen estado de los arrecifes (Alvarado y Chiriboga, 2008; Alvarado et al., 2012).
Los estudios realizados en bahías de Huatulco, Oaxaca, se han enfocado en su mayoría al sistema arrecifal de La Entrega, por ser uno de los más grandes y con mayor presencia de D. mexicanum, aunque algunos autores han incluido en su estudio a comunidades arrecifales cercanas (Benítez-Villalobos et al., 2008; Herrera-Escalante et al., 2005; Zamorano y Leyte-Morales, 2005). No obstante, en dichos trabajos únicamente se registra la densidad poblacional de esta especie y/o la bioerosión que ocasiona. En general, existen pocos estudios enfocados en la ecología de la población de este erizo en el Pacífico tropical mexicano (Díaz-Martínez, Benítez-Villalobos y López-Serrano, en prensa; Espino-Barr, Cibrián y García-Boa, 1996). Benítez-Villalobos, Díaz-Martínez y Martínez-García (2009) registraron un evento de mortandad masiva de D. mexicanum en la localidad de La Entrega, la cual pasó de ser la localidad con la mayor abundancia de este erizo de mar, a no presentar ningún organismo. Los autores concluyeron que esta mortandad se debió a una enfermedad, la cual se extendió a otras localidades cercanas en las que se pudieron observar organismos enfermos que presentaban signos similares a los descritos por Bak, Carpay y De Ruyter Van Steveninck (1984) durante el evento de mortandad masiva de D. antillarum, ocurrido durante los años 80 en los arrecifes del Caribe. El evento de mortandad masiva de D. mexicanum sin duda tuvo un efecto significativo en la ecología de los erizos de las localidades afectadas, no obstante, aún no se ha publicado información acerca de los posibles cambios en la dinámica de las poblaciones afectadas por dicho evento.
El objetivo del presente estudio fue estimar la densidad, mortalidad natural y distribución espacial del erizo de mar D. mexicanum, además de evaluar la relación existente entre la abundancia mensual y los valores de diferentes variables oceanográficas en las localidades de San Agustín e isla Montosa, bahías de Huatulco, Oaxaca, México, de mayo de 2009 a octubre de 2011.
Material y métodosÁrea de estudioEl estudio se realizó en 2 comunidades coralinas de bahías de Huatulco, Oaxaca: San Agustín (15°41’09” N, 96°14’05” O) e isla Montosa (15°45’48” N, 96°04’56” O) (fig. 1). El ecosistema arrecifal de bahías de Huatulco es reconocido como uno de los sistemas coralinos más importantes en el Pacífico mexicano (Glynn y Leyte-Morales, 1997; Reyes-Bonilla y Leyte-Morales, 1998). Los arrecifes coralinos de este sistema se extienden en bahías independientes, por lo que todos están físicamente aislados entre sí y se desarrollan en sitios someros, adyacentes a la costa sin formar una laguna entre la cresta del arrecife y la tierra (Leyte-Morales, 2001).

Trabajo de campo. Para la recolecta de datos, se realizaron muestreos mensuales de mayo de 2009 a octubre de 2011, colocando 10 transectos de 10 m de largo cada uno, perpendiculares a la línea de costa y separados entre ellos por aproximadamente 10 m. Se aseguró que los transectos quedaran colocados desde la parte somera hasta la parte profunda del arrecife. En cada transecto se inspeccionó 1 m a cada lado del mismo utilizando una varilla de PVC como referencia, dividiendo dicho transecto en 10 cuadrantes adyacentes de 1×2 m. Finalmente, se contabilizaron y registraron todos los erizos encontrados en cada uno de los cuadrantes (CARICOMP, 2001).
Análisis estadístico, densidadLos datos se analizaron mediante un modelo lineal generalizado; estos modelos se utilizan en casos en los que las varianzas no son constantes y los errores no presentan una distribución normal. Los datos de un conteo en el que la variable de respuesta es un número entero y se encuentra una gran cantidad de 0 es uno de estos casos, ya que la varianza puede incrementar linealmente con la media (Crawley, 2007).
Distribución espacialPara estimar el patrón de distribución espacial de D. mexicanum se utilizó el índice de Morisita (1959). Para comprobar si la distribución es aleatoria, se utilizó el modelo de distribución de Poisson. Por otra parte, para probar si la distribución es o no agregada, se utilizó el modelo de distribución binomial negativa (LUdwig y Reynolds, 1988).
MortalidadPara estimar la tasa de mortalidad natural (M) de los organismos, se utilizó el modelo de Berry modificado (Díaz-Martínez et al., en prensa; Ramos-Cruz, Sánchez-Meraz, Carrasco-Ayuso y Cervantes-Hernández, 2006). El algoritmo matemático para el cálculo de la mortalidad con este modelo es el siguiente:

donde: a= ordenada al origen, b= pendiente de la regresión o valor de la tasa de mortalidad total (Z parcial), t= intervalo de tiempo en el cual existe una disminución en la abundancia.
Variables oceanográficas. Para cuantificar la relación entre las variables oceanográficas de pH, temperatura superficial del mar (TS), precipitación pluvial mensual (PT) y la variación mensual de la densidad poblacional, se utilizó el modelo de regresión lineal múltiple paso a paso, representado a continuación

donde: Nt= densidad mensual, α= ordenada al origen en el modelo de regresión, bi= coeficientes que representan la contribución de cada una de las variables independientes en la predicción de la variable dependiente.
ResultadosDensidad. En San Agustín la densidad media (± DE) estimada fue de 0.64±0.42 ind.m–2, y se mantuvo relativamente constante durante el periodo de mayo 2009 a octubre 2011, con fluctuaciones entre 0.23 y 1.79 ind.m–2 como se puede observar en la figura 2. Únicamente se muestra un aumento en la densidad durante el mes de agosto de 2010.

Por otra parte, en isla Montosa la densidad media (± DE) fue de 1.29±1.24 ind.m–2, aproximadamente el doble que en San Agustín, con fluctuaciones mucho mayores que en el caso anterior, entre 0.14 y 5.08 ind.m–2. Se pueden apreciar 2 periodos (abril-mayo de 2010 y julio-septiembre de 2010), en los cuales la densidad se incrementa de manera importante y, a partir de septiembre 2010 comienza a disminuir hasta alcanzar valores similares a los del inicio del muestreo (fig. 3).

El modelo lineal generalizado mostró que hay diferencia significativa tanto entre las densidades encontradas en ambas localidades como en las encontradas en los diferentes meses comprendidos entre mayo 2009 y octubre 2011, siendo mayo, agosto, septiembre y octubre de 2010 aquellos que presentan mayor diferencia (p< 0.001) (fig. 4).
 encontrados en ambas localidades durante el periodo de mayo de 2009 a octubre de 2011. Las cajas blancas representan la localidad de isla Montosa, mientras que las grises representan a San Agustín.")
En San Agustín la población tuvo una distribución espacial de tipo agregada en 17 de los 22 meses, mientras que en los 5 meses restantes se presentó una distribución aleatoria (septiembre y diciembre de 2010 y de julio a septiembre de 2011).
De la misma forma, para el caso de isla Montosa el análisis mostró que la población tuvo una distribución espacial de tipo agregada en 16 de los 21 meses. Al igual que en San Agustín, en los 5 meses restantes la distribución fue aleatoria. No obstante, los únicos meses en los cuales tanto San Agustín como isla Montosa coincidieron en este tipo de distribución fueron agosto y septiembre de 2011. Los demás meses con distribución aleatoria fueron julio de 2009, febrero y abril de 2011.
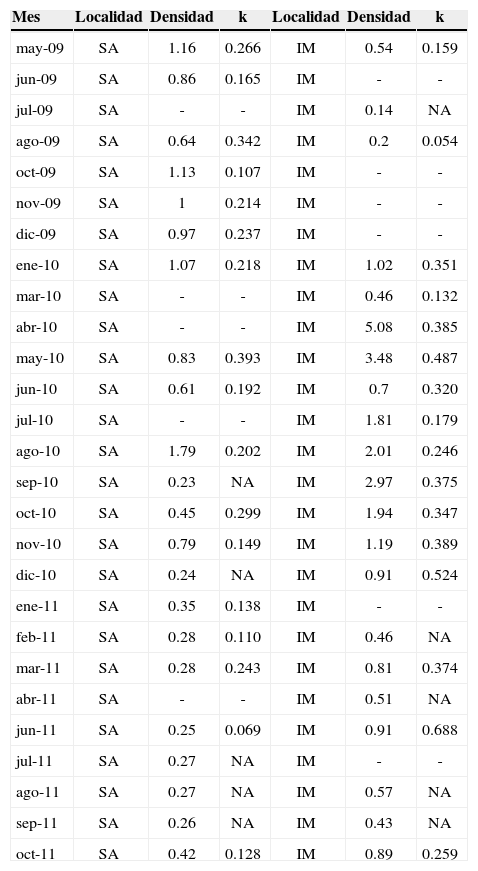
En términos del parámetro k, los meses que presentaron valores más altos en la localidad de San Agustín fueron mayo y agosto de 2009 (0.266 y 0.342, respectivamente), y mayo y octubre de 2010 (0.393 y 0.299, respectivamente). Estos valores indican que durante dichos meses, los organismos de la población se encontraron poco agregados. Por otra parte, los valores más bajos se presentaron en los meses de octubre de 2009 y junio de 2011, así como en el periodo de noviembre de 2010 a febrero de 2011, lo que indica mayor agregación en dichos meses (tabla 1).
Valores de agregación (k) y densidad promedio (ind.m–2) para cada uno de los meses comprendidos entre mayo de 2009 y octubre de 2011 en las localidades de San Agustín (SA) e isla Montosa (IM)
| Mes | Localidad | Densidad | k | Localidad | Densidad | k |
|---|---|---|---|---|---|---|
| may-09 | SA | 1.16 | 0.266 | IM | 0.54 | 0.159 |
| jun-09 | SA | 0.86 | 0.165 | IM | - | - |
| jul-09 | SA | - | - | IM | 0.14 | NA |
| ago-09 | SA | 0.64 | 0.342 | IM | 0.2 | 0.054 |
| oct-09 | SA | 1.13 | 0.107 | IM | - | - |
| nov-09 | SA | 1 | 0.214 | IM | - | - |
| dic-09 | SA | 0.97 | 0.237 | IM | - | - |
| ene-10 | SA | 1.07 | 0.218 | IM | 1.02 | 0.351 |
| mar-10 | SA | - | - | IM | 0.46 | 0.132 |
| abr-10 | SA | - | - | IM | 5.08 | 0.385 |
| may-10 | SA | 0.83 | 0.393 | IM | 3.48 | 0.487 |
| jun-10 | SA | 0.61 | 0.192 | IM | 0.7 | 0.320 |
| jul-10 | SA | - | - | IM | 1.81 | 0.179 |
| ago-10 | SA | 1.79 | 0.202 | IM | 2.01 | 0.246 |
| sep-10 | SA | 0.23 | NA | IM | 2.97 | 0.375 |
| oct-10 | SA | 0.45 | 0.299 | IM | 1.94 | 0.347 |
| nov-10 | SA | 0.79 | 0.149 | IM | 1.19 | 0.389 |
| dic-10 | SA | 0.24 | NA | IM | 0.91 | 0.524 |
| ene-11 | SA | 0.35 | 0.138 | IM | - | - |
| feb-11 | SA | 0.28 | 0.110 | IM | 0.46 | NA |
| mar-11 | SA | 0.28 | 0.243 | IM | 0.81 | 0.374 |
| abr-11 | SA | - | - | IM | 0.51 | NA |
| jun-11 | SA | 0.25 | 0.069 | IM | 0.91 | 0.688 |
| jul-11 | SA | 0.27 | NA | IM | - | - |
| ago-11 | SA | 0.27 | NA | IM | 0.57 | NA |
| sep-11 | SA | 0.26 | NA | IM | 0.43 | NA |
| oct-11 | SA | 0.42 | 0.128 | IM | 0.89 | 0.259 |
NA indica que no se encontró distribución agregada y los guiones indican que no se realizó muestreo durante ese mes en dicha localidad.
En la localidad de isla Montosa se presentaron valores más altos de k que en la localidad de San Agustín, lo que indica que la población de esta localidad se encuentra menos agregada que la población de San Agustín en la mayoría de los meses. Además, las fluctuaciones en dichos valores fueron mayores que para la localidad anterior: se pueden observar meses con valores muy altos del parámetro k, como mayo y diciembre de 2010 (0.487 y 0.524, respectivamente) y junio de 2011 (0.688), indicando muy poca agregación de los organismos. Sin embargo, el mes de agosto de 2009 muestra un valor muy bajo de k (0.054), mostrando un nivel alto de agregación durante ese mes (tabla 1).
MortalidadPara la localidad de San Agustín, a partir de 3 intervalos de disminución mensual continua en la abundancia de D. mexicanum (mayo-agosto 2009, enero-junio 2010, y enero-junio 2011), se estimó una tasa de mortalidad natural promedio de 0.219 (p< 0.05).
En isla Montosa se estimó una tasa de mortalidad natural promedio de 0.605 (p< 0.05), considerando 3 intervalos: abril-junio 2010, septiembre de 2010 a febrero de 2011, y junio-septiembre 2011.
Variables oceanográficasEn la localidad de San Agustín la temperatura superficial promedio del mar durante el periodo de muestreo fue de 28.1±2.04 °C y el valor medio de pH fue de 8.53±0.71 (fig. 5A). En el caso de isla Montosa, la temperatura superficial promedio fue de 27.83±2.2 °C y el valor promedio de pH fue de 8.59±0.74 (fig. 5B).
 y los valores de precipitación (rombos); pH (cuadrados) y temperatura (círculos); en San Agustín (A) e isla Montosa (B).")
La temporada de lluvias en la región de las bahías de Huatulco abarcó el periodo de mayo a diciembre en el 2009, y de mayo a octubre en el 2010 y 2011. La precipitación pluvial promedio del periodo de estudio fue de 85.091mm, con el valor máximo en agosto de 2010 (640.1mm) y valores de 0 en los meses de enero, marzo, abril, noviembre y diciembre de 2010 y el periodo de enero a abril de 2011 (fig. 5).
En el caso de San Agustín, el análisis de regresión lineal múltiple se resolvió en un paso, resultando en el siguiente modelo:

Este modelo mostró que la varianza total explicada sobre la densidad poblacional de D. mexicanum en términos de las variables oceanográficas fue de 41.7% (R2=0.417, R=0.646, p múltiple< 0.05), con un valor de correlación de 64.6%. La única variable que entró al modelo fue la precipitación, la cual mostró explicar un 28.4% de la varianza total. El resto de las variables en conjunto explicaron el 13.3% restante de la varianza y, por tanto, no entraron al modelo. En el caso de isla Montosa, el modelo mostró que ninguna variable explicaba la abundancia de D. mexicanum en dicha localidad.
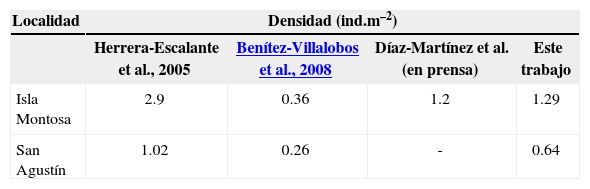
DiscusiónLos resultados muestran que existe un aumento considerable con respecto a la densidad poblacional registrada anteriormente para la localidad de San Agustín por Benítez-Villalobos et al. (2008), aunque aún no se alcanzan las densidades poblacionales documentadas por Herrera-Escalante et al. (2005) (tabla 2). A pesar de esto, la densidad encontrada en esta localidad es bastante menor a la encontrada en isla Montosa, lo cual coincide con trabajos anteriores, en los cuales San Agustín se ha registrado como la localidad con menor densidad (Benítez-Villalobos et al., 2008; Herrera-Escalante et al., 2005). En isla Montosa la densidad encontrada fue mayor a la documentada por Benítez-Villalobos et al. (2008), aunque nuevamente, no se alcanzan las densidades poblacionales documentadas por Herrera-Escalante et al. (2005) (tabla 2).
Comparación de las densidades poblacionales (ind.m–2) registradas anteriormente para San Agustín e isla Montosa y las documentadas en el presente trabajo
| Localidad | Densidad (ind.m–2) | |||
|---|---|---|---|---|
| Herrera-Escalante et al., 2005 | Benítez-Villalobos et al., 2008 | Díaz-Martínez et al.(en prensa) | Este trabajo | |
| Isla Montosa | 2.9 | 0.36 | 1.2 | 1.29 |
| San Agustín | 1.02 | 0.26 | - | 0.64 |
La cobertura coralina de San Agustín es mayor que la de isla Montosa (Leyte-Morales, 2001) y, sin embargo, sus densidades resultan menores. Esto no corresponde totalmente con lo propuesto por Zamorano y Leyte-Morales (2009), quienes establecen que existe una correlación positiva de la densidad de D. mexicanum con la cobertura coralina. Por otra parte, los resultados coinciden con otros trabajos en los cuales se asocia una alta densidad de D. mexicanum con una disminución de la cobertura por mortalidad del coral, debido a la preferencia de este erizo por arrecifes con gran cantidad de coral muerto, puesto que se alimentan de las algas que crecen sobre él (Alvarado et al., 2012; Eakin, 2001).
Otro factor que podría explicar las diferencias encontradas en la densidad, es la complejidad del hábitat. Clemente y Hernández (2008) señalan que al incrementar la complejidad estructural física de un hábitat aumenta significativamente la proporción de erizos de mar, tal como se observa en el presente estudio, donde el arrecife de San Agustín muestra un ambiente muy homogéneo, conformado casi en su totalidad por Pocillopora spp, mientras que el arrecife de isla Montosa presenta un ambiente más heterogéneo y complejo, al estar conformado por 7 especies de coral y gran cantidad de coral muerto y rocas (Leyte-Morales, 2001; López-López, 2011).
El evento de mortandad de D. mexicanum afectó a un menor número de organismos en estas comunidades que en la comunidad arrecifal de La Entrega; no obstante, se aprecia una disminución en la densidad de ambas localidades durante los primeros meses del estudio (figs. 2 y 3), correspondientes a los meses inmediatos al evento de mortandad de La Entrega, por lo que la disminución podría deberse a este suceso. Las fluctuaciones muy marcadas de densidad que se observan en los meses siguientes, principalmente en el caso de isla Montosa, posiblemente se deban a un periodo de inestabilidad de la población, el cual duró hasta los primeros meses del 2011, periodo a partir del cual las fluctuaciones fueron más atenuadas, indicando una posible estabilización de la densidad.
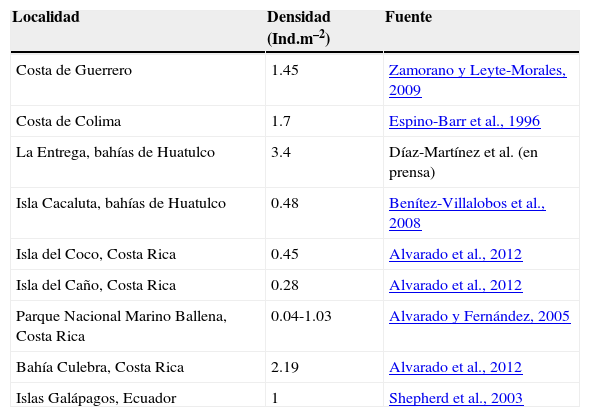
Comparando con los últimos datos registrados para otros sitios del Pacífico oriental tropical, la densidad de San Agustín resulta ser menor que las demás localidades, con excepción de isla Cacaluta, bahías de Huatulco; e isla del Caño e isla del Coco, Costa Rica. Por otra parte, isla Montosa presenta una densidad muy similar a la registrada para las costas de Guerrero y Colima, islas Galápagos y el Parque Nacional Marino Ballena de Costa Rica (tabla 3). Cabe destacar que La Entrega es el arrecife en el que se registraba mayor densidad, sin embargo, los últimos datos documentados (Díaz-Martínez et al., en prensa) fueron tomados antes del evento de mortandad que causó una drástica disminución de su población (Benítez-Villalobos et al., 2009).
Últimos datos reportados de densidad de D. mexicanum en otras localidades del Pacífico oriental tropical
| Localidad | Densidad (Ind.m–2) | Fuente |
|---|---|---|
| Costa de Guerrero | 1.45 | Zamorano y Leyte-Morales, 2009 |
| Costa de Colima | 1.7 | Espino-Barr et al., 1996 |
| La Entrega, bahías de Huatulco | 3.4 | Díaz-Martínez et al. (en prensa) |
| Isla Cacaluta, bahías de Huatulco | 0.48 | Benítez-Villalobos et al., 2008 |
| Isla del Coco, Costa Rica | 0.45 | Alvarado et al., 2012 |
| Isla del Caño, Costa Rica | 0.28 | Alvarado et al., 2012 |
| Parque Nacional Marino Ballena, Costa Rica | 0.04-1.03 | Alvarado y Fernández, 2005 |
| Bahía Culebra, Costa Rica | 2.19 | Alvarado et al., 2012 |
| Islas Galápagos, Ecuador | 1 | Shepherd et al., 2003 |
No existe información publicada acerca de la distribución espacial de D. mexicanum en estas localidades, sin embargo, se ha observado comportamiento de agregación tanto en esta especie como en otras del género Diadema (Levitan, 1988b; Maciá, Robinson y Nalevanko, 2007), por lo que se esperaría que la población presentara este tipo de distribución a lo largo de todo el periodo de estudio. El hecho de que esto no ocurra podría ser resultado de la poca cantidad de organismos observados durante ciertos meses, principalmente en la localidad de San Agustín. Además, la inestabilidad reflejada en las fluctuaciones del grado de agregación (k) podría ser también resultado del evento de mortandad que sufrió la región poco antes del periodo de muestreo (tabla 1).
Shepherd, Toral-Granda y Edgar (2003) mencionan que la distribución de D. mexicanum, al ser una especie altamente dependiente de grietas y escondites, posiblemente esté influenciada mayormente por las características del hábitat. Sin embargo, Alvarado (2008) menciona que en los diademátidos tropicales estas agregaciones representan actividades sociales y no parecen ser simples respuestas a limitantes ambientales. Este parece ser el caso en las localidades de estudio, puesto que el arrecife de isla Montosa presenta un ambiente mucho más heterogéneo y propicio que el de San Agustín y, sin embargo, no muestra una mayor agregación como habría de esperarse.
Asimismo, es poco probable que la distribución esté relacionada con la reproducción, ya que ciertos meses en los cuales se encontró una distribución aleatoria coinciden con la época reproductiva (mayo a septiembre) mencionada por Espino-Barr et al. (1996) y es justamente durante esta época cuando se esperaría que el erizo tuviera una distribución agregada, de acuerdo con otros autores que han mencionado este comportamiento en otras especies de erizos (D. antillarum: Bauer, 1976; Strongylocentrotus franciscanus: Levitan, Sewell y Chia, 1992). Además, se encontraron tanto valores muy altos como valores muy bajos de k durante el periodo reproductivo, lo cual no proporciona evidencia de diferencias entre la época reproductiva y el resto de los meses. Esto coincide con un estudio realizado por Levitan (1988b) con D. antillarum y con el realizado por McCarthy y Young (2002) con Lytechinus variegatus, en los cuales se menciona que no hay relación aparente entre el alistamiento reproductivo y la agregación.
Por otro lado, Pearse y Arch (1969) sugieren que las grandes agregaciones formadas por D. savignyi y D. setosum funcionan como unidades de protección social y se forman cuando las densidades de la población son tan altas que las grietas y escondites son limitados, no obstante, tampoco se aprecia una relación positiva entre la densidad y el grado de agregación en este estudio, ya que los valores de k son, en general, menores en San Agustín, indicando mayor agregación que en isla Montosa, a pesar de que esta última presenta una mayor densidad poblacional.
En lo que respecta a la mortalidad, se estimó una tasa de mortalidad natural para la población de isla Montosa muy alta y superior a la estimada para San Agustín. Una de las condiciones que podría causar esta diferencia es la tasa de sedimentación, ya que afecta a la comunidad béntica reduciendo el espacio disponible para el forrajeo y el movimiento, además de alterar la topografía del sustrato (Granja-Fernández y López-Pérez, 2008; Rogers, 1990), y se ha registrado que la tasa de sedimentación de ambas localidades es significativamente diferente, teniendo San Agustín una tasa de 133.30kg.m–2 al año, mientras que isla Montosa presenta una tasa mucho mayor (653.31kg.m–2 al año), debido a su relativa cercanía con el río Copalita (Granja-Fernández y López-Pérez, 2008).
La tasa de mortalidad estimada para San Agustín resulta ser similar a las estimadas para otras especies del mismo género: 0.36 para D. antillarum (Karlson y Levitan, 1990) y 0.29 para D. setosum (Ebert, 1982). Por otra parte, la tasa estimada para isla Montosa resulta mucho mayor, lo que evidencia nuevamente la inestabilidad a la que se encuentra sometida la población.
En cuanto a las variables oceanográficas, para la localidad de isla Montosa no se encontró relación de la densidad de D. mexicanum con ninguna variable. Dumas, Kulbicki, Chifflet, Fichez y Ferraris (2007) señalan que hay ocasiones en las que la abundancia de erizos en los arrecifes exhibe patrones espaciales complejos que no pueden ser relacionados con gradientes ambientales. Cuando esto sucede, es probable que muchos de estos factores estén ocurriendo como gradientes interrelacionados, lo cual vuelve difícil elucidar la distribución resultante de las especies. Esto podría explicar los resultados obtenidos en esta localidad.
Por otra parte, para la localidad de San Agustín únicamente la precipitación mostró una relación directa con la abundancia (R= 0.646). El aumento de la abundancia y la densidad de D. mexicanum durante la temporada de lluvias ha sido registrado anteriormente por Zamorano y Leyte-Morales (2009), quienes explicaron este incremento en la densidad como una respuesta al estrés que experimentan los erizos al verse sometidos a cambios drásticos de parámetros ambientales, debido al incremento de la temperatura y un decremento tanto de la salinidad como del contenido de oxígeno del agua y de la transparencia. Es posible que la población de San Agustín, al habitar en una zona poco dinámica, se vea más afectada por la combinación de estos factores y, por ello, aumente su abundancia como respuesta al estrés. La población de isla Montosa, en cambio, habita en una zona mucho más dinámica y, por esto, el posible efecto letal de sus altos niveles de sedimentación durante el periodo de lluvias se ve reducido por la influencia de la turbulencia del agua en la remoción del sedimento (Granja-Fernández y López-Pérez, 2008).
Diversos autores han mencionado a otros factores como controladores de la abundancia y densidad de erizos, tales como salinidad, turbidez, rugosidad del sustrato, fuerza del oleaje, contenido de oxígeno y fluctuaciones de mareas (Alves, Chícharo, Serrão y Abreu, 2001; Bauer, 1976; Clemente y Hernández, 2008; Espino-Barr et al., 1996; Muthiga y McClanahan, 2007; Zamorano y Leyte-Morales, 2009), por lo que sería necesario realizar estudios posteriores con dichos factores para comprobar si están relacionados o no con la abundancia de D. mexicanum en las localidades de San Agustín e isla Montosa.
AgradecimientosEste trabajo forma parte de la tesis de licenciatura de BRM y se realizó dentro del proyecto “Monitoreo de las poblaciones de Diadema mexicanum en las bahías de Huatulco y comunidades coralinas aledañas a Puerto Ángel” (CUP 21R1001), fue financiado por la Universidad del Mar.