




Los disturbios ocurridos en los arrecifes coralinos incrementan la competencia por espacio entre organismos. Nuestro estudio evaluó el efecto de los céspedes algales en el crecimiento y supervivencia del coral Pocillopora capitata al unir ramas muertas —colonizadas por algas— y vivas con ramas vivas del coral en 2 tipos de adhesión —lateral y apical—, en 2 temporadas —lluvias y secas—, en el Pacífico tropical mexicano. La información se obtuvo a través de la comparación de fotografías entre el inicio y final del experimento para cada temporada. Los resultados muestran que el crecimiento de las ramas de P. capitata no cambió de manera significativa, a pesar de la manipulación, las lesiones —corte y adhesión apical— y el contacto con los céspedes algales. Obtuvimos valores de supervivencia para las ramas de coral entre el 65 y el 100%. Así, en nuestros resultados y en los de otros trabajos se ha encontrado que el rápido crecimiento, la capacidad de resistir a la manipulación y al sobrecrecimiento de los céspedes algales, e incluso sobrecrecerlos con tejido coralino nuevo, además de que nuestra especie proviene de una familia útil en la recuperación de los arrecifes, sugieren que P. capitata puede ser utilizada en proyectos de restauración en las comunidades coralinas degradadas del Pacífico tropical mexicano.
The disturbances on coral reefs increase the competition for space between organisms. This study evaluated the effect of algal turfs on the growth and survival of the coral Pocillopora capitata. The experimental work was to unite dead branches —colonized by algae— and live with live coral branches in 2 types of adhesion —lateral and apical—, in 2 seasons —rainy and dry— in the Mexican tropical Pacific. The information was obtained through comparative pictures between the start and end of the experiment for each season. The results show that the coral branches growth did not change significantly despite the manipulation, the injuries —apical cut and adhesion— and by contact with algal turfs, survival values obtained for the branches of coral between 65 and 100%. Thus, the results presented here, and the results of other studies have found some qualities of P. capitata as rapid growth, ability and to manipulation resistance, also the turf algae overgrow resistance, and in addition their membership to a family used in the recovery of reefs make to this species useful in restoration projects, in degraded coral communities of the Mexican tropical Pacific.
Los disturbios naturales y antropogénicos a los que están expuestos los arrecifes de coral disminuyen los microhábitats y refugios para los diferentes habitantes de estos ecosistemas, lo que aumenta la posibilidad de que los diversos grupos de organismos compitan por los pocos espacios disponibles (Bell et al., 1991), afectando la composición de las especies en la comunidad. Por ejemplo, los disturbios disminuyen la cobertura de los corales escleractínios, ya sea por pérdida de tejido vivo y la mortalidad parcial o total de las colonias (Mumby et al., 2005).
Las respuestas de las interacciones entre corales y algas pueden ser variables, dependiendo de los mecanismos involucrados y de la morfología y la fisiología de ambos organismos (Jompa y McCook, 2002a; Titlyanov et al., 2007). Generalmente, se asume que las algas pueden colonizar a los corales si éstos se encuentran fisiológicamente debilitados por diversos disturbios (Jompa y McCook, 2002b; Titlyanov et al., 2007). Por ejemplo, si la proliferación de las algas debida a la ausencia de herbívoros y/o a cambios bruscos en el ambiente se mantienen por mucho tiempo, también es posible la muerte parcial de los corales con muy poca probabilidad de recuperación (Díaz-Pulido y McCook, 2002, 2004; Jompa y McCook, 2003). Sin embargo, las algas pueden ser capaces de estresar y colonizar a los corales (Quang-Young y Espinoza-Avalos, 2006). Las algas pueden afectar directa o indirectamente el crecimiento de los corales a través de diversos mecanismos como la abrasión, el sobrecrecimiento, el sombreado y la alelopatía (Jompa y McCook, 2003; Nugues y Bak, 2006; Titlyanov et al., 2007; Venera-Ponton et al., 2011), y es muy probable que los efectos sean especie-específicos (Nugues y Bak, 2006).
Otra condición que debilita a los corales es el aumento de lesiones; es decir, la remoción de tejido o esqueleto coralino, provocada por agentes físicos y/o biológicos como tormentas, incremento de la temperatura del agua, aumento de la turbidez, altas tasas de sedimentación, ramoneo de herbívoros y aumento de actividades humanas, por mencionar algunos ejemplos, lo que puede promover la colonización algal o producir la mortalidad parcial o total de las colonias (Box y Mumby, 2007; Fung et al., 2011). Por ejemplo, las lesiones artificiales infringidas en Montastraea annularis (taxón denominado actualmente como Orbicella annularis) fueron principalmente colonizadas por céspedes algales (Alvarado y Acosta, 2009).
En contraparte, los corales también pueden afectar el crecimiento de las algas (Jompa y McCook, 2002a; Nugues et al., 2004). El coral Porites cylindrica afecta el crecimiento del alga Lobophora variegata (Jompa y McCook, 2002a), mientras que O. annularis fue capaz de crecer sobre los céspedes algales (Vermeij et al., 2010).
En el Pacífico tropical mexicano la estructura general de las comunidades arrecifales es bastante homogénea y está constituida, principalmente, por colonias del género Pocillopora (Reyes-Bonilla, 1993, 2003). Dentro de los arrecifes del Pacífico tropical mexicano, los de la región de Ixtapa-Zihuatanejo están entre los mejor preservados (Leyte-Morales et al., 2006). En esta región también se han registrado alrededor de 150 especies de algas, incluidas aquellas formadoras de céspedes (Salcedo-Martínez et al., 1988; López, 2001).
Los corales del género Pocillopora son ramificados, resistentes y de rápido crecimiento. El crecimiento de las partes apicales de las ramas —con yemas de crecimiento— son estimuladas al ser consumidas o ramoneadas (Glynn y Wellington, 1983; Reyes-Bonilla, 2003; Reyes-Bonilla et al., 2005). Estos corales ramificados extienden su cobertura a lo largo de la fragmentación y una vez que los fragmentos se fijan, éstos crecen rápidamente al inicio del proceso (Jompa y McCook, 2002b). García-Ocampo (2005) estudió en esa misma región de Ixtapa-Zihuatanejo, la dinámica y estructura de tallas de reclutas, y determinó una distribución mayoritariamente aleatoria de las colonias, dominando las de talla pequeña, sugiriendo una baja sobrevivencia de las colonias grandes de las especies del género Pocillopora, como resultado de las múltiples actividades que se realizan en esta zona.
En la localidad de Las Gatas, Zihuatanejo, debido a su fácil acceso, se realizan múltiples actividades turísticas y de recreo, así como acciones de desprendimiento de las colonias de coral para obtener bivalvos (obs. pers.), por lo que es considerada como una de las playas con mayor impacto negativo (Zamorano y Leyte-Morales, 2009).
El objetivo de este estudio fue evaluar experimentalmente el efecto de los céspedes algales en la supervivencia y el crecimiento, en una de las especies de corales más abundantes en el Pacífico tropical mexicano, P. capitata, con base en el supuesto de que las colonias de corales sometidas a fuertes y diversos impactos serían más susceptibles a la colonización algal.
Materiales y métodosÁrea de estudio. El trabajo se llevó a cabo en playa Las Gatas (17°37'16.8” N, 101°33'08.55” O), localizada en la porción sur de la bahía de Zihuatanejo, Guerrero, México (Fig. 1). El clima en la región es de tipo cálido subhúmedo con lluvias en verano y se observan 2 temporadas climáticas bien marcadas: la de secas, de noviembre a abril y la de lluvias, de mayo a octubre (Salcedo-Martínez et al., 1988).

Playa Las Gatas alcanza 600m de amplitud con orientación E-O y está dividida por un enrocamiento (pretil) que encierra la porción sureste de la ensenada (parte interna) y la separa de la porción noroeste (parte externa). La comunidad de corales se encuentra aproximadamente a unos 150m al noreste del pretil, siendo P. capitata la especie de coral ramificado dominante. El área de muestreo está delimitada por el pretil en la porción más cercana a la costa y por el arenal en el otro extremo, con profundidades entre 3 y 5m.
Diseño experimental. El trabajo experimental para evaluar la supervivencia y el crecimiento de P. capitata al competir por espacio con los céspedes algales, se llevó a cabo con base en 4 tratamientos durante las temporadas de lluvias (de julio a noviembre de 2007) y secas (de noviembre de 2007 a junio de 2008). El crecimiento y la supervivencia fueron evaluados con fotografías obtenidas al inicio y al final del experimento para cada temporada.
El proceso consistió en fijar lateral y apicalmente fragmentos de coral vivo —sin algas— y muerto — cubiertos por céspedes algales— a ramas de coral sano, basado en 4 tratamientos (Fig. 2). En el tratamiento 1 (T1), fue adherido lateralmente un fragmento muerto colonizado por algas a una rama sana (Fig. 2B); en el tratamiento 2 (T2), fue unido de manera lateral un fragmento con tejido coralino vivo y sin algas a una rama sana; en el tratamiento 3 (T3), un fragmento muerto colonizado por algas fue adherido a la parte apical de una rama sana (Fig. 2A); y en el tratamiento 4 (T4), un fragmento con tejido coralino vivo fue unido a la parte apical de una rama sana (Figs. 2B, C). Para los tratamientos donde se unió apicalmente el fragmento, se hizo un corte en la punta de la rama de coral de la colonia. El número de unidades experimentales para cada tratamiento se presenta en el cuadro 1.
 Tratamiento T3; B) tratamiento T4 y T1; C) tratamiento T4, crecimiento de tejido coralino sobre la superficie de la manguera de plástico que une una rama viva a una rama de la colonia de coral con lesión apical.")
Los fragmentos de coral muerto y colonizados por algas fueron obtenidos del fondo, mientras que los fragmentos de coral vivos se extrajeron de otra colonia de la misma especie. Los fragmentos o ramas de corales vivos y muertos midieron entre 5 y 15cm de largo. No se requirió permiso de recolecta, debido a que no es una especie protegida; sin embargo, con el fin de no impactar la comunidad, se hicieron las réplicas mínimas necesarias para que fueran representativas.
Obtención de datos. En las fotografías obtenidas se utilizó una tabla de poliestireno reticulada para determinar la línea inicial del crecimiento del tejido coralino en las ramas de coral vivo y el crecimiento algal para el caso de los tratamientos con fragmentos muertos. La comparación de las fotografías —inicio y final del experimento— fue evaluada para lluvias (140 días) y secas (190 días). Las fotografías fueron analizadas con el software Sigma Scan Pro versión 5.0 para estimar el área de crecimiento del tejido coralino o el avance de las algas. Con las fotografías se calculó el total de la superficie de crecimiento coralino o algal en centímetros cuadrados y después se dividió entre el número de días que duró cada tratamiento para obtener el promedio del crecimiento diario. Posteriormente, se multiplicó por 30 días para obtener el crecimiento mensual y, así, estandarizar los datos para ambas temporadas en cm2 mes−1. Análisis estadísticos. Los porcentajes de supervivencia de cada temporada fueron analizados a través de una prueba de comparación de proporciones. Los datos de crecimiento se evaluaron con un análisis de varianza de una vía no paramétrico (Prueba de Kruskal-Wallis) y, posteriormente, con una prueba de comparación múltiple de rangos (Prueba de Dunn). Todos los análisis se realizaron en el software Statistica versión 10.
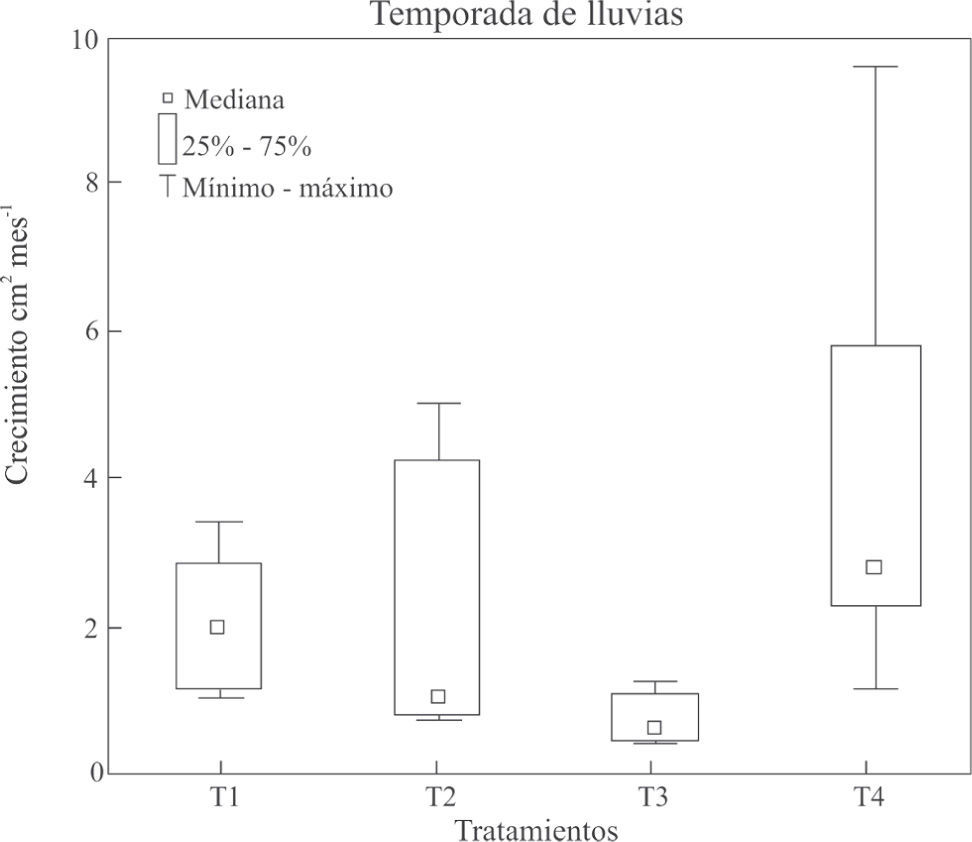
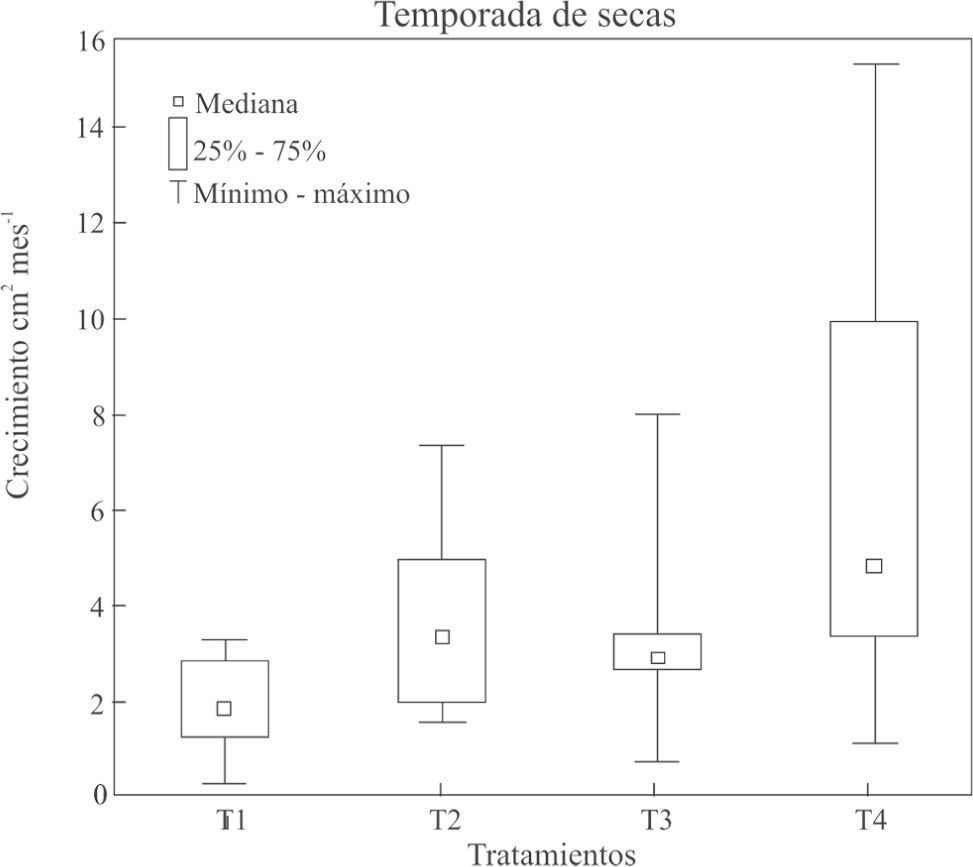
ResultadosSupervivencia y crecimiento del coral. La supervivencia de la temporada de lluvias (90.6%; n=32) fue mayor a la temporada de secas (73%; n=64; p=0.05) (Cuadro 1). Respecto al crecimiento de tejido coralino, en la temporada de lluvias hubo un efecto significativo de las condiciones experimentales —tipo de adhesión de fragmentos con y sin colonización algal— en el crecimiento del coral (p<0.05); es decir, las ramas del T4 —fragmento vivo colocado apicalmente— tuvieron un mayor crecimiento coralino que las del T3 —fragmento muerto con algas colocado apicalmente—, con un crecimiento de 4.06cm2 mes−1 vs. 0.78cm2 mes−1, respectivamente(p<0.05) (Figs. 2B). En la temporada de secas también se obtuvo una diferencia significativa en las condiciones experimentales entre el tipo de adhesión del fragmento con y sin colonización algal, en el crecimiento del coral, donde son significativamente distintos(p<0.05). Las ramas del tratamiento T1 — fragmento muerto adherido lateralmente— tuvieron un menor crecimiento comparado a las ramas del T4 — fragmento vivo adherido apicalmente—, con crecimientos de 1.92cm2 mes−1vs. 6.09cm2 mes−1, respectivamente(p<0.05) (Figs. 2C).
Efectos de los céspedes algales en las ramas de P. capitata para ambas temporadas (lluvias y secas)
| Temporadas | ||||||||
|---|---|---|---|---|---|---|---|---|
| Lluvias† | Secas‡ | |||||||
| Tipo de adhesión | Lateral | Apical | Lateral | Apical | ||||
| Tipo de fragmentos | muerto | vivo | muerto | vivo | muerto | vivo | muerto | vivo |
| Tratamiento | T1 | T2 | T3 | T4 | T1 | T2 | T3 | T4 |
| Número de unidades experimentales | 8 | 8 | 8 | 8 | 16 | 16 | 16 | 16 |
| Supervivencia % | 87.5 | 88 | 100 | 88 | 75 | 65 | 65 | 87.5 |
| Promedio crecimiento de coral (cm2 mes−1) | 2.01±0.13 | 2.02±0.26 | 0.78*±0.05 | 4.06*±0.36 | 1.92±0.08 | 3.63±0.19 | 3.32±0.26 | 6.09±0.53 |
| Promedio crecimiento de coral (cm mes−1) | 1.41 | 1.42 | 0.8 | 2.01 | 1.4 | 1.9 | 1.8 | 2.5 |
Crecimiento algal. En las 96 ramas de coral de los 4 tratamientos experimentales no hubo colonización algal sobre el tejido coralino. En 82 ramas se mantuvo la cobertura algal sin cambios aparentes —respecto al inicio de cada experimento— y en los 14 restantes hubo crecimiento de tejido coralino que incluso cubrió a las algas de los fragmentos muertos adheridos.
DiscusiónLa interacción entre los corales y las algas es variable y cmpleja; por un lado las algas, en su carácter de oportunistas, pueden sobrecrecer al coral sólo en condiciones de debilitamiento causado por factores de estrés externos (McCook, 2001), favoreciendo su proliferación (Díaz-Pulido y McCook, 2002; 2004; Jompa y McCook, 2003). En este estudio, a pesar de que sobre las ramas del coral P. capitata se introdujeron factores estresores —adhesión y corte, según fuera el tratamiento—, para provocar el debilitamiento del coral y promover la colonización algal, en ninguno de los tratamientos se observó colonización algal sobre las ramas de coral. Estos resultados sugieren que P. capitata es altamente resistente a la adhesión y fragmentación, como sucede con Acropora palmata que, a pesar de los continuos impactos antropogénicos y/o ambientales, mantiene su crecimiento relativamente constante (Jordan-Dahlgren, 1992). Estudios en corales masivos han mostrado resultados no concluyentes sobre la superior capacidad competitiva de los corales sobre las algas en ausencia de estresores o disturbios (McCook, 2001). Por ejemplo, se ha observado que M. annularis (taxón actualmente denominado como O. annularis), una importante especie constructora de arrecifes en el Caribe, fue capaz de desplazar a las algas con una tasa de crecimiento promedio de 0.12mm 3 semanas− 1, pero en el tratamiento con adición de nutrientes el efecto se revirtió y el césped algal sobrecreció a los corales en una tasa mayor de 0.34mm 3 semanas−1, mostrando que a altas concentraciones de nutrientes las algas son los competidores dominantes (Vermeij et al., 2010).
Estudios con manipulación experimental han mostrado que los resultados de la interacción competitiva entre corales y algas pueden ser variables y específicos entre las especies involucradas (Quan-Young y Espinoza-Avalos, 2006; Vermeij et al., 2010; Venera-Ponton et al., 2011). Por ejemplo, el coral Montastraea faveolata (taxón denominado actualmente como O. faveolata) fue completamente sobrecrecido por un césped mixto de algas en aproximadamente 6-9 meses después de que las algas fueron adheridas al esqueleto del coral, probablemente por el sombreo producido por el césped y quizá por la acumulación de sedimento que ocasiona una especie de enterramiento del coral (Quan-Young y Espinoza-Avalos, 2006). Otras especies como Montastraea franski (taxón denominado actualmente como Orbicella franski) y Mycetophyllia aliciae tuvieron alta capacidad competitiva, con bajo sobrecrecimiento por algas, no obstante, no se puede despreciar el hecho de que toda la información disponible sobre las interacciones coral-alga, es parcial y que no se puede extrapolar a largo plazo (Tanner, 1995; Nugues y Bak, 2006).
Por otra parte, los fragmentos con tejido de P. capitata que se fijaron con una técnica de mínima manipulación —adhesión de los fragmentos a ramas— tuvieron una supervivencia de hasta el 97%, mientras que en los que se usó una técnica más invasiva —corte apical y adhesión de fragmentos a ramas—, la supervivencia fue del 83% y con una tasa promedio del 80% para ambas temporadas. Además, estos corales tuvieron un crecimiento promedio de 2.21cm2 mes−1 (aproximadamente 1.4cm mes−1) en época de lluvias y 3.74cm2 mes−1 (aproximadamente 1.9cm mes−1) en secas. Nuestros resultados son menores a los registrados por Liñán-Cabello et al. (2011), donde la supervivencia de los fragmentos de Pocillopora spp. fue de 95.5% y la tasa de crecimiento coralino de aproximadamente 3.52cm mes−1 para una localidad de Colima con condiciones muy parecidas a las de nuestro sitio de estudio. Sin embargo, la tasa de crecimiento en P. capitata es mucho mayor a la documentada por Guzmán y Cortez (1989) para P. damicornis que fue de 21.3mm en la estación seca (aproximadamente 0.42cm mes−1) que fue la estación de mayor rendimiento en isla Caño en Costa Rica.
En las ramas de coral donde se adhieren fragmentos de coral vivo y muerto —sobrecrecidos por céspedes algales—, se observó que los que tenían fragmentos de coral vivo (T2 y T4) tuvieron un mayor crecimiento coralino, a diferencia de los que tenían fragmentos de coral muerto (T1 y T3) para ambas temporadas (Fig. 2). También, se observó un proceso de fusión entre el fragmento vivo implantado y la rama de coral receptora. Nuestros resultados fueron similares a los registrados por Liñán-Cabello et al. (2011) para la misma especie y por Díaz-Pulido et al. (2011) para Acropora intermedia. Ambos trabajos sugieren que este mecanismo de fusión puede ser una estrategia de fijación mecánica para la colonia, lo que promueve la ampliación de espacios para el establecimiento de organismos asociados a los corales.
En los tratamientos T3 y T4 (con lesión apical), los corales que tuvieron mayor crecimiento fueron los de T4 en ambas temporadas (Figs. 3, 4) con respecto a los de la adhesión lateral. Esta respuesta puede ser resultado del corte apical, debido a que este tipo de lesión estimula las “yemas” que crecen mucho más rápido que el resto de las ramas (Reyes-Bonilla, 2003; Reyes-Bonilla et al., 2005). Dicho fenómeno se produce en condiciones naturales cuando el coral se fragmenta o cuando es ramoneado por herbívoros (Guzmán y Cortés, 1993).
 Adhesión lateral de una rama con algas a una rama sana; T2) adhesión lateral de una rama viva a otra rama sana; T3) adhesión apical de una rama con algas en una rama sana; T4) adhesión apical de una rama viva a una rama sana. Datos de la temporada de lluvias (junio a noviembre del 2007).")
Crecimiento de tejido de Pocillopora capitata para los diferentes tratamientos. T1) Adhesión lateral de una rama con algas a una rama sana; T2) adhesión lateral de una rama viva a otra rama sana; T3) adhesión apical de una rama con algas en una rama sana; T4) adhesión apical de una rama viva a una rama sana. Datos de la temporada de lluvias (junio a noviembre del 2007).
 Adhesión lateral de una rama con algas a una rama sana; T2) adhesión lateral de una rama viva a una rama sana; T3) adhesión apical de una rama con algas en una rama sana; T4) adhesión apical de una rama viva a una rama sana. Temporada de secas (de noviembre de 2007 a junio de 2008).")
Crecimiento de tejido de Pocillopora capitata para los diferentes tratamientos. T1) Adhesión lateral de una rama con algas a una rama sana; T2) adhesión lateral de una rama viva a una rama sana; T3) adhesión apical de una rama con algas en una rama sana; T4) adhesión apical de una rama viva a una rama sana. Temporada de secas (de noviembre de 2007 a junio de 2008).
En la temporada de secas se encontró un mayor crecimiento coralino con respecto a la temporada de lluvias, excepto para el T1 de la temporada de secas, con menor crecimiento. Este resultado concuerda con lo hallado para P. damicornis por Guzmán y Cortez (1989). Este hecho, se puede correlacionar con una reducción de la turbidez del agua en la época de secas, lo cual promueve el crecimiento coralino (Roy y Smith, 1971; Jompa y McCook, 2002b; Titlyanov et al., 2007).
Los resultados del presente estudio muestran que el crecimiento del tejido coralino de P. capitata no fue inhibido por el tipo de lesión y adhesión ni por la presencia de céspedes algales, ya que éstos no fueron capaces de colonizar al coral. Esta especie de coral puede ser altamente competitiva como se ha registrado para el resto de las especies del género Pocillopora en el Pacífico mexicano (Reyes-Bonilla, 2003). El coral P. capitata es una especie que responde favorablemente a los trasplantes y a diferentes tipos de manipulación, con una alta tasa de crecimiento (Liñán-Cabello et al., 2011). Además, las especies de la familia Pocilloporidae son usadas para la recuperación de las comunidades arrecifales, debido a su rápido crecimiento (Edwards y Gomez, 2007). El rápido crecimiento y la capacidad de resistir a la manipulación y de resistir al sobrecrecimiento de los céspedes algales, e incluso sobrecrecerlos con tejido coralino nuevo, así como provenir de una familia que contribuye a la recuperación de los arrecifes, sugieren que P. capitata puede ser utilizada en proyectos de restauración para la región de Zihuatanejo. En síntesis, nuestro trabajo contribuye al conocimiento de la diversidad de respuestas encontradas en las interacciones entre corales y algas, donde los corales tuvieron la capacidad de sobrecrecer a las algas y resaltar la capacidad de esta especie de coral para resarcir el deterioro de las comunidades coralinas del Pacífico mexicano.
Estamos en deuda con C. Candelaria, J. M. Oseguera, G. Hernández y N. A. López por la considerable ayuda con el trabajo de campo; con la Dra. M. B. I. Escandón, por la revisión del manuscrito y con I. Ruiz-Boijseauneau por el apoyo técnico brindado en la elaboración de esta publicación. Agradecemos al Programa de Doctorado del Posgrado de Ciencias Biológicas de la UNAM. NACN agradece al Conacyt por la beca de doctorado otorgada. Este manuscrito forma parte de la tesis doctoral de la primera autora y constituye un requisito para la obtención del grado de Doctora en Ciencias del Posgrado en Ciencias Biológicas de la UNAM. Asimismo, los autores agradecen el trabajo cuidadoso y constructivo que hicieron los revisores anónimos y el editor de la revista.