


El chorlo nevado (Charadrius nivosus) es un ave playera amenazada en México y en EUA. La población reproductora de la costa del Pacífico es un segmento poblacional distinto (SPD) que se distribuye desde Washington, EUA, hasta Baja California Sur, México. San Quintín alberga el 50% de la población del SPD de la península de Baja California, por lo cual es prioritario para la especie. Se usaron conteos históricos (1991) y actuales (2007-2014), además de información sobre aves marcadas y recapturadas, para evaluar cambios poblacionales del SPD que anida en la península de Baja California. Se estimó que entre 1991 y 2008 la población peninsular disminuyó un 33%. Sin embargo, la población de San Quintín no siguió esta tendencia y se consideró estable. En invierno, la población residente de chorlos de San Quintín se duplicó por la inmigración de individuos del norte. La abundancia relativa de chorlos en la temporada reproductora, aunque variable, fue mayor en salitrales que en playas arenosas y salinas. En los 2 últimos inviernos la distribución de los chorlos ha cambiado entre playas arenosas y salitrales. La conservación del SPD mexicano depende en un 50% de la protección del hábitat y la conservación de San Quintín.
The snowy plover (Charadrius nivosus) is a shorebird that is threatened in Mexico and US. The breeding population of the Pacific coast is a distinct population segment (DPS) whose distribution goes from Washington, USA, to Baja California Sur, Mexico. San Quintin hosts 50% of the DPS of the Baja California peninsula and, therefore, is a priority site for the species. Historical (1991) and current surveys (2007-2014), plus information on marked and recaptured birds, were used to assess population changes in the DPS nesting in the Baja California peninsula. It was estimated that between 1991 and 2008 the peninsular population declined by 33%. However the population of San Quintin did not follow this trend and it was considered stable. In Winter the resident population of plovers of San Quintin doubled by the immigration of individuals from the north. Density of plovers in the breeding season, although variable, was higher in salt works and sandy beaches. In the last 2 winters there was a shift in the plover distribution between sandy beaches and salt flats. Conservation of the Mexican DPS depends 50% on habitat protection and conservation in San Quintin.
En las últimas décadas han habido disminuciones poblacionales significativas en el 30% de las especies de aves playeras (Charadriiformes) de Norteamérica (Bart, Brown, Harrington y Morrison, 2007; Morrison et al., 2006). Las razones de estas disminuciones no son claras (Butler, Ydenberg, Donaldson y Brown, 2004), pero dado que son varias las especies afectadas y todas tienen en común una distribución amplia durante el periodo no reproductivo, se presume que las causas no son solo locales, sino que tienen una distribución regional o global.
El chorlo nevado (Charadrius nivosus) es una de las especies cuya tendencia poblacional indica que está disminuyendo (Andres et al., 2012). Con un tamaño poblacional estimado para toda Norteamérica de 18,200 individuos, es de las aves playeras menos abundantes en Norteamérica (Thomas et al., 2012) y en el hemisferio occidental (Wetlands International, 2006). Por ello, y por los problemas de conservación de sus hábitats en toda su distribución, esta especie está protegida legalmente con estatus de amenazada a escala federal tanto en México (Semarnat, 2010) como en la costa del Pacifico de los EUA (USFWS, 1993).
En 1993 la población de la costa del Pacífico de EUA se designó legalmente como un segmento poblacional distinto (SPD), debido a su disminución poblacional y a la reducción y degradación de su hábitat de reproducción (USFWS, 1993). Su intervalo geográfico incluye a los individuos que anidan dentro de los 80km de la zona costera, y se extiende hasta Baja California Sur, México. Para la recuperación de esta población se desarrolló un plan de manejo oficial (USFWS, 2007) que actualmente se implementa en 6 unidades de recuperación entre Washington y California. Además, el resto de las poblaciones geográficas también son prioritarias en los planes de conservación de aves playeras de EUA (Brown, Hickey, Harrington y Gill, 2001) y de México (Semarnat, 2008).
Aunque la distribución del chorlo nevado en México es relativamente amplia, pues incluye las costas del Pacífico y del golfo de México, así como los desiertos y llanuras del centro y norte del país (Howell y Webb, 1995; Page, Warriner, Warriner y Paton, 2009), los sitios en donde se reproduce son restringidos (De Sucre-Medrano, Cervantes-Zamudio, Ramírez-Bastida y Gómez del Ángel, 2011; Luévano, Mellink y Riojas-López, 2010; Mellink, Riojas-López y Luévano, 2009; Thomas et al., 2012). En México, la población reproductora se estimó en 2,646 individuos en 2009, lo que representa el 9% de la población total para la especie (Thomas et al., 2012). El 93% de los individuos se encontró en las costas del Pacífico mexicano y el 7%, en el interior del país, y casi el 80% de la población costera mexicana se concentró en el noroeste de México (Thomas et al., 2012).
Los factores que hacen vulnerable al chorlo nevado a los efectos negativos de la pérdida o degradación de la calidad de su hábitat se relacionan, principalmente, con su biología reproductiva, ya que anidan en el suelo y su potencial reproductivo en general es bajo (Page et al., 2009). Además, esta especie usa tanto en la temporada reproductora como en la invernal hábitats discretos y efímeros, y que al mismo tiempo usa el ser humano con fines recreativos o productivos, como las playas y barras arenosas, islas e islotes arenosos y salitrales (Page et al., 2009). En México, los estudios sobre esta especie son escasos, pero demuestran que existe una gran variabilidad interanual en sus números poblacionales y en aspectos de su biología reproductiva, supervivencia y dispersión (Küpper, Cruz-López, Rojas-Abreu, Lozano-Angulo y Serrano-Meneses, 2012). Se han documentado también los efectos negativos de la alteración por la actividad humana y de pérdida del hábitat en esta especie en Sinaloa (Vega-Picos, 2008). No obstante, el estatus actual de las poblaciones de chorlos de la península de Baja California se desconoce, específicamente su distribución espacial y temporal. En este estudio se describen los cambios en la distribución espacial y temporal del SPD del chorlo nevado en la península de Baja California en las últimas 2 décadas, con énfasis en el complejo lagunar San Quintín, el sitio más importante para esta especie en la península. También se analiza la composición estacional de la población de San Quintín y su conectividad con otros sitios del Pacífico tanto en la temporada reproductora como en la invernal.
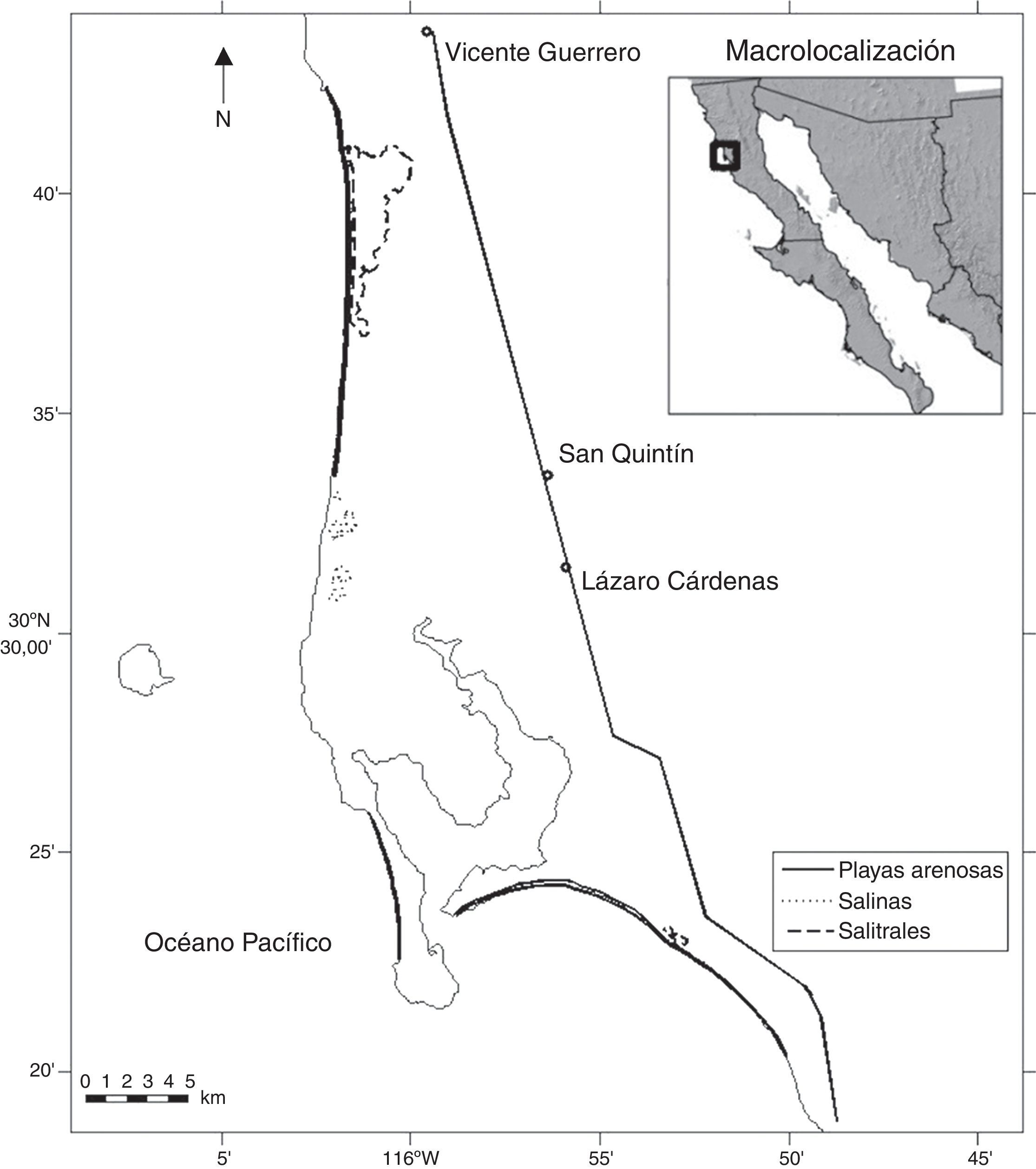
Materiales y métodosPara la evaluación poblacional del SPD que se reproduce en México, se consideraron los 6 humedales más importantes para el chorlo nevado en la costa occidental de la península de Baja California. Estos humedales incluyen el estero de Punta Banda, el complejo lagunar San Quintín, el complejo lagunar Ojo de Liebre-Guerrero Negro, la laguna San Ignacio, San Juanico y Bahía Magdalena. En Massey y Palacios (1994) y Palacios, Alfaro y Page (1994) pueden encontrarse la descripción de estos sitios y un mapa con su localización. El área de estudio donde se enfocó el esfuerzo mayor de este trabajo se denomina complejo lagunar San Quintín, de aquí en adelante referido solo como San Quintín. Este humedal se localiza en el noroeste de Baja California, entre los 30°20’-30°50’ N; 115°45’-116°03’ O (fig. 1). Su área aproximada es de 876.9km2 y comprende planicies costeras, playas arenosas extensas, planicies lodosas, dunas, salinas, salitrales, bahías, desembocaduras de arroyos, marismas y matorral costero. La parte marina del área se compone de 2 cuerpos de agua: Bahía Falsa en el oeste y Bahía San Quintín en el este, con una superficie aproximada de 49.61km2, y separadas parcialmente por una península interna conocida como Monte Ceniza. Bahía Falsa está separada del océano Pacífico por una península externa (Punta Mazo), y hacia el sur de ambas bahías sobresale otra barra arenosa conocida como Punta Azufre. En este estudio, San Quintín también incluyó laguna Figueroa, ubicada al norte del complejo lagunar, la cual es una laguna hipersalina, cerrada, alargada y separada del océano por una barrera de dunas de unos 20km de largo, 150 m de ancho y hasta 12 m de altura. La laguna se extiende tierra adentro entre 0.5 y 3km a partir de esta barrera de dunas (Horodyski, 1977). San Quintín tiene varias denominaciones como sitio de importancia regional dentro la red hemisférica de aves playeras (RHRAP), área de importancia para las aves (IBA No. 13) y humedal de importancia internacional (Ramsar), aunque en esta última el polígono no incluye a laguna Figueroa. En toda el área de estudio los chorlos nevados se distribuyen durante la temporada reproductora e invernal, principalmente en 3 tipos de hábitat: playas arenosas, salitrales y en un conjunto de salinas ubicadas al norte de la bahía (fig. 1) (Palacios y Alfaro, 1991; Palacios et al., 1994).
 en San Quintín, B. C., México.")
En 2007 y 2008 se realizaron conteos en todas las costas y zonas interiores de México en las que existían registros de distribución reproductora histórica o potencial del chorlo nevado, como parte de un esfuerzo conjunto para estimar el tamaño de la población de esta especie en Norteamérica (Thomas et al., 2012). La información sobre las poblaciones del interior (De Sucre-Medrano, Cervantes-Zamudio, Ramírez-Bastida, y Gómez del Ángel, 2011; Luévano et al., 2010) y las costas del macizo continental del Pacífico mexicano ya se publicó (Mellink et al., 2009), pero la relacionada con el SPD que se distribuye en la península de Baja California no fue tratada a detalle por Thomas et al. (2012). Por consiguiente, en este trabajo se incluyeron los datos de 2007 y 2008 para compararlos con los obtenidos hace más de 2 décadas por Palacios et al. (1994) de la península de Baja California.
Por cuestiones logísticas cada sitio de la península se contó una sola vez cada año, y las ventanas de tiempo en las que se realizaron los conteos incluyeron los periodos de monitorización acordados para el SPD; es decir, tercera semana de mayo para los conteos de la población reproductora y enero para la población invernante. Estos periodos concuerdan con el pico de mayor actividad reproductora y los momentos en que las fracciones poblacionales que son migratorias ya se encuentran en sus zonas de invernada (USFWS, 2007). Se realizaron los conteos de la población en San Quintín en 2007 y 2008, y del 2011 al 2014, dentro del periodo designado para el SPD. A partir del invierno 2011-2012 se iniciaron los conteos de la población invernal en San Quintín, en sincronía con el periodo de conteo designado para el SPD.
El método de conteo usado fue igual al de estudios anteriores (De Sucre-Medrano et al., 2011; Luévano et al., 2010; Mellink et al., 2009; Palacios et al., 1994; Thomas et al., 2012), y consistió en coordinar 5 equipos de 1 a 3 personas para realizar muestreos simultáneos en cada sitio. Tanto en el conteo de 2007 como en el de 2008 participaron 5 equipos con 1 observador experimentado cada uno, que censaron 14 localidades en Baja California y 52 en Baja California Sur. Se anotó el número de chorlos nevados encontrados en los recorridos a pie o en vehículos todoterreno, en cada uno de los hábitats. Con ayuda de binoculares (10×) y telescopios (20-60×) se identificó y determinó la clase de edad de los individuos de acuerdo con Page et al. (2009). Durante la temporada reproductora fue relativamente fácil determinar el sexo de la mayoría de los individuos sin capturarlos; pero en el invierno, en condiciones adversas de luz y a gran distancia, el sexo de la mayoría de los individuos no pudo determinarse. Para comparar entre las temporadas de reproducción e invernada y con información proveniente de otros estudios, solo nos referimos a la población de adultos sin importar su sexo.
El área potencial del hábitat que usa el chorlo nevado en San Quintín se estimó en Google Earth® y la validamos en campo con base en la localización de los nidos y áreas de alimentación y descanso durante las temporadas reproductora e invernal. En todas las temporadas reproductoras estimamos que el área fue constante para playas arenosas, salitrales y salinas (245.3, 510 y 34.04ha, respectivamente). Sin embargo, en la temporada invernal el área de alimentación disponible para los chorlos aumentó en los salitrales por la formación de charcas producto de una precipitación mayor en los últimos 2años (esto es, 2011-2012 con 360ha; 2012-2013 con 410ha y 2013-2014 con 510ha).
Captura, marcaje y monitorización de individuos marcados en San QuintínDurante las temporadas reproductoras 2012, 2013 y 2014 se buscaron nidos de la especie en los 3 tipos de hábitat, y se capturaron adultos en el nido usando trampas de embudo (Székely, Argüelles-Ticó, Kosztolányi y Küpper, 2011). Se referenció geográficamente cada nido usando un dispositivo de posicionamiento global (GPS). Todos los individuos capturados fueron sexados de acuerdo con Page et al. (2009) y marcados con combinaciones únicas de anillos de colores tipo Darvic. Además, para evitar confusiones con individuos que hubiesen sido marcados en EUA durante las últimas 3décadas (Page, Stern y Paton, 1995; Stenzel et al., 1994; Stenzel et al., 2007) o en México durante los últimos 8 años (De Sucre-Medrano et al., 2011; Küpper et al., 2012), a cada ave se le colocó un anillo de metal sobre uno de color azul cielo en el tarso izquierdo como indicativo de que fue marcada en San Quintín. La captura y marcaje de las aves fue autorizada por la Dirección General de Vida Silvestre (oficio Núm. SGPA/DGVS/02078/12 y oficio Núm. SGPA/DGVS/06485/13).
Aunque se empezaron a marcar chorlos en el 2012, la búsqueda de individuos anillados se realizó desde el invierno de 2011-2012, ya que existían antecedentes de chorlos nevados de EUA que invernan en la costa oriental y occidental de la península de Baja California, incluyendo San Quintín (Page et al., 1995; Stenzel et al., 2007). Se realizó la búsqueda a pie o en vehículos todoterreno en los 3 tipos de hábitat, y se usaron binoculares y telescopios para leer las combinaciones de anillos de los individuos a una distancia máxima de 30m. Se registró con un GPS la posición de cada individuo marcado, y determinamos su sexo (macho o hembra), que fue corroborado posteriormente con la información proveniente de su captura. En el invierno el monitorización fue intenso (media=7h/día); se dedicaron 5 días en 2011-2012, 10 días en 2012-2013 y 15 días en 2013-2014. Durante la temporada reproductora el monitorización fue menos intenso (media=2.5h/día), pero la búsqueda se prolongó hasta 63 días en 2012 y 2014 y 65 días en 2013. Para el invierno, el número de «recapturas» se refiere exclusivamente al número de individuos marcados que fueron avistados, pero para la temporada reproductora incluye a individuos marcados que fueron solamente avistados y a aquellos que fueron capturados en sus nidos por segunda vez.
Para determinar la estacionalidad (residente o migratorio) de la población, a cada chorlo nevado marcado y sexado se le asignó un estatus de estacionalidad según su historial de recaptura. Los chorlos residentes fueron aquellos recapturados en 1) el invierno posterior a su captura, o 2) en el invierno posterior a su captura y en la siguiente temporada reproductora. Los chorlos migratorios fueron aquellos recapturados exclusivamente en la temporada reproductora posterior a su captura, y que no fueron detectados en el área durante el invierno. Es decir, individuos que llegan solo para reproducirse en San Quintín y que probablemente pasan el invierno en otra área. De esta forma, la información sobre recapturas se dividió en: 1) un primer ciclo anual que considera a las aves marcadas en la temporada reproductora 2012 y recapturadas durante el invierno 2012-2013 y hasta la temporada reproductora 2013, y 2) un segundo ciclo anual que incluye a individuos marcados en la temporada reproductora 2013 y recapturados durante el invierno 2013-2014 y hasta la temporada reproductora 2014. Así, al inicio de cada ciclo se tuvo una muestra independiente de individuos marcados por primera vez.
La información de las recapturas de la población marcada en San Quintín fue complementada con avistamientos realizados durante visitas esporádicas en el invierno a otros sitios de la península de Baja California o de registros de observadores en California, EUA. El sitio y fecha de captura de los chorlos nevados marcados en otros sitios y observados durante las temporadas de invierno en San Quintín se obtuvo de la base de datos de Point Blue Conservation Science (F. Bidstrup, comunicación personal).
Para establecer la tendencia numérica de la población reproductora en San Quintín, usamos la prueba de tendencia de Mann-Kendall (Mann, 1945) para probar si el número de chorlos nevados adultos era independiente del tiempo. Se compararon la proporciones de chorlos nevados residentes y migratorios, sin importar su sexo, entre años con una prueba X2 de independencia. De las recapturas del 2013-2014 se excluyó a todos los individuos marcados en la temporada reproductora previa (2012), porque nuestro interés principal era explorar si la composición de la población, en cuanto a su estatus de estacionalidad, variaba anualmente. De esta manera, se realizó el seguimiento de 2 muestras independientes que iniciaron con individuos capturados por primera vez. También se usó una X2 de independencia para probar si el estatus de estacionalidad (residente o migratorio) era independiente del sexo de los individuos. Se aplicaron pruebas X2 para determinar si la abundancia relativa de chorlos nevados era homogénea entre hábitats y años. Este análisis se llevó a cabo por separado para las temporadas de reproducción e invernal. Todos los análisis estadísticos se realizaron en el ambiente de programación R (R Development Core Team, 2007), con un nivel de significación del 0.05.
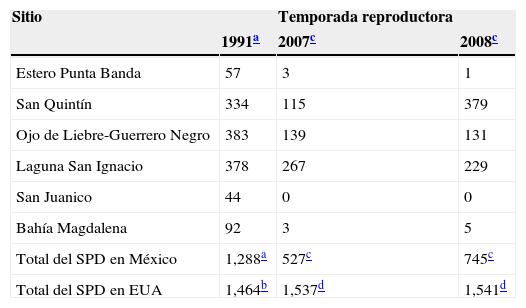
ResultadosEn 2007 y 2008 se realizó un conteo de la población de chorlos nevados en los sitios más importantes de la costa occidental de la península de Baja California durante la temporada reproductora. Comparamos estos resultados con los de 1991, único conteo histórico disponible para los mismos sitios. La población de chorlos en la península disminuyó un 59% entre 1991 y 2007 (tabla 1). Para validar este resultado, en 2008 se repitió el conteo en todos los sitios, y la disminución poblacional con respecto a 1991 fue del 42%. La diferencia del tamaño poblacional entre 2007 y 2008 fue del 29%, y se debió principalmente al incremento de la población de San Quintín en 2008. En los demás sitios el tamaño poblacional se mantuvo en niveles bajos, e incluso disminuyó aún más en 2008 (tabla 1). En 1991, la proporción del SPD en la península de Baja California era similar a la de EUA, pero disminuyó del 47% en 1991 al 33% en 2008, básicamente por una tendencia negativa en México y una población estable en EUA, al menos durante el periodo de estudio (tabla 1).
Número de adultos de chorlo nevado (Charadrius nivosus) del SPD de la península de Baja California, México y EUA.
| Sitio | Temporada reproductora | ||
|---|---|---|---|
| 1991a | 2007c | 2008c | |
| Estero Punta Banda | 57 | 3 | 1 |
| San Quintín | 334 | 115 | 379 |
| Ojo de Liebre-Guerrero Negro | 383 | 139 | 131 |
| Laguna San Ignacio | 378 | 267 | 229 |
| San Juanico | 44 | 0 | 0 |
| Bahía Magdalena | 92 | 3 | 5 |
| Total del SPD en México | 1,288a | 527c | 745c |
| Total del SPD en EUA | 1,464b | 1,537d | 1,541d |
SPD: segmento poblacional distinto.
En algunos sitios la disminución numérica de los chorlos adultos fue tan grave que estas poblaciones se consideran casi extirpadas. Estos sitios incluyen al estero de Punta Banda, San Juanico y Bahía Magdalena, cuyas poblaciones disminuyeron de entre 44 a 92 individuos a menos de 5 por sitio. La población de chorlos reproductores también disminuyó en los 2 sitios de la Reserva de la Biósfera El Vizcaíno. Entre 1991 y 2008, la disminución fue del 66% en el complejo lagunar Ojo de Liebre-Guerrero Negro, y del 39% en la laguna San Ignacio (tabla 1).
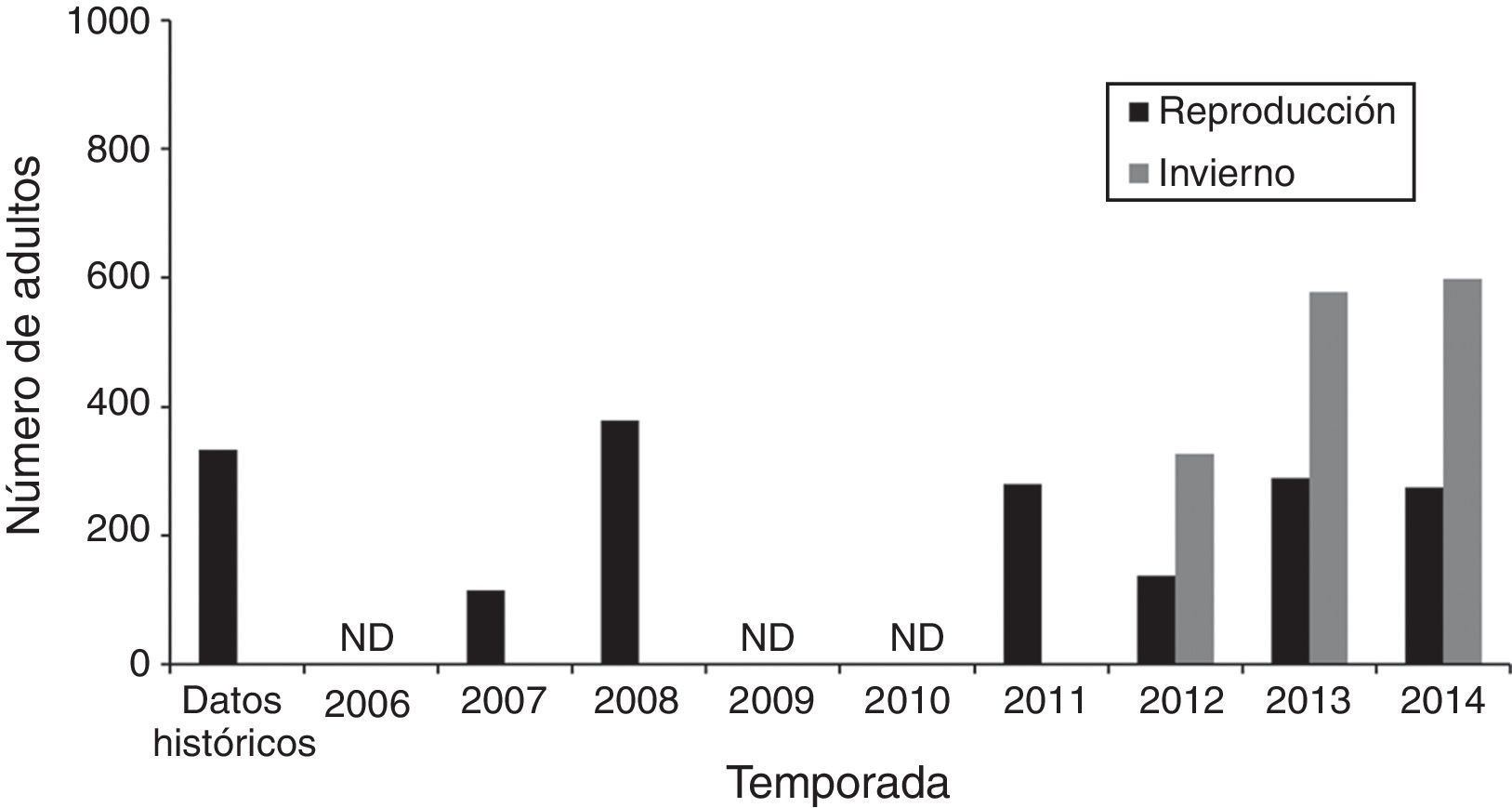
En San Quintín se encontró a la población reproductora más importante del chorlo nevado en la península de Baja California. El máximo histórico de 379 adultos representa el 51% del SPD que se reproduce en México (tabla 1). No obstante, durante las 7 temporadas reproductoras el número de adultos registrado varió anualmente (fig. 2). En 5 temporadas se observaron variaciones menores con respecto a la referencia histórica (1991), pero en 2 (2007 y 2012) se registraron disminuciones del 65 y el 50%, respectivamente, con respecto al conteo inmediato anterior, e incrementos de la misma magnitud en 2008 y 2013 (fig. 2). En comparación con el conteo de 1991 (334 adultos), solo en el 2008 se registró un número mayor de adultos, y la tendencia de la población de chorlos en San Quintín fue estable (prueba de tendencia de Mann-Kendall τ= –0.14; n=7; p=0.76).
 adultos en San Quintín, B. C., México, durante la temporada reproductora e invernal. El conteo de 1991 (Palacios et al., 1994) es la referencia histórica para San Quintín, B. C.")
Abundancia de chorlos nevados (Charadrius nivosus) adultos en San Quintín, B. C., México, durante la temporada reproductora e invernal. El conteo de 1991 (Palacios et al., 1994) es la referencia histórica para San Quintín, B. C.
En la temporada reproductora 2012 se marcaron 77 adultos (48 hembras y 29 machos), mientras que en 2013 fueron 72 (44 hembras y 28 machos). Este sesgo hacia las hembras capturadas en ambas temporadas no está relacionado con la estructura de la población sino con el hecho de que las hembras incuban de día, mientras que los machos incuban de noche. Esto ocasiona que la captura de hembras, diurna, sea más fácil que la de machos, nocturna. De hecho, con base en la información de los conteos en las 7 temporadas reproductoras, se estimó que la proporción macho: hembra fue en promedio de 1.5:1.
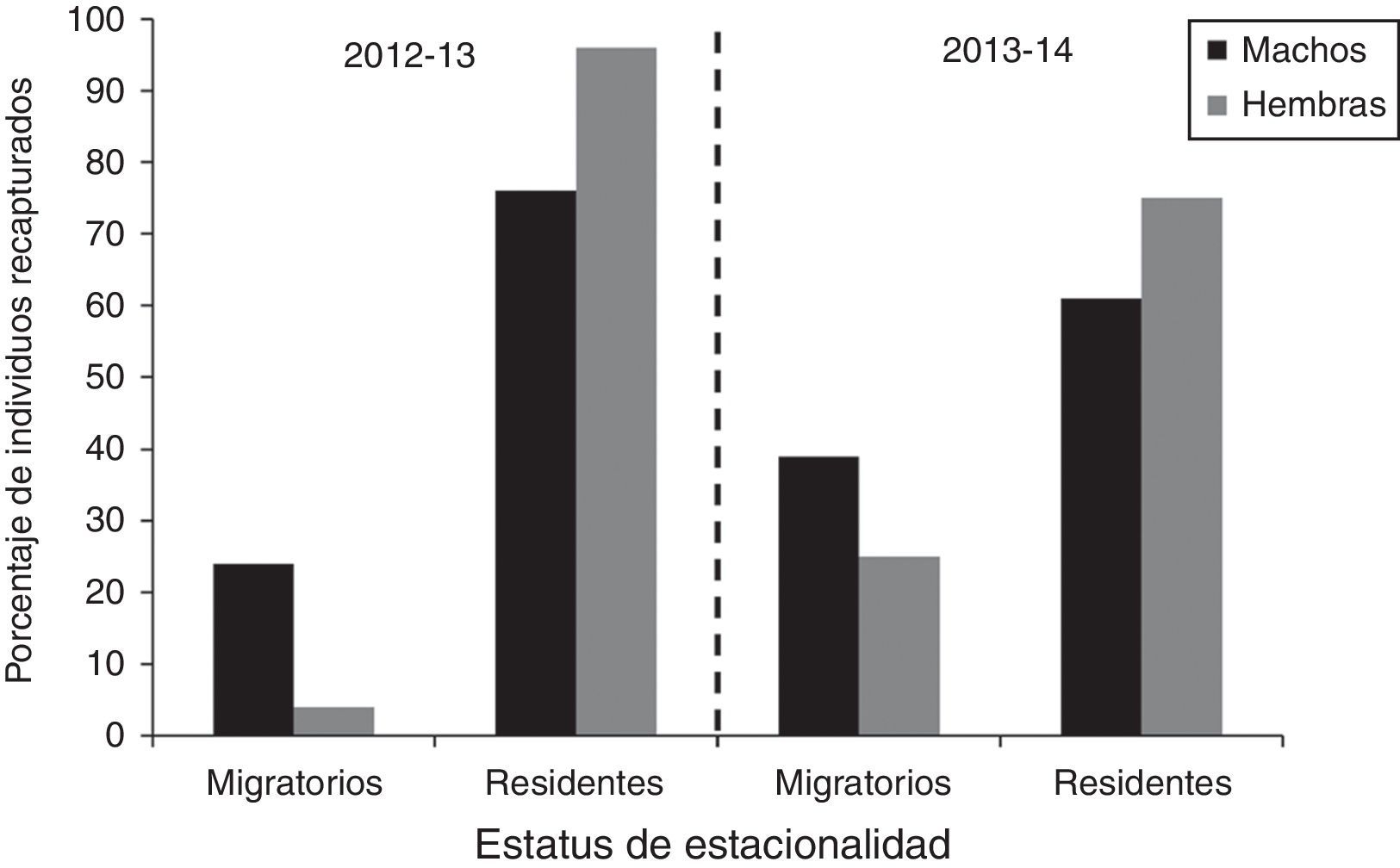
En total se recapturó al 67% de los individuos marcados en 2012 y 2013 (58 y 76%, respectivamente). En ambos años, la proporción de hembras recapturadas con respecto al total marcado (58 y 73%) fue similar a la de machos (59 y 82%). Por otra parte, basándose en el número de individuos recapturados en cada ciclo anual se calculó la proporción de chorlos nevados migratorios y residentes. En general, la mayoría de los individuos se comportaron como residentes tanto en el ciclo 2012-2013 como en 2013-2014 (fig. 3), aunque la proporción de residentes varió entre años. En el ciclo 2012-2013 el 89% de los individuos fue residente, mientras que en el 2013-2014 lo fue el 69% (X2=12.05; 1g. l.; p<0.001). El estatus de estacionalidad también fue diferente entre los sexos en ambos ciclos (fig. 3). La proporción de hembras residentes fue mayor que la de machos, mientras que la proporción de machos migratorios fue mayor que la de hembras (ciclo 2012-2013: X2=16.61; 1g. l.; p<0.001; 2013-2014: X2=4.50; 1g. l.; p=0.03).
 recapturados según su estatus de estacionalidad en San Quintín, B. C., México, en los ciclos anuales 2012-2013 y 2013-2014.")
La población invernal aumentó más del 40% en las últimas 2 temporadas con respecto al primer conteo de invierno (2011-2012). Entre la temporada reproductora 2011 y el invierno 2011-2012 el número de adultos no aumentó, pero entre las temporadas reproductoras 2012 y 2013 y los inviernos 2012-2013 y 2013-2014, respectivamente, se observaron aumentos de más del 50% (fig. 2). En el primer invierno (2011-2012) no se detectó a ningún chorlo nevado que hubiera sido marcado fuera de San Quintín, aunque el esfuerzo de búsqueda fue el más bajo de todos los inviernos (5 días). En el invierno siguiente se avistó a 5 individuos del SPD de EUA (California) y uno de la población interior (Gran Lago Salado, Utah). En el último invierno (2013-2014) se avistaron 9 individuos del SPD (8 de California y uno de Oregón). Dos de los individuos de California también fueron avistados en el invierno anterior. En la temporada reproductora 2014 solo se detectó en San Quintín a un individuo marcado en California.
De las aves marcadas en San Quintín, solo una hembra capturada en la temporada reproductora 2012 fue vista 2meses después en una playa de San Diego, California. Esta hembra se observó en repetidas ocasiones durante 2013 en el mismo sitio en San Diego, donde tuvo un registro de anidación exitoso el 8 de agosto de 2014. Por otro lado, otra hembra capturada en la temporada reproductora 2013, pasó el invierno en Santa Rosalillita, B. C., unos 300km al sur de San Quintín (observada el 22 de enero de 2014), y regresó para reproducirse exitosamente al área de estudio en la temporada reproductora 2014.
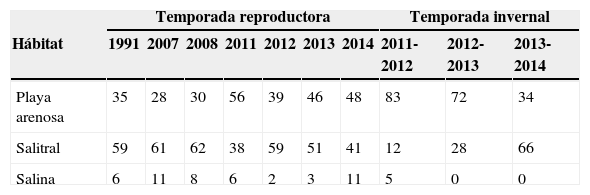
La abundancia relativa de los chorlos nevados por hábitat en San Quintín fue variable en cada temporada reproductora (X2=37.89; 12g.l.; p<0.001), particularmente en las salinas (tabla 2). La abundancia relativa de adultos fue mayor en los salitrales (mediana=59) que en las playas arenosas (mediana=39) y salinas (mediana=6) (tabla 2). También en el invierno la abundancia relativa de chorlos nevados en cada hábitat fue diferente (X2=74.53; 4g.l.; p<0.001). Los hábitats preferidos por los chorlos fueron las playas arenosas y salitrales, mientras que las salinas solo fueron usadas en el invierno de 2011-2012. En las últimas 2 temporadas invernales la importancia de las playas disminuyó en un 47%, mientras que la de los salitrales aumentó en un 58% (tabla 2).
Abundancia relativa de chorlos nevados (Charadrius nivosus) adultos por hábitat, durante las temporadas de reproducción e invernada en San Quintín, Baja California, México.
| Temporada reproductora | Temporada invernal | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Hábitat | 1991 | 2007 | 2008 | 2011 | 2012 | 2013 | 2014 | 2011-2012 | 2012-2013 | 2013-2014 |
| Playa arenosa | 35 | 28 | 30 | 56 | 39 | 46 | 48 | 83 | 72 | 34 |
| Salitral | 59 | 61 | 62 | 38 | 59 | 51 | 41 | 12 | 28 | 66 |
| Salina | 6 | 11 | 8 | 6 | 2 | 3 | 11 | 5 | 0 | 0 |
Con esta investigación se documentó la disminución poblacional del SPD que anida en la península de Baja California. Entre 1991 y 2007/2008 el cambio en las poblaciones de los 6 sitios más importantes para el chorlo nevado en la región fue negativo. La extensión geográfica de esta disminución poblacional fue amplia, lo que hace pensar que las causas subyacentes estuvieron distribuidas también de forma amplia. En condiciones naturales las aves playeras son limitadas por el alimento o el hábitat en cualquier momento de su ciclo anual, por lo que la pérdida o degradación de su hábitat tiene consecuencias negativas en sus poblaciones (Colwell, 2010). La alteración por la actividad humana es una forma de degradación de la calidad del hábitat (Gill y Sutherland, 2000) que engloba varias actividades humanas que tienen el potencial de afectar negativamente el éxito reproductivo y la supervivencia y, en consecuencia, reducir los tamaños poblacionales de las aves playeras.
Entre 1991 y 1992, Palacios et al. (1994) documentaron que en solo 3 de las 16 localidades visitadas en toda la península de Baja California encontraron evidencia de tráfico intenso de vehículos todoterreno. En este estudio se visitaron las mismas localidades que Palacios et al. (1994), y en todas fue evidente que el tránsito de vehículos es más intenso, e incluso en áreas de difícil acceso el impacto humano es evidente por la presencia de basureros clandestinos (observación personal). Varios estudios de chorlos han demostrado una correlación negativa entre el uso del hábitat y el éxito reproductivo con la alteración por la actividad humana (Burger, 1994; Flemming, Chiassion, Smith, Austin-Smith y Bancroft, 1998; Lafferty, Goodman y Sandoval, 2006; Ruhlen, Abbot, Stenzel y Page, 2003). La pérdida o degradación del hábitat de varias especies de aves playeras debido al crecimiento poblacional y al desarrollo de actividades productivas como la agricultura y pesca, ha venido acompañado por nuevas comunidades de depredadores asociadas a estos paisajes alterados. Los cambios en la composición y abundancia de los depredadores en estas comunidades se han asociado con la disminución de la productividad de muchas poblaciones de aves playeras (Helmers y Gratto-Trevor, 1996). Por ejemplo, las tasas de depredación altas en hábitats costeros se han atribuido al incremento de las poblaciones de depredadores naturales (coyotes, cuervos) e introducidos (ratas, perros y gatos domésticos o asilvestrados) que son atraídos por fuentes de alimento alternativas; por ejemplo, basureros clandestinos cerca de los centros poblacionales humanos (Elliott-Smith y Haig, 2004).
En aquellos sitios con los cambios poblacionales negativos más extremos tales como el estero de Punta Banda, San Juanico y Bahía Magdalena, los cambios en la cantidad del hábitat potencial para los chorlos no son significativos; sin embargo, la calidad del mismo sí se ha degradado. Por ejemplo, en el estero de Punta Banda se recolectan las macroalgas varadas en la playa arenosa con ayuda de maquinaria pesada, con fines comerciales (S. González, comunicación personal; observación personal). En algunas playas de California, EUA, este tipo de actividad reduce significativamente la riqueza, abundancia y biomasa de muchas especies de invertebrados y, por ende, disminuye la abundancia de aves playeras como el chorlo nevado que se alimenta de los invertebrados asociados a las macroalgas varadas (Dugan, Hubbard, McCrary y Pierson, 2003). En la playa de bolsillo de San Juanico donde el chorlo nevado está extirpado como especie reproductora, el uso del hábitat por el ser humano ha aumentado a tal grado que no hay espacio disponible para la anidación de los chorlos por el número de embarcaciones varadas, tránsito de vehículos de todo terreno y construcciones en esa playa, además de las actividades pesqueras y recreativas asociadas. En Bahía Magdalena, las playas arenosas y barras arenosas son muy extensas y ofrecen hábitat de anidación para los chorlos. Sin embargo, estas playas son usadas por los pescadores como vías de comunicación entre campos pesqueros distantes, como entre Cabo San Lázaro y Puerto Adolfo López Mateos, y entre Puerto Magdalena y el campo pesquero de Bahía Santa María. Otras especies como el ostrero americano (Haematopus palliatus) y el gallito marino californiano (Sternula antillarum) también han disminuido su presencia en el área (E. Amador, comunicación personal).
La baja numérica que se observó en los chorlos nevados del SPD de la península de Baja California sucedió tanto en áreas protegidas como en áreas sin protección, en áreas remotas como en áreas cercanas a centros urbanos importantes. Por lo tanto, esto hace descartar la idea de que la alteración por la actividad humana sea la única causa de las disminuciones en las poblaciones reproductoras del chorlo nevado. Es probable que, cuando menos, la disminución en la temporada reproductora 2007 en toda la península de Baja California también esté relacionada con lo sucedido a todo el SPD en EUA (USFWS, 2014), y que se especula se debió a la supervivencia baja de los chorlos nevados en el invierno 2006-2007 por condiciones climáticas severas, como tormentas y episodios de temperaturas muy bajas (J. Watkins, comunicación personal). Por ejemplo, en Humboldt, California, en enero de 2007 se registraron más de 20 días con temperaturas por debajo de los 2.29°C que ocasionaron una supervivencia aparente baja en los chorlos nevados de esa población (Eberhart-Phillips, 2012; Eberhart-Phillips y Colwell, 2014). En otro estudio llevado a cabo en California, la desaparición del 19% de una población marcada de chorlos nevados se asoció con un episodio de temperaturas bajo cero durante una semana en diciembre de 1990 (Stenzel et al., 2007). De forma similar en San Quintín, en enero y febrero de 2007, se registraron temperaturas mínimas promedio de 2.6-2.8°C (CLICOM, 2014) que pudieron haber ocasionado una disminución de la población de adultos en la temporada reproductora de ese año. En comparación con 1991, la referencia histórica de este estudio, la temperatura mínima promedio en los mismos meses varió entre 5.1 y 6.8°C (CLICOM, 2014). No obstante, el efecto del clima en las disminuciones poblacionales de los chorlos nevados de San Quintín no es claro. En enero y febrero de 2008 las temperaturas mínimas promedio fueron incluso más bajas que en 2007 (1.8-2.3°C; CLICOM, 2014), y sin embargo, la población de adultos en la temporada reproductora 2008 aumentó y fue 3 veces mayor que en 2007. En el resto de los sitios de la península de Baja California (tabla 1) y del SPD de EUA el número de adultos varió muy poco de 2007 a 2008 (USFWS, 2014). No se dispone de datos que muestren si el incremento observado en San Quintín en 2008 se debió al reclutamiento de nuevos individuos a la población local, al arribo de individuos migratorios que solo llegaron a reproducirse, o a ambos.
Los datos climáticos del periodo invernal 2010-2011 hasta el presente no están disponibles y no pudieron relacionarse con las variaciones en el número de adultos entre las temporadas reproductoras de 2011 y 2013 (fig. 2). La disminución poblacional en 2012 (∼50% menos que en 2011) pudo haber estado relacionada con una disminución en la inmigración de chorlos reproductores hacia San Quintín. En esa temporada, el 89% de los individuos en la población fue residente, mientras que el componente migratorio fue exiguo (fig. 3); por lo tanto, tal disminución no pudo ser amortiguada por inmigración. Otra evidencia que apoya esta interpretación fue el hecho de que una hembra con registro de anidación en San Quintín en julio de 2012, emigró 2 meses después hacia San Diego, California, y desde entonces tiene registros de invernada y reproducción en esa zona.
El incremento de aproximadamente el 50% que se observó en 2013 con respecto a 2012 no estuvo relacionado con el reclutamiento de juveniles en la población local, ya que el éxito de anidación en la temporada reproductora 2012 fue muy bajo (11.9%), y se tiene registro de solo un pollo de 63 marcados en esa temporada que sobrevivió hasta su primer invierno (D. Galindo-Espinosa, datos no publicados). Tal incremento probablemente se debió a la llegada de un número mayor de individuos migratorios que solo se reproducen en San Quintín, ya que en el ciclo 2013-2014 la población migratoria fue un 30% más grande que en el ciclo anterior (fig. 3).
La estabilidad de la población reproductora de 2013 a 2014 (fig. 2) puede explicarse por una presencia mayor del componente migratorio (fig. 3), por el éxito mayor de anidación en 2013 (38.5%) y el subsecuente reclutamiento de juveniles en la población local (por ejemplo, 4 de 89 pollos marcados en 2013 se observaron anidando o en cortejo en la temporada reproductora 2014; D. Galindo Espinosa, datos no publicados).
Para la temporada de invierno se conocía muy poco sobre la distribución y abundancia del chorlo nevado en la península de Baja California. Aunque existen programas de monitorización de aves playeras en el invierno, generalmente y atendiendo a los hábitos de alimentación de este grupo de aves, los conteos se realizan principalmente en planicies lodosas (Page et al., 1997); sin embargo, los chorlos nevados en el invierno se distribuyen principalmente en playas arenosas y salitrales, por lo que en general su abundancia es subestimada (Danemann, Carmona y Fernández, 2002; Page et al., 1997). Con base en este estudio (fig. 2), además de lo observado en todas las poblaciones de la costa del Pacífico de EUA (USFWS, 2014), ahora se sabe que durante el invierno la población reproductora de chorlos se duplica en San Quintín en algunos años, y probablemente lo mismo ocurra en todos los demás sitios de la península de Baja California. Por ejemplo, en septiembre de 2005 se contaron 16 chorlos nevados adultos y 7 pollos en una playa arenosa de Santa Rosalillita, B. C., y en noviembre del mismo año se contaron 115 adultos, incluyendo a un individuo que fue marcado en el condado de San Diego, California (F. Bidstrup, comunicación personal).
Estudios previos han establecido que algunas fracciones migratorias de los chorlos nevados de la costa del Pacífico de EUA y de sitios del interior de California (Mono Lake) y Oregón (Lake Abert) invernan en la costa occidental de la península de Baja California (Page, Bidstrup, Ramer y Stenzel, 1986; Page et al., 1995). Este trabajo confirma que tal patrón se mantiene, ya que 12 de los 13 individuos que avistamos durante el invierno provienen de las poblaciones costeras de EUA. También se conocía que algunos chorlos nevados de la población interior más grande en Norteamérica (Gran Lago Salado, Utah; Thomas et al., 2012) pasan el invierno principalmente en ambas costas del golfo de California y en la costa occidental de Baja California Sur (Page et al., 1986), pero este estudio provee evidencia de que la costa occidental de Baja California es también un sitio de invernada para los individuos de esa población. En general, el aumento de la población invernante en San Quintín y en otros sitios de la costa occidental de la península de Baja California se debe principalmente a la influencia de poblaciones norteñas de la costa del Pacífico y marginalmente del interior de EUA. La información proveniente de 2 series de conteos navideños de aves realizados en Ensenada, B. C. (13 conteos), y en La Paz, B. C. S. (9 conteos), muestra que la tendencia poblacional de los chorlos nevados es estable o está ligeramente aumentando (National Audubon Society, 2014). Por consiguiente, durante el invierno, la composición de la población es diferente de la temporada reproductora, y está modulada por la inmigración de otras poblaciones norteñas que provienen del SPD y del interior del continente de EUA. Además, la observación de una hembra capturada en San Quintín durante la temporada reproductora 2013 y observada unos 300km al sur en el invierno siguiente, sugiere que dentro de la península también existen movimientos migratorios a menor escala que probablemente repercuten en el tamaño de la población invernante en cada sitio.
En San Quintín los chorlos nevados se distribuyeron principalmente en salinas, playas arenosas y salitrales. Aunque algunos chorlos también usan las planicies lodosas solo para alimentarse cuando hay mareas muy altas que limitan el hábitat disponible en playas arenosas, su uso es marginal y no se consideró en este estudio. Para la temporada reproductora, la abundancia relativa de chorlos se relacionó directamente con la extensión de cada hábitat, lo que refleja los hábitos territoriales de esta especie (Page et al., 2009). Puesto que los salitrales poseen la extensión mayor, albergan a la mayor parte de la población reproductora de chorlos.
Durante el invierno, los chorlos muestran hábitos gregarios (Page et al., 2009), y la importancia de las salinas fue casi insignificante, mientras que el hábitat de playa arenosa tuvo la abundancia relativa mayor de individuos (tabla 2). En el invierno las aves playeras seleccionan hábitats cuyos atributos (por ejemplo, poca vegetación, abiertos) reducen el riesgo de depredación (Fernández y Lank, 2006; Pomeroy, 2006). En comparación con las playas arenosas y salitrales, las salinas en San Quintín conforman un hábitat más cerrado y con rasgos fisiográficos como lomeríos, arbustos de tamaño mediano y estructuras como postes de cercos, que pueden ser percibidos por los chorlos como un hábitat más riesgoso. También durante el invierno la distribución espacial de las aves playeras está correlacionada con la distribución y disponibilidad del alimento (Brindock y Colwell, 2011; Colwell y Landrum, 1993). En este estudio se observó que la abundancia relativa de chorlos nevados en los salitrales aumentó en las últimas 2 temporadas invernales, mientras que en las playas arenosas disminuyó (tabla 2), pero el número total de chorlos nevados invernantes fue constante en San Quintín entre esos 2 inviernos (fig. 2). Esto sugiere un cambio en la distribución entre los hábitats de playas arenosas y salitrales que puede estar relacionado con el incremento en el área de alimentación disponible para los chorlos en los salitrales, debido a una mayor frecuencia e intensidad de lluvias invernales en esos años.
El hábitat de salitrales es el más extenso y albergó a la mayor parte de la población de chorlos. El 97.6% de este hábitat se localiza en laguna Figueroa, cuyo hábitat carece de protección. El hecho de que laguna Figueroa albergue una población importante de una especie amenazada brinda una oportunidad para lograr la protección legal específica de este lugar bajo la designación de Hábitat Crítico Acuático (Ley General de Vida Silvestre; Artículos 64 y 65), lo cual podría ser complementario al decreto de área natural protegida (ANP) que se está proponiendo para la bahía de San Quintín. Aunque la Bahía de San Quintín es un sitio Ramsar, el polígono no incluye a laguna Figueroa. En este sentido, la información generada en este estudio es un dato importante para justificar la propuesta de creación de área protegida y para desarrollar su programa de manejo.
Agradecemos al Conacyt por la beca de doctorado otorgada a D. Galindo-Espinosa (No. 99866). El trabajo de campo fue posible gracias al apoyo en distintas etapas de L. Alfaro, E. Amador, R. Arreola, M. Cruz, A. De Sucre, E. Fernández, J. Girón, S. Gómez del Ángel, S. González, C. Küpper, E. Palacios-Alfaro, F. Puebla, A. Romo-Piñera, J. Vargas y E. Zamora. A M. A. Cosío por realizar el mapa del área de estudio y a F. Bidstrup (Point Blue Conservation Science) por la información sobre las aves marcadas en EUA. Agradecemos los comentarios de 2 revisores anónimos que ayudaron a mejorar este trabajo. El financiamiento para realizar esta investigación fue otorgado por el United States Fish and Wildlife Service (Neotropical Bird Conservation Act, FWS Agreement No.: MX-N691), Sonoran Joint Venture, Copper River International Migratory Bird Initiative (United States Forest Service-International Programs), Point Blue Conservation Science, Terra Peninsular, A. C., Pronatura Noroeste, A. C., Centro de Investigación Científica y de Educación Superior de Ensenada y Tracy Aviary.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.