



Black bears (Ursus americanus) are considered endangered in Mexico. In northern Mexico, bears are considered responsible for preying on livestock, damaging private property, and crops; therefore they are killed to protect locals' livelihoods. There is a need to understand bears' ecological requirements in order to create conservation plans that include human-bear conflict resolution and that reduce the hunting of this endangered species. We studied the diet of black bears in the Sierra de Picachos located in the Northeast portion of the State of Nuevo León, Mexico. Our objective was to determine the diet of black bears during 3 consecutive seasons (i.e., dry, rainy, and post-rainy), and to determine the presence and frequency of livestock and crops remains in bear scats. We collected 169 scats from December 2004 to November 2005, and estimated diet composition using percentage of occurrence and volume of each food item. Plants were the most common item (73.4%), animals (16.8%), and undetermined wood fragments (9.3%). There was no statistically significant difference among seasons. Our analysis also failed to find cattle residues in any of the scats; therefore, black bears in the study site either do not or rarely feed upon livestock. We recommend that management plans include informing local people on bear behavior. Also, specific actions to avoid bear proximity should be implemented along with government actions aimed at improving bear habitat that minimize bear-human conflict in the area.
El oso negro (Ursus americanus) es considerado en peligro de extinción en México y se cree que depreda ganado, destruye propiedad privada y cultivos, por lo que los pobladores locales los cazan para proteger su subsistencia. Existe la necesidad de investigar los requerimientos ecológicos de este úrsido para crear planes de conservación que minimicen conflictos entre humanos y osos. Se estudió la dieta del oso negro en la sierra de Picachos, ubicada en la región noreste de Nuevo León, México. Nuestro objetivo fue determinar la dieta del oso negro durante 3 temporadas (secas, lluvias y post-lluvias) y determinar la presencia y frecuencia de ganado y cultivos en los excrementos. Se recolectaron 169 excrementos de diciembre del 2004 a noviembre del 2005 y se estimó la composición de su dieta en porcentaje de presencia y en volumen de cada alimento. Las plantas fueron los elementos más comunes (73.4%), seguido de los animales (6.8%) y fragmentos sin determinar (9.3%). No hubo diferencias estadísticamente significativas entre estaciones. No hubo señales de ganado en las heces. Se recomienda un plan de gestión que incluya información a la población local sobre el comportamiento del oso y acciones específicas para mejorar el hábitat de estos animales y así minimizar los conflictos entre humanos y osos.
Black bears (Ursus americanus) are considered endangered in Mexico (Nom-059-Semarnat-2010) despite their status, they are threatened by fragmentation, loss of habitat, and poaching (Medellin et al., 2005). Black bears are known to feed on a variety of vegetation, insects and other animals (Holcrof and Herrero, 1991; Bull and Wertz, 2001). They have an opportunistic foraging behavior, and they are influenced by the availability of their food sources (Pelton, 2003). Black bear diet studies in Canada (MacHutchon, 1989), the US (Bull and Wertz, 2001; Greenleaf et. al., 2009), and Mexico (Sierra-Corona et al., 2005; Niño-Ramírez 1989; Doan-Crider, 1995) have shown bears eating mostly plants and insects, with a lower percentage of vertebrates, from <3% in Yukon (MacHutchon, 1989) to >28% in the Rocky Mountain National Park (Baldwin and Bender, 2009). However, some of these studies are of limited local value because bear diet changes among habitats, years, and seasons.
Black bear habitat overlaps with people in parts of its distribution; bears are perceived as a threat to private property, crops and cattle. Private property damage by bears is common in rural areas; bears become accustomed to feeding from trash or food left outside households (McKinney and Delgadillo, 2004). Occasionally black bears destroy beehives, gardens and private infrastructure in search of food. In some cases, ranchers attract bears inadvertently by placing artificial feeders to supplement food for deer (mostly corn or other commercial foods). Deer feeders are an attraction for bears and many of them become accustomed to the feeders. Black bear predation on mule deer (Odocoileus hemionus), white tail deer (Odocoileus virginianus), elk (Cervus elaphus), moose (Alces alces), cattle (Bos taurus), and sheep (Ovis aries) has been documented in United States and Canada (Murie, 1948; Smith and Follmann, 1993; Singer et al., 1997). Furthermore, black bears are considered as significant predators of calves in areas such as Alaska (Franzmann et al., 1980), Idaho, Wyoming, Alberta, and Minnesota (Schlegel, 1976; Kunkel and Mech, 1994; Horstman and Gunson, 1982).
In Mexico, black bear habitat overlaps with extensive cattle ranching and farmlands, which are the main source of local income. When bears become a nuisance, they are trapped and eliminated to protect cattle and crops. However, the role of bear predation on cattle and local crops in Nuevo León, Mexico, is unknown. Knowledge of bear diet in local areas provides insight into local food items, habitat selection, if and when they feed on cattle or crops. Thus, this information will aid in the effective management of bears in the area.
Fecal analysis is a common method to study diets of large and elusive animals such as black bears (Bull and Wertz, 2001). Food items collected from scats are identified and related to diets through frequency, relative density, or volume (Hellgren, 1993). Our objective is to provide information on the seasonal importance of food items in the diet of black bears inhabiting the Sierra de Picachos, Nuevo León, Mexico.
Materials and methodsStudy area. The study area included a region in the south central area of the Sierra de Picachos (Fig. 1), located in the Sierra Madre Oriental, in the sub-province of the Sierras y Llanuras Coahuilenses, in the State of Nuevo León, Mexico. It has an area of 1 405km2, between 25°50'55” and 26°19'22” N, 99°42'57” al 100°06'40” W (Conabio, 2004). The mean annual temperature is 18°C (low annual mean temperature is 12°C, and the high annual mean temperature is 22°C). The annual rainfall is between 500cc -2 500cc (García, 1981). The topography is irregular and the altitude ranges from 400–1 200m asl (Detenal, 1978). The soil is predominantly litosol with secondary soils of rendzina and limestone regosol (www.inegi.org.mx). Sierra de Picachos includes 3 types of vegetation: scrub, pine, and oak forest (Ramos-Silva, 2004). There is poaching pressure on some wildlife species (e.g., black bear, mountain lion [Puma concolor], bobcat [Lynx rufus], and jaguarundi [Puma yagouaroundi]). The study site is on private land and it is considered a well-conserved sky island (Programa de manejo de la zona sujeta a conservacion estatal cerro Picachos, 2002).

Scat Collection. We collected 169 scats along roads and trails opportunistically during weekly hikes in the study area during December of 2004-November of 2005, which covered 3 seasons: dry season (January-July), rainy season (August-October), and post-rainy season (November- December). Each scat was placed in individually labeled paper bags. We assigned each scat a number and recorded date of collection and location (Universal Transverse Mercator coordinates). Scats were air dried to avoid mold from growing until they were processed.
Scat Analysis. We placed the scats individually in cloth bags and washed them by water displacement until clean. We dried the samples at room temperature and separated hair, insects, and plants manually to identify the diet components from each sample. For the identification of the plant material we created a reference collection of plants from the study area. We used electron microscope at the laboratory of plant development at the Facultad de Ciencias, UNAM.
We used patterns in the hair medulla and cortex to determine mammal species using reference catalogues (Arita, 1985; Baca-Ibarra y Sánchez-Cordero, 2004). We present the results as percent occurrence (PO) and importance index (PV). The PO was expressed as the number of times they appeared as a food component in the total number of feces collected (Martínez-Meyer, 1994; Hidalgo, 1998):
where: Fi is the total number of scats where food component i is present and N is the total number of scats.
For each element the importance index (PV) was quantified in cubic centimeters (cc) for each month and for each season. Volumes <0.2 cc were considered as a trace items. Importance index was calculated as (Niño-Ramírez, 1989):
where: V is the volume of the samples and Σ V is the sum of the volume of all samples.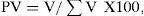
We also quantified the importance (I) of each food item as Craighead y Sumner (1982):
where: I=importance in percentage; Im=is given by: (100 PA/Σ PA)+(100 v/Σ v).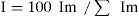
To estimate which season had the most trophic diversity (H'), we used the Shannon-Weaver index (Moreno, 2001):
where N is the number of all items in all samples and fi is the presence of food items i.
We used IBM SPSS Statistics 11 to do the Anova test to determine if the differences in the diversity of component among seasons were statistically significant using p<0.05 (Pardo and Ruiz, 2002).
ResultsScat analysis. Twenty different food items were detected, 73.4% plants, 16.8% animals (i.e., 16.1% invertebrates, 0.7% mammals), and debris (9.8%; mostly pulverized wood fragments and sand). We identified 12 items to the species level, 1 to genus, 4 to a family level, and we were unable to identify 2 items. The hair pattern of the medulla and cortex of the guard hairs found in the scats corresponded to white-tailed deer (Odocoileus virginianus) and armadillo (Dasypus novemcinctus).
Variation in the percentage of occurrence (PO) of food components. We identified 12 food items: 10 plants, and 2 animals during the dry season (January-July); 12 food items: 8 plants, and 4 animal species during the rainy season (August-October), and 10 different food items from the scats during the post-rainy season (November-December). However, the percent occurrence of the components identified in the scats did not vary between the different months and seasons (p=0.65).
The food items consumed in greatest proportion was the Texas Persimmon (Dyospyros texana; 19.7%), followed debris, small unidentified and undigested plant fragments and sand; (16.2%), acorns (Quercus spp.; 17.7%), thick leaf milk berry (Chiococca pachyphylla; 11.8%), prickly pear fruit (Opuntia spp.; 10.1%), fiber (undigested and unidentified plant pieces; 7.5%), sotol (Dasylirion texanum; 6.1%), and grapes (Vitis cinerea; 4.5%). Within the invertebrate items, mostly beetles (Coleoptera) were found (4.7%), ants, and bees were also found in a smaller percentage as well as other items (0.9%). We found the remains of an armadillo in one scat sample and white-tailed deer in another.
Variation in the percentage of volume (PV) of food components. The total plant material was 4 088cc, which accounted for 72.71% of total consumption. Animal matter was 64cc (1.14%), and debris accounted for 1 470 cc (26.15%). The 3 items with the largest PV value present were acorns (16.7%), fiber (14.76%), and Texas Persimmon (13.91%).
Importance of each food item. In general, oak acorns had the highest importance value (17.65%), followed by fiber (13.87%) and then Texas Persimmon (12.04%), other food items had low importance values (<7.0%).
Trophic diversity. The Shannon-Weaver diversity index (H ‘) was similar during the 3 seasons (dry=0.85, rainy=0.87 and post-rainy=0.88). The Anova test showed no statistically significant differences in the frequency analysis in food items consumed between seasons (p=0.65).
DiscussionThe analysis of the scats revealed that black bear diet in the Sierra de Picachos, Nuevo León, Mexico is mostly plants with a low percentage of non-cattle animal items, being insects. These results are consistent with previous studies in Sierra Madre Oriental (Delgadillo-Villalobos, 2001) and other studies in North America (Beeman and Pelton, 1980; Eagle and Pelton, 1983; Maehr and Brady, 1984). During this study, black bears presented a broad food spectrum with little variability in composition between seasons; however the percentage of different items changes between years and localities when results are compared with other studies in Mexico and other parts of North America.
It has been reported that the diet of black bears correlates with the fruiting season of each plant species. Since there is production of acorns all year round, we did not detect any variation on diet in relation with fruiting season. Seed production is highly variable and usually asynchronous, suggesting that reproductive strategies are different between the species of oak (Marroquín, 1997). The high consumption of acorns is probably due to the relative abundance in the region and to the highenergy content; acorns are also relatively high in fat and carbohydrates; additionally contain proteins, vitamins, calcium, and phosphorus (Landers et al., 1979; Eagle and Pelton, 1983).
Fiber consumption in our study site was high compared with other items, but lower than in other studies (Beeman and Pelton, 1980; Graber and White, 1981; Hellgren and Vaughan, 1988). This could be due to the fact that the fruits that are available are richer in water and sugar. Also, the consumption of debris seems to be mostly associated with the consumption of ants (Hellgren, 1993; Noyce et al., 1997). Invertebrates, consisting mainly of ants and beetles, were present in the diet of black bears over 9 months of the study. Their presence could be explained as a source of specific nutrients and amino acids that cannot be obtained from other dietary sources; they are also a rich source of nitrogen and lipids (Noyce et al., 1997). We found very few bees in the scats analyzed, it seems that in our study bees may be related to the consumption of honey; however, Landers et al. (1979), Maehr and Brady (1984) reported bees in stomach contents, but they reported higher amounts of bees than this study.
The consumption of prickly pear (Opuntia spp.) was about 50%, higher than the percentages found by Hellgren (1993) in the Big Bend area in Texas (7 to 17%) and DoanCrider (1995) that reported a similar percentage fruit density in scats analyzed in the foothills of the Burro, Coahuila. Notably, thick leaf milkberry (Chiococca pachyphylla) is a species frequently consumed and important for black bear in the Sierra de Picachos; however, this plant species has not been reported in black bear diet for other populations. We found a small percentage of Laredo Oregon-grape (Mahomia trifoliata) intake, although there was a high density of fruiting in our study area. This contrasts with a report by Hellgren (1993) in Big Bend National Park, where this fruit was one of the dominant food components in the diet of black bears between the months of May-June.
Bears in the area eat the fruit of Persea pachypoda but we found no support for bears consuming the leaves. In contrast, Landers et al (1979) found that black bears from southeastern North Carolina eat only leaves and no fruits of Persea borbonia. Hellgren (1993) reported bears eating sotol (Dasylirion wheeleri) in the study area in small amounts, similar results were presented by Silva-Hurtado (2004). The Texas persimmon (Dyospyros texana) was one of the most common fruits consumed by bears, the equivalent of which occurs in the Big Bend National Park during the months of September and October (DoanCrider, 1995).
For the Sierra de Picachos it has been reported that black bears have killed foals, goats, and sheep (Jiménez-Guzmán et al., 1999); there were also reports of bears eating carrion. Mammals represent a small portion of bear diet; this study, as well as other scat studies in the area has shown a low presence (16.8%) of vertebrate remains and an even lesser presence of cattle and goat remains (Niño-Ramírez, 1989). We found only small amount of deer and no cattle remains in the diet; therefore we cannot confirm previous reports.
In black bear diet, plants were the most common item, animals and mainly insects were the next most common items, and undetermined wood fragments were least common. There was no statistically significant difference among seasons. We found only small amounts of deer remains, but it is unknown if these were taken through predation or consumed as carrion. Our analysis also failed to find cattle residue in any of the scats; therefore, black bears in this study site either do not, or rarely feed upon livestock. Cattle predation should be managed by personnel trained to differentiate predation by bear, cougar, coyote and bobcat (Lynx rufus). The misidentification of livestock deaths due to black bears, usually results in unnecessary trapping and in many cases the elimination of the species.
Another concern is presence of corn in black bear diet, which warrants further monitoring given the propensity of bears to eat crops such as corn and oats. Bears break and smash corn stalks in the process of consuming the entire cob and they can damage large areas of cropland. In the Sierra de Picachos, cornfields are a significant source of income and food for local communities. Fencing has proven to be effective in deterring bears from eating crops, and if possible energizing fences with 3 000 volts should keep bears away. However these fences are expensive and can be cost-prohibitive. Ranchers often face the choice of comparing the cost of bears eating corn vs. the cost of implementing preventive measures, such as fencing or hunting. Therefore, these considerations and tradeoffs need to be understood by wildlife managers when developing prevention programs that diminish the hunting of bears in order to protect crops.
Black bears are currently listed as an “endangered species” and it is imperative to assure their long-term survival. Management actions, public involvement and outreach need to be implemented to foster the understanding and participation of local communities in the efforts to protect this endangered species. Bears live where there is shelter, water and food, in some cases these requirements overlap with ranches, farms and even towns. Bear-human conflict is not a new problem, but it is essential to remediate the conflict for the sake of bear survival. We have to find a way for bears to coexist with people. Government officials could implement an extensive and targeted outreach campaign (flyers/signs and/or workshops) for local communities with information regarding how to deal with bear presence on their lands. Specific actions can prevent damage caused by bears, such as not feeding bears, not approaching or harassing bears, keeping garbage in containers with locking lids, storing livestock feed and pet food in closed containers or inside buildings with closed doors, erecting fences around gardens and crops, among others. Also, there is a need for more governmental action to protect bear habitat and deter bear presence in human settlements such as the removal of invasive woody species, implementing water conservation and erosion control activities. Community-based actions combined with governmental actions would improve black bear habitat and should help maintain the availability of food, water and cover for bears and minimize bear-human conflicts.
This paper constitutes a partial fulfillment of the Graduate Program in Biological Sciences of UNAM. L. A. Juárez-Casillas acknowledges the scholarship and financial support provided by the Conacyt and UNAM. We thank the specialists from the National Herbarium (MEXU), UNAM, and the Herbarium (UANL), especially to Martha Virginia Olvera-García. We would like to thank Andrés M. Sada for allowing us to collect samples on his ranch; the researchers from the Organización Vida Silvestre, A. C., Marivel Medrano-Padilla and Cecilia Hernández-Cavazos for their help in the sample collection; Marcela González-Álvarez at the UANL Herbarium for helping in the identification of seeds.