



Three individuals of Multicalyx cristata (Aspidocotylea) were identified in the gallbladder of a hammerhead shark, Sphyrna lewini (Elasmobranchii), in Playa Chachalacas, Veracruz, Mexico. This report makes a significant contribution to our understanding of the biogeography of the Aspidocotylea species that parasitize sharks, expanding their documented range into the Mexican waters of the Gulf of Mexico and in the Atlantic Ocean.
Se determinaron 3 individuos de Multicalyx cristata (Aspidocotylea) de la vesícula biliar de un tiburón martillo Sphyrna lewini (Elasmobranchii) en playa Chachalacas, Veracruz, México. Este registro es una contribución significativa en términos de la biogeografía de las especies de Aspidocotylea que parasitan tiburones, incrementando su distribución en aguas mexicanas del golfo de México y océano Atlántico.
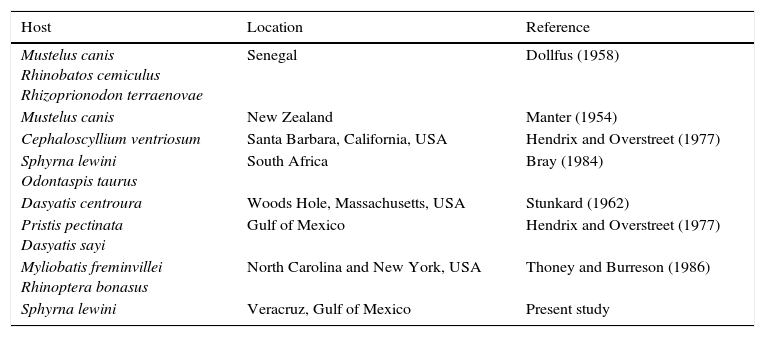
Multicalyx cristata (Faust & Tang, 1936) (Fig. 1) was originally described in the spiral valve (intestine) of a cownose ray, Rhinoptera bonasus (Mitchill, 1815), near Biloxi Bay, Mississippi, USA. Since then, M. cristata has been reported in the gallbladders of various elasmobranch species (Bray, 1984; Dollfus, 1958; Hendrix & Overstreet, 1977; Stunkard, 1962; Thoney & Burreson, 1987) in both the Atlantic and the Pacific Oceans (Table 1). Although the species has been reported in the spiral valves of some hosts (R. bonasus and Sphyrna lewini), M. cristata typically target the gallbladders of adult hosts (Dollfus, 1958; Stunkard, 1962) and the bile ducts of juvenile hosts (Hendrix & Overstreet, 1977). Manter (1954) and Hendrix and Overstreet (1977) reported juvenile M. cristata in Menticirrhus americanus (Linnaeus, 1758) (Sciaenidae) and Sphoeroides testudineus (Linnaeus, 1758) (Tetraodontidae), indicating that teleost fishes can serve as paratenic or transport hosts; this also may be a route of infection for neritic predators, like sharks. The intermediate hosts of M. cristata are currently unknown; however, Gibson and Chinabut (1984) mention that the order (Stichocotylida) to which M. cristata belongs can use crustaceans as intermediate hosts.
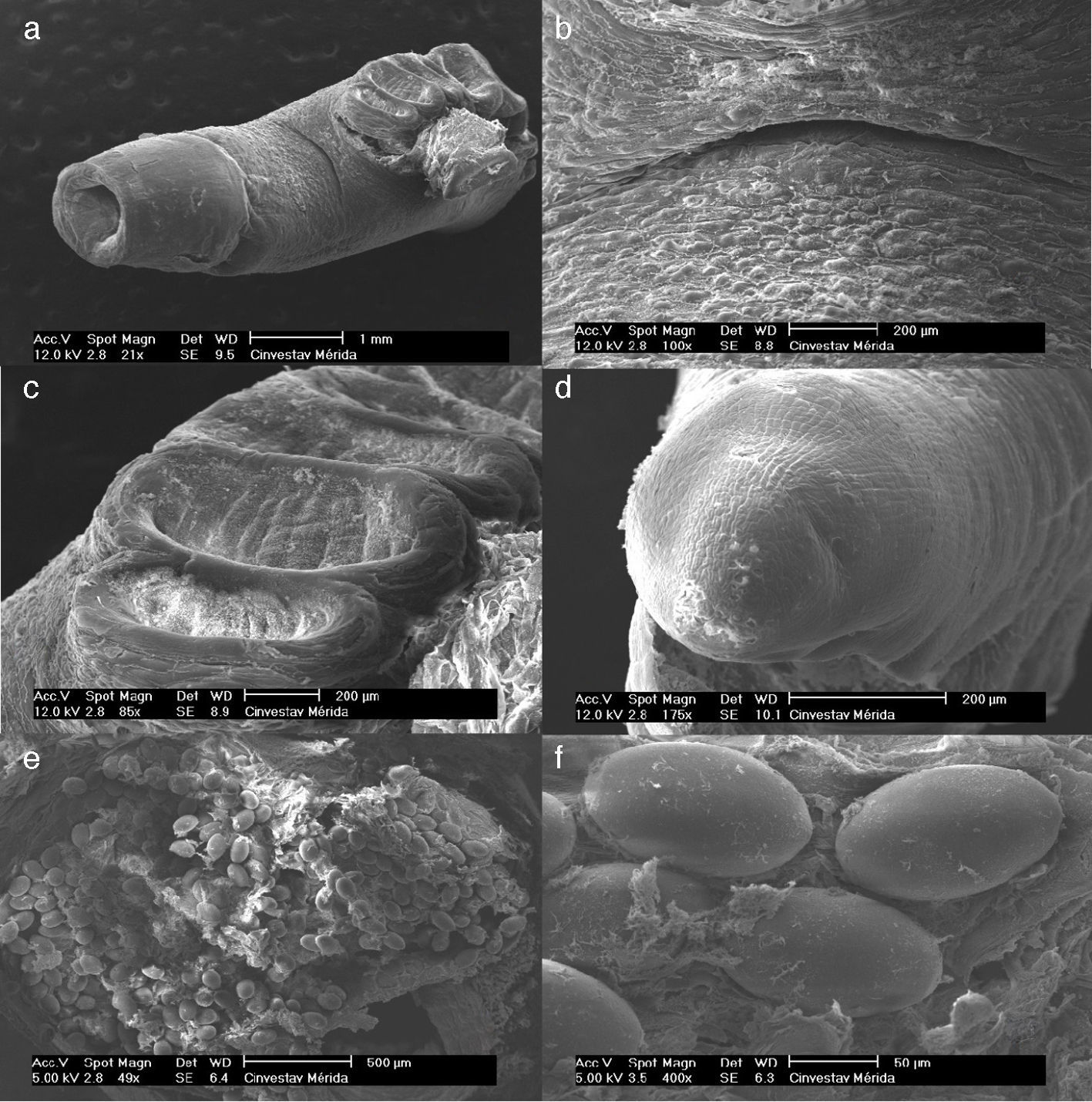
 complete individual, the arrow shows the anterior portion (10×); (b) anterior view showing the mouth, genital pore (arrow), and alveoli (top) (40×); (c) side view showing the transverse ridges of the alveoli (40×); (d) posterior view showing the position of the excretory papilla (arrow) (40×); (e) mid-section showing part of the uterus with eggs and the vitellogen (arrow) in the lateral position (40×, and (f) embryonated eggs (40×). Images (a)–(e) were captured with a Motic SMZ-168 stereoscopic microscope, (f) was taken with an Olympus BX50 optical microscope.")
Morphology of Multicalyx cristata: (a) complete individual, the arrow shows the anterior portion (10×); (b) anterior view showing the mouth, genital pore (arrow), and alveoli (top) (40×); (c) side view showing the transverse ridges of the alveoli (40×); (d) posterior view showing the position of the excretory papilla (arrow) (40×); (e) mid-section showing part of the uterus with eggs and the vitellogen (arrow) in the lateral position (40×, and (f) embryonated eggs (40×). Images (a)–(e) were captured with a Motic SMZ-168 stereoscopic microscope, (f) was taken with an Olympus BX50 optical microscope.
Reports of Multicalyx cristata in elasmobranch species.
| Host | Location | Reference |
|---|---|---|
| Mustelus canis Rhinobatos cemiculus Rhizoprionodon terraenovae | Senegal | Dollfus (1958) |
| Mustelus canis | New Zealand | Manter (1954) |
| Cephaloscyllium ventriosum | Santa Barbara, California, USA | Hendrix and Overstreet (1977) |
| Sphyrna lewini Odontaspis taurus | South Africa | Bray (1984) |
| Dasyatis centroura | Woods Hole, Massachusetts, USA | Stunkard (1962) |
| Pristis pectinata Dasyatis sayi | Gulf of Mexico | Hendrix and Overstreet (1977) |
| Myliobatis freminvillei Rhinoptera bonasus | North Carolina and New York, USA | Thoney and Burreson (1986) |
| Sphyrna lewini | Veracruz, Gulf of Mexico | Present study |
Thoney and Burreson (1986) examined 16 species of sharks collected between Cape Fear, North Carolina, and Long Island, New York, USA, identifying specimens of M. cristata exclusively in Myliobatis freminvillei (Lesueur, 1824) (Bullnose eagle ray) and Rhinoptera bonasus (Mitchill, 1815). According to the authors, the absence of M. cristata in the elasmobranchs they examined was due to the small sample size. However, no M. cristata were reported in 42 individuals of the shark Mustelus canis (Mitchill, 1815) (Triakidae).
We report the presence of 3 individuals of M. cristata in the gallbladder of a male hammerhead shark, S. lewini (Griffith & Smith, 1834), caught by the artisanal fishery in Chachalacas, Veracruz, Mexico. The specimens were placed in a bottle with 7% saline solution, and cooled in ice to encourage their relaxation and to be able to separate them. Subsequently, they were fixed in warm formalin at 4%. In the laboratory the samples were transferred to 70% ethanol. Due to the large size of the specimens, it was not possible to prepare permanent slides; however, 1 individual was prepared for scanning electron microscopy (SEM). For SEM, the specimen was washed for 30min with 0.1M sodium cacodylate buffer, and post-fixed with 1% osmium tetroxide for 30min. Subsequently the sample was washed again for 30min in sodium cacodylate and then dehydrated in ethanol gradients from 30% to 100% for 10min at each step; finally it was dried in a critical point dryer. The sample was subsequently recovered with a film of typically one evaporator in high vacuum (sputter/coater Polaron SC7640). Micrographs were obtained using a Philips XL-30 environmental SEM. One specimen was deposited in the Colección Nacional de Helmintos (CNHE) at the Instituto de Biología, UNAM, as reference number CNHE-9994.
The 3 specimens were identified as M. cristata because their characteristics match those described by Faust and Tang (1936). The recovered specimens were adults with abundant eggs in the uterus. The bodies were elongated and cylindrical, reaching 202–223mm (211.6±10.5) total length (TL). Despite their large size, these organisms did not obstruct the gallbladder as there were only 3 individuals. Thoney and Burreson (1988) mention that mature M. cristata can reach 60cm TL. The individuals reported by Hendrix and Overstreet (1977) were juveniles measuring 28–101mm TL. In contrast, Multicalyx elegans (Olsson, 1869) reach sizes of 23mm and parasitizes the gallbladders of holocephals. M. cristata can occur individually or in groups of 2 or more organisms; however, there are records of a single individual with developing eggs within the uterus suggesting that self-fertilization may occur (Thoney & Burreson, 1986). Intraspecific competition may occur due to the large size attained by M. cristata in the gallbladder. Further research is required to confirm this; however, authors like Thoney and Burreson (1988) suggest that such competition would limit the intensity of infection to 1 or 2 individuals.
The ventral haptor extends almost the entire length of the body, consisting of a single row of 1668 alveoli subdivided by the transverse septum (Figs. 1a–d and 2a, c). This is similar to Bray's (1984) report of 1500 alveoli in a specimen of M. cristata obtained from a hammerhead shark, S. lewini, on the shores of South Africa. Our M. cristata specimen has a terminal mouth in the form of funnel; a ventral genital pore anterior to the haptor (Figs. 1b and 2a, b); oval, operculated eggs measuring 0.12×0.08μm (Figs. 1e, f and 2e, f); and vitellaria formed by 2 bands of lateral (Fig. 1f).
 anterior view showing the mouth funnel, genital pore, and anterior alveoli; (b) genital pore; (c) alveoli; (d) excretory papilla; (e) uterus and eggs; and (f) embryonated eggs.")
The excretory papilla is located on the posterior end of the body (Figs. 1d and 2d).
This is the first report of M. cristata in S. lewini in Mexican waters and for the coast of Veracruz. Further research is necessary as we currently know very little about the life cycle of these organisms. Such studies should include a parasitological examination of the stomach contents of other elasmobranch species, particularly those that have already been identified as hosts (Table 1), as well as a pathology assessment of the damage caused to the gallbladder. Once this information has been collected, it will be possible to identify a potential intermediate host; it is unlikely that a single intermediate mollusk host occurs in such a broad range of diverse habitats.
Our report of Multicalyx cristata off the Veracruz coast is an important contribution to our understanding of the biogeography of the Aspidocotylea species that parasitize sharks. The presence of this parasite in the hammerhead shark S. lewini in the Mexican waters of the Gulf of Mexico expands its range in the Atlantic Ocean.
The authors thank the fishermen of Chachalacas for supplying the hammerhead shark. Thanks to José Luis Recio Silva and Miguel Ángel Dorantes González for their assistance during field work and to Ana Ruth Cristobal for capturing the electronic microscopy images. We thank Consejo Nacional de Ciencia y Tecnología for the scholarship program for post-doctoral financial support. Sample analysis was carried out at the Laboratory of Aquatic Pathology at the Centro de Investigación y de Estudios Avanzados, Unidad Mérida, Instituto Nacional Politécnico.
Peer Review under the responsibility of Universidad Nacional Autónoma de México.