




Se describe una especie nueva de tiburón perteneciente a la familia Ginglymostomatidae; la distribución de esta especie es amplia en el Pacífico oriental tropical (POT), donde es endémica. Ginglymostoma unami sp. nov. se describe a partir de ejemplares procedentes del POT identificados bajo el nombre de Ginglymostoma cirratum, especie que se consideraba con distribución anfiamericana; esta última ahora se restringe al océano Atlántico. Ginglymostoma unami sp. nov. se distingue de G. cirratum por una serie de caracteres morfométricos, tales como la distancia entre la región prebranquial y la interdorsal, aquella entre el extremo posterior de la segunda aleta dorsal y el inicio del lóbulo caudal superior, que son más cortas y, por último, por la distancia entre la axila y el inicio de la aleta anal; también difiere por la posición de la inserción de la primer aleta dorsal con respecto a aquella de las pélvicas y en la forma y número de quillas en los dentículos dérmicos y morfología dental.
A new species of shark belonging to the family Ginglymostomatidae is herein described; the new species show a wide distribution in the Tropical Eastern Pacific, where it is endemic. Ginglymostoma unami sp. nov. was previously recognized as G. cirratum exhibiting an amphi-American distribution; however that species is now considered to be restricted to the Atlantic Ocean. Ginglymostoma unami sp. nov. can be readily distinguished from G. cirratum by comparing several meristic characters such as the distance between the prebanchial and interdorsal regions, and that between posterior end of the second dorsal fin and the beginning of the caudal lobe, both being shorter; the new species also differs by the position of the insertion of the first dorsal fin with regard to the pelvic fins and in the form and number of keels on the dermal denticles and teeth morphology.
La diversidad de condrictios que existe a nivel mundial incluye aproximadamente 1227 especies (Eschmeyer y Fong, 2014), riqueza que se ha incrementado en los últimos años con el descubrimiento de nuevas especies en diferentes localidades. Ejemplo de ello, el reciente descubrimiento del tiburón Carcharhinus humani en el océano Índico (White y Weigmann, 2014), la raya dulceacuícola Potamotrygon limai descrita para la cuenca del Amazonas, Brasil (Fontenelle, Da Silva y De Carvalho, 2014) y la raya de profundidad Rajella paucispinosa descrita en el Índico occidental (Weigmann, Stehmann y Thiel, 2014). Dentro de los condrictios, la familia Ginglymostomatidae (Orectolobiformes: Elasmobranchii) está representada actualmente por 3 géneros, todos ellos monotípicos (sensuCompagno, 2002, 2005; Eschmeyer, 2014): Nebrius Rüppell, 1837, con la especie Nebrius ferrugineus (Lesson, 1831), ampliamente distribuida en los océanos Índico y Pacífico occidental y central; Pseudoginglymostoma Dingerkus, 1986, con Pseudoginglymostoma brevicaudatum (Günther, 1867), restringida al océano Índico occidental; y Ginglymostoma Müller y Henle, 1837, representado hasta este momento por la especie G. cirratum (Bonnaterre, 1788), la cual incluye al menos 3 poblaciones disyuntivas (Pacífico oriental, Atlántico occidental y oriental), cuya diferenciación morfológica aún no ha sido estudiada (Compagno, 2002).
En los últimos años se han realizado trabajos taxonómicos donde se implementan herramientas de análisis genéticos que complementan los datos morfológicos y de distribución geográfica. En este contexto, las especies de peces con distribución disyunta en las costas del continente Americano, es decir, las conocidas como anfiamericanas, han recibido notable atención (Banford y Collette, 2001; Burhanuddin y Parin, 2008; Craig et al., 2009; Pfeiler, Van der Heiden, Ruboyianes y Watts, 2011). Durante la realización del catálogo actualizado de los condrictios que habitan en las costas de México (Del Moral-Flores y Pérez-Ponce de León, 2013), se evaluó el estatus taxonómico del tiburón gata G. cirratum, en correspondencia a la observación de Compagno (2002), a partir de lo cual se reconocieron diferencias morfológicas entre los ejemplares procedentes de las costas del Atlántico occidental y aquellos del POT; estas diferencias permiten sustentar los resultados de Karl, Castro y Garla (2012), quienes previamente analizaron la región mitocondrial control y un total de 8 microsatélites en individuos de diferentes poblaciones del Atlántico y del Pacífico para evaluar la variación genética, encontrando que la población del Pacífico está muy diferenciada de aquellas del Atlántico y sugiriendo que podría representar una especie críptica. El estudio detallado de la morfología de ejemplares recientemente colectados, en conjunto con la observación de ejemplares depositados en colecciones biológicas, reveló la existencia de una serie de caracteres morfológicos que permiten separar a la especie del Pacífico de aquella del Atlántico, sugerida como especie críptica. En este trabajo se describe esta nueva especie.
Material y métodosLa evaluación comparativa de la taxonomía de G. cirratum (sensu lato), se basó en la diagnosis original de la especie, sus sinónimos y modificaciones taxonómicas (por ejemplo, Bancroft, 1830; Bloch y Schneider, 1801; Bonnaterre, 1788; De Brito-Capello, 1867; Griffith y Smith, 1834; Lacepède, 1800; Poey, 1861), a partir de lo cual se corroboró la nueva entidad taxonómica cuya distribución comprende el POT. Asimismo, se revisó material depositado en las siguientes colecciones de referencia: Colección Nacional de Peces, Instituto de Biología de la Universidad Nacional Autónoma de México (CNPE-IBUNAM); Colección de Peces de la Facultad de Estudios Superiores Iztacala, UNAM (CPFESI); Colección Ictiológica del Instituto de Ciencias del Mar y Limnología de la UNAM (CI-ICMYL); Colección Ictiológica del Centro Interdisciplinario de Ciencias Marinas (CI-CICIMAR) del Instituto Politécnico Nacional (IPN); Colección de Peces del Laboratorio de Ecología Marina de la Escuela Nacional de Ciencias Biológicas del IPN (ENCB-IPN-LEM); Colección de Material Reciente del Museo de Geología del Instituto de Geología de la UNAM (IGM-CMR); Colección de Peces del Laboratorio de Ictiología y Biología Pesquera de la Universidad del Mar, campus Puerto Ángel, Oaxaca (CP-UMAR); Colección de Peces de la Universidad Autónoma de Yucatán (UADY); Museo de Zoología, Escuela de Biología, Universidad de Costa Rica (UCR). Además, se recolectaron 2 ejemplares que se integraron en la serie tipo: uno en Puerto Ángel, Oaxaca, México, el cual se trasladó a la Facultad de Estudios Superiores Zaragoza, UNAM, donde se obtuvieron las biometrías correspondientes y se tomaron las fotografías para registrar detalles de la coloración, el ejemplar fue depositado en la CNPE-IBUNAM (Núm. de catálogo: CNPE-IBUNAM 18850); el segundo espécimen fue capturado en la costa Pacífica de Costa Rica, se trasladó al UCR donde fue depositado (UCR 3011-01), previamente se obtuvieron las medidas y fotografías correspondientes. De ambos organismos se resguardan muestras de tejido en las respectivas colecciones.
Los datos morfométricos de los ejemplares se obtuvieron siguiendo la terminología anatómica establecida por Compagno (1984, 2002) y Goto (2001), mientras que la fórmula y morfología dental se describió bajo el esquema de Applegate (1965). Se tomaron 2 muestras de piel de uno de los ejemplares (CNPE-IBUNAM 18850) para analizar la morfología de los dentículos dérmicos, mediante el empleo de un microscopio electrónico de barrido (Hitachi SU 1510 a 10 kv) siguiendo el método descrito en Mendoza-Garfias y Pérez-Ponce de León (2005).
DescripciónGinglymostoma unami sp. nov.
Ginglymostoma sp. (Castro, 2011a: 191) (figs. 1-5; tabla 1)
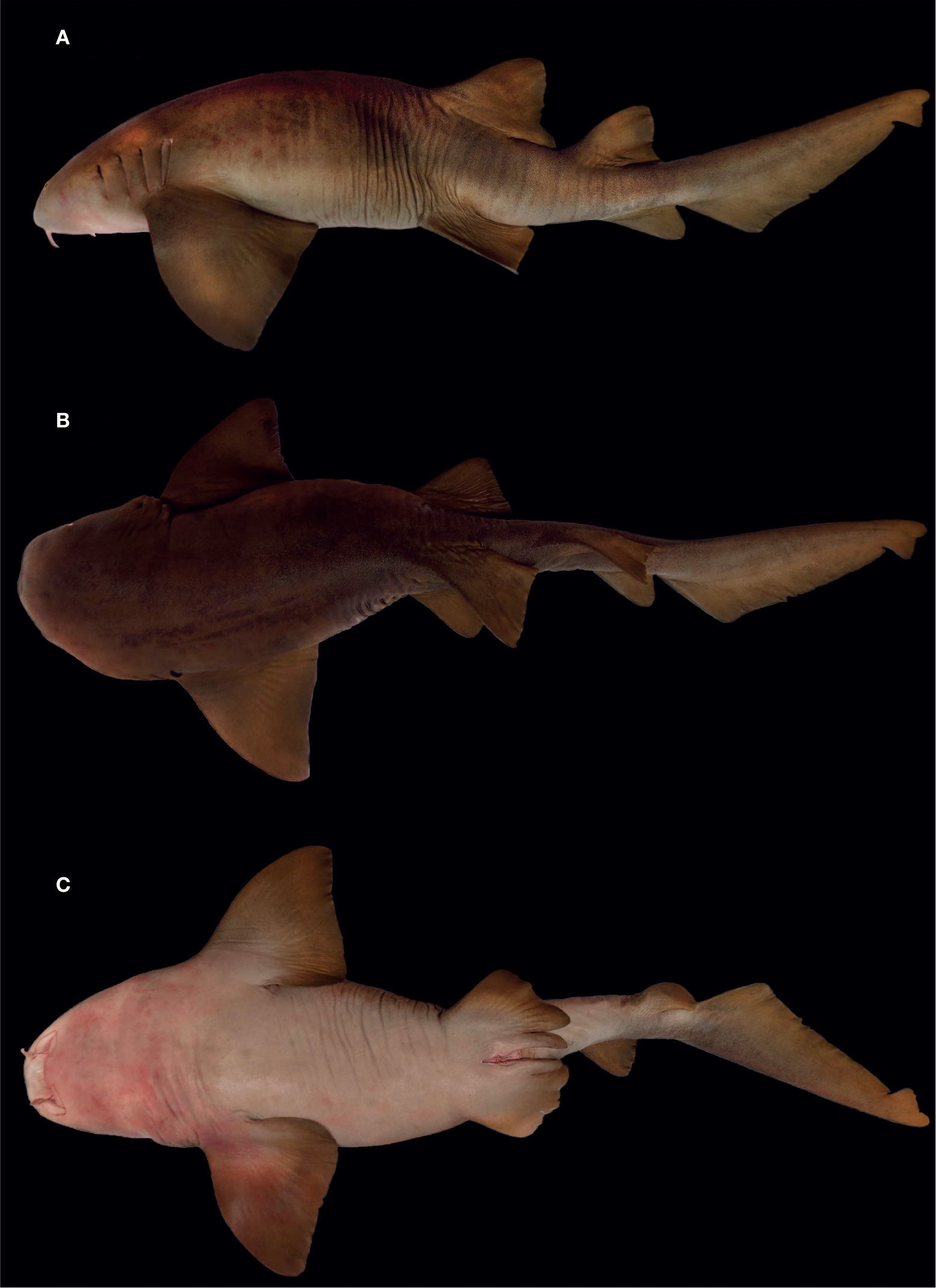
: A, vista lateral; B, vista dorsal; C, vista ventral.")
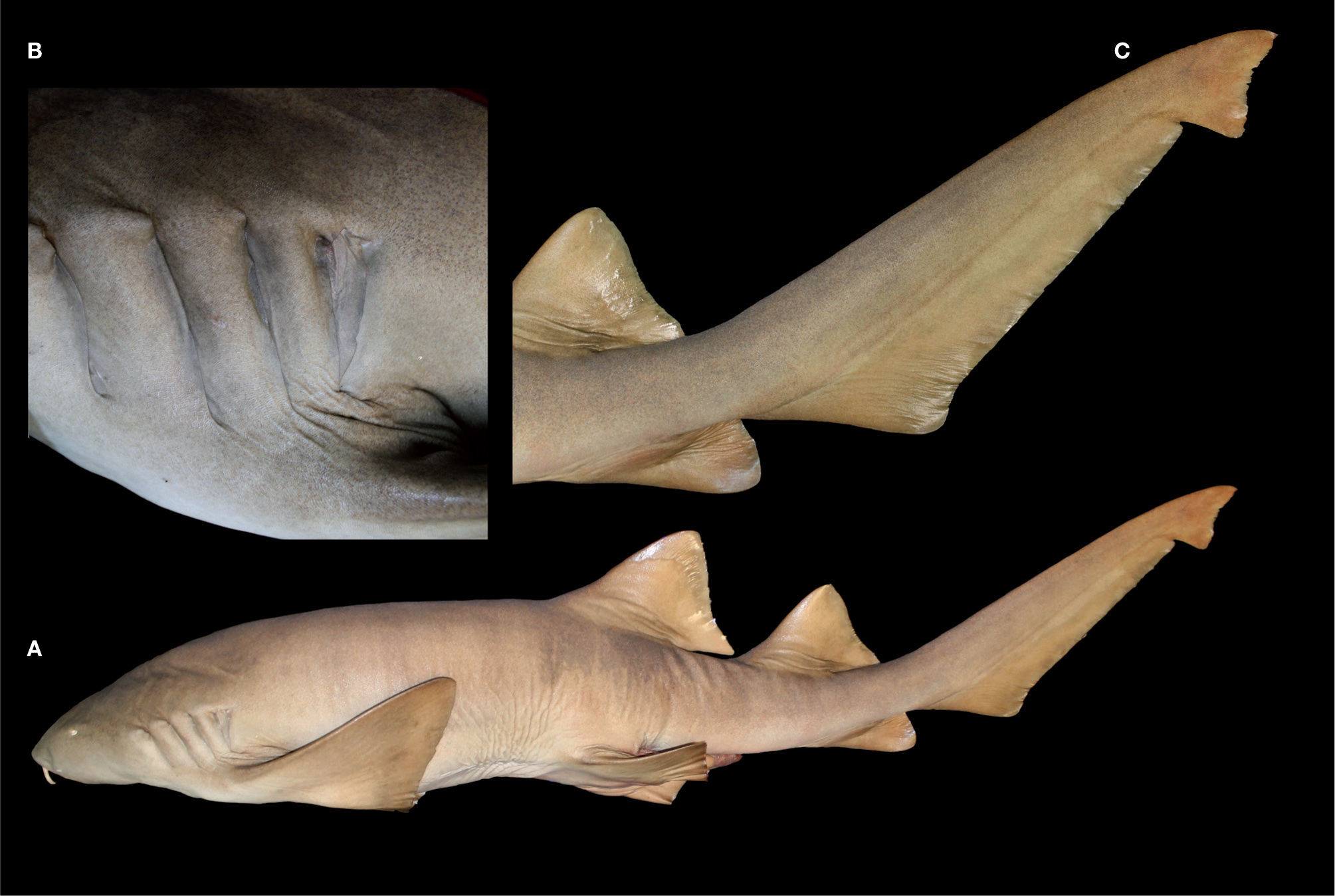
: A, vista lateral; B, acercamiento región branquial; C, acercamiento región caudal.")
: A, vista lateral; B, vista dorsal; C, vista ventral.")


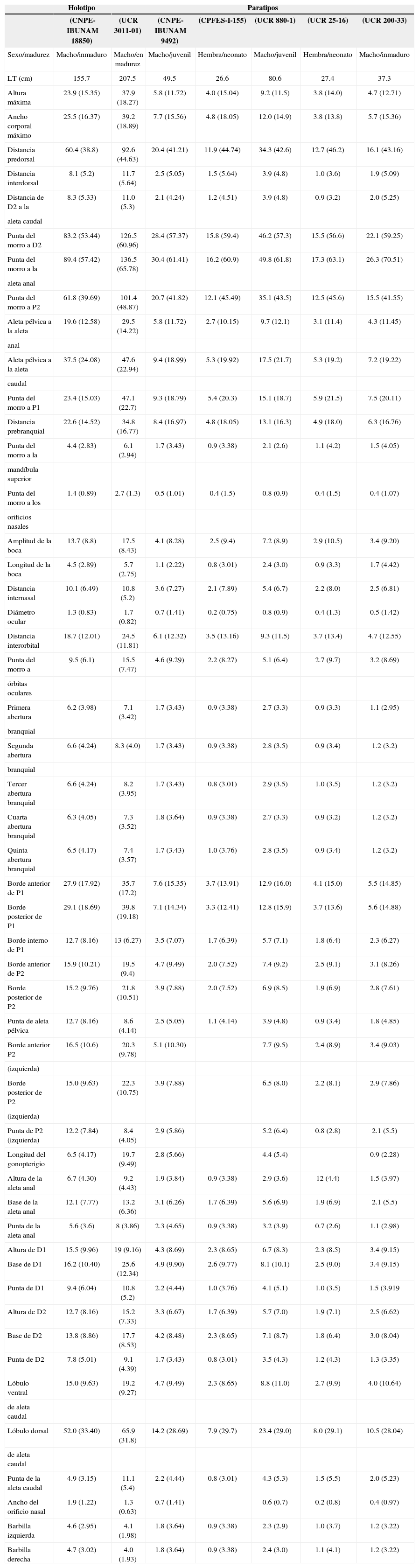
Datos morfométricos de Ginglymostoma unami sp. nov.; entre paréntesis se exponen las proporciones expresadas como porcentaje de la longitud total (LT). Las medias están en cm
| Holotipo | Paratipos | ||||||
|---|---|---|---|---|---|---|---|
| (CNPE-IBUNAM 18850) | (UCR 3011-01) | (CNPE-IBUNAM 9492) | (CPFES-I-155) | (UCR 880-1) | (UCR 25-16) | (UCR 200-33) | |
| Sexo/madurez | Macho/inmaduro | Macho/en madurez | Macho/juvenil | Hembra/neonato | Macho/juvenil | Hembra/neonato | Macho/inmaduro |
| LT (cm) | 155.7 | 207.5 | 49.5 | 26.6 | 80.6 | 27.4 | 37.3 |
| Altura máxima | 23.9 (15.35) | 37.9 (18.27) | 5.8 (11.72) | 4.0 (15.04) | 9.2 (11.5) | 3.8 (14.0) | 4.7 (12.71) |
| Ancho corporal máximo | 25.5 (16.37) | 39.2 (18.89) | 7.7 (15.56) | 4.8 (18.05) | 12.0 (14.9) | 3.8 (13.8) | 5.7 (15.36) |
| Distancia predorsal | 60.4 (38.8) | 92.6 (44.63) | 20.4 (41.21) | 11.9 (44.74) | 34.3 (42.6) | 12.7 (46.2) | 16.1 (43.16) |
| Distancia interdorsal | 8.1 (5.2) | 11.7 (5.64) | 2.5 (5.05) | 1.5 (5.64) | 3.9 (4.8) | 1.0 (3.6) | 1.9 (5.09) |
| Distancia de D2 a la | 8.3 (5.33) | 11.0 (5.3) | 2.1 (4.24) | 1.2 (4.51) | 3.9 (4.8) | 0.9 (3.2) | 2.0 (5.25) |
| aleta caudal | |||||||
| Punta del morro a D2 | 83.2 (53.44) | 126.5 (60.96) | 28.4 (57.37) | 15.8 (59.4) | 46.2 (57.3) | 15.5 (56.6) | 22.1 (59.25) |
| Punta del morro a la | 89.4 (57.42) | 136.5 (65.78) | 30.4 (61.41) | 16.2 (60.9) | 49.8 (61.8) | 17.3 (63.1) | 26.3 (70.51) |
| aleta anal | |||||||
| Punta del morro a P2 | 61.8 (39.69) | 101.4 (48.87) | 20.7 (41.82) | 12.1 (45.49) | 35.1 (43.5) | 12.5 (45.6) | 15.5 (41.55) |
| Aleta pélvica a la aleta | 19.6 (12.58) | 29.5 (14.22) | 5.8 (11.72) | 2.7 (10.15) | 9.7 (12.1) | 3.1 (11.4) | 4.3 (11.45) |
| anal | |||||||
| Aleta pélvica a la aleta | 37.5 (24.08) | 47.6 (22.94) | 9.4 (18.99) | 5.3 (19.92) | 17.5 (21.7) | 5.3 (19.2) | 7.2 (19.22) |
| caudal | |||||||
| Punta del morro a P1 | 23.4 (15.03) | 47.1 (22.7) | 9.3 (18.79) | 5.4 (20.3) | 15.1 (18.7) | 5.9 (21.5) | 7.5 (20.11) |
| Distancia prebranquial | 22.6 (14.52) | 34.8 (16.77) | 8.4 (16.97) | 4.8 (18.05) | 13.1 (16.3) | 4.9 (18.0) | 6.3 (16.76) |
| Punta del morro a la | 4.4 (2.83) | 6.1 (2.94) | 1.7 (3.43) | 0.9 (3.38) | 2.1 (2.6) | 1.1 (4.2) | 1.5 (4.05) |
| mandíbula superior | |||||||
| Punta del morro a los | 1.4 (0.89) | 2.7 (1.3) | 0.5 (1.01) | 0.4 (1.5) | 0.8 (0.9) | 0.4 (1.5) | 0.4 (1.07) |
| orificios nasales | |||||||
| Amplitud de la boca | 13.7 (8.8) | 17.5 (8.43) | 4.1 (8.28) | 2.5 (9.4) | 7.2 (8.9) | 2.9 (10.5) | 3.4 (9.20) |
| Longitud de la boca | 4.5 (2.89) | 5.7 (2.75) | 1.1 (2.22) | 0.8 (3.01) | 2.4 (3.0) | 0.9 (3.3) | 1.7 (4.42) |
| Distancia internasal | 10.1 (6.49) | 10.8 (5.2) | 3.6 (7.27) | 2.1 (7.89) | 5.4 (6.7) | 2.2 (8.0) | 2.5 (6.81) |
| Diámetro ocular | 1.3 (0.83) | 1.7 (0.82) | 0.7 (1.41) | 0.2 (0.75) | 0.8 (0.9) | 0.4 (1.3) | 0.5 (1.42) |
| Distancia interorbital | 18.7 (12.01) | 24.5 (11.81) | 6.1 (12.32) | 3.5 (13.16) | 9.3 (11.5) | 3.7 (13.4) | 4.7 (12.55) |
| Punta del morro a | 9.5 (6.1) | 15.5 (7.47) | 4.6 (9.29) | 2.2 (8.27) | 5.1 (6.4) | 2.7 (9.7) | 3.2 (8.69) |
| órbitas oculares | |||||||
| Primera abertura | 6.2 (3.98) | 7.1 (3.42) | 1.7 (3.43) | 0.9 (3.38) | 2.7 (3.3) | 0.9 (3.3) | 1.1 (2.95) |
| branquial | |||||||
| Segunda abertura | 6.6 (4.24) | 8.3 (4.0) | 1.7 (3.43) | 0.9 (3.38) | 2.8 (3.5) | 0.9 (3.4) | 1.2 (3.2) |
| branquial | |||||||
| Tercer abertura branquial | 6.6 (4.24) | 8.2 (3.95) | 1.7 (3.43) | 0.8 (3.01) | 2.9 (3.5) | 1.0 (3.5) | 1.2 (3.2) |
| Cuarta abertura branquial | 6.3 (4.05) | 7.3 (3.52) | 1.8 (3.64) | 0.9 (3.38) | 2.7 (3.3) | 0.9 (3.2) | 1.2 (3.2) |
| Quinta abertura branquial | 6.5 (4.17) | 7.4 (3.57) | 1.7 (3.43) | 1.0 (3.76) | 2.8 (3.5) | 0.9 (3.4) | 1.2 (3.2) |
| Borde anterior de P1 | 27.9 (17.92) | 35.7 (17.2) | 7.6 (15.35) | 3.7 (13.91) | 12.9 (16.0) | 4.1 (15.0) | 5.5 (14.85) |
| Borde posterior de P1 | 29.1 (18.69) | 39.8 (19.18) | 7.1 (14.34) | 3.3 (12.41) | 12.8 (15.9) | 3.7 (13.6) | 5.6 (14.88) |
| Borde interno de P1 | 12.7 (8.16) | 13 (6.27) | 3.5 (7.07) | 1.7 (6.39) | 5.7 (7.1) | 1.8 (6.4) | 2.3 (6.27) |
| Borde anterior de P2 | 15.9 (10.21) | 19.5 (9.4) | 4.7 (9.49) | 2.0 (7.52) | 7.4 (9.2) | 2.5 (9.1) | 3.1 (8.26) |
| Borde posterior de P2 | 15.2 (9.76) | 21.8 (10.51) | 3.9 (7.88) | 2.0 (7.52) | 6.9 (8.5) | 1.9 (6.9) | 2.8 (7.61) |
| Punta de aleta pélvica | 12.7 (8.16) | 8.6 (4.14) | 2.5 (5.05) | 1.1 (4.14) | 3.9 (4.8) | 0.9 (3.4) | 1.8 (4.85) |
| Borde anterior P2 | 16.5 (10.6) | 20.3 (9.78) | 5.1 (10.30) | 7.7 (9.5) | 2.4 (8.9) | 3.4 (9.03) | |
| (izquierda) | |||||||
| Borde posterior de P2 | 15.0 (9.63) | 22.3 (10.75) | 3.9 (7.88) | 6.5 (8.0) | 2.2 (8.1) | 2.9 (7.86) | |
| (izquierda) | |||||||
| Punta de P2 (izquierda) | 12.2 (7.84) | 8.4 (4.05) | 2.9 (5.86) | 5.2 (6.4) | 0.8 (2.8) | 2.1 (5.5) | |
| Longitud del gonopterigio | 6.5 (4.17) | 19.7 (9.49) | 2.8 (5.66) | 4.4 (5.4) | 0.9 (2.28) | ||
| Altura de la aleta anal | 6.7 (4.30) | 9.2 (4.43) | 1.9 (3.84) | 0.9 (3.38) | 2.9 (3.6) | 12 (4.4) | 1.5 (3.97) |
| Base de la aleta anal | 12.1 (7.77) | 13.2 (6.36) | 3.1 (6.26) | 1.7 (6.39) | 5.6 (6.9) | 1.9 (6.9) | 2.1 (5.5) |
| Punta de la aleta anal | 5.6 (3.6) | 8 (3.86) | 2.3 (4.65) | 0.9 (3.38) | 3.2 (3.9) | 0.7 (2.6) | 1.1 (2.98) |
| Altura de D1 | 15.5 (9.96) | 19 (9.16) | 4.3 (8.69) | 2.3 (8.65) | 6.7 (8.3) | 2.3 (8.5) | 3.4 (9.15) |
| Base de D1 | 16.2 (10.40) | 25.6 (12.34) | 4.9 (9.90) | 2.6 (9.77) | 8.1 (10.1) | 2.5 (9.0) | 3.4 (9.15) |
| Punta de D1 | 9.4 (6.04) | 10.8 (5.2) | 2.2 (4.44) | 1.0 (3.76) | 4.1 (5.1) | 1.0 (3.5) | 1.5 (3.919 |
| Altura de D2 | 12.7 (8.16) | 15.2 (7.33) | 3.3 (6.67) | 1.7 (6.39) | 5.7 (7.0) | 1.9 (7.1) | 2.5 (6.62) |
| Base de D2 | 13.8 (8.86) | 17.7 (8.53) | 4.2 (8.48) | 2.3 (8.65) | 7.1 (8.7) | 1.8 (6.4) | 3.0 (8.04) |
| Punta de D2 | 7.8 (5.01) | 9.1 (4.39) | 1.7 (3.43) | 0.8 (3.01) | 3.5 (4.3) | 1.2 (4.3) | 1.3 (3.35) |
| Lóbulo ventral | 15.0 (9.63) | 19.2 (9.27) | 4.7 (9.49) | 2.3 (8.65) | 8.8 (11.0) | 2.7 (9.9) | 4.0 (10.64) |
| de aleta caudal | |||||||
| Lóbulo dorsal | 52.0 (33.40) | 65.9 (31.8) | 14.2 (28.69) | 7.9 (29.7) | 23.4 (29.0) | 8.0 (29.1) | 10.5 (28.04) |
| de aleta caudal | |||||||
| Punta de la aleta caudal | 4.9 (3.15) | 11.1 (5.4) | 2.2 (4.44) | 0.8 (3.01) | 4.3 (5.3) | 1.5 (5.5) | 2.0 (5.23) |
| Ancho del orificio nasal | 1.9 (1.22) | 1.3 (0.63) | 0.7 (1.41) | 0.6 (0.7) | 0.2 (0.8) | 0.4 (0.97) | |
| Barbilla izquierda | 4.6 (2.95) | 4.1 (1.98) | 1.8 (3.64) | 0.9 (3.38) | 2.3 (2.9) | 1.0 (3.7) | 1.2 (3.22) |
| Barbilla derecha | 4.7 (3.02) | 4.0 (1.93) | 1.8 (3.64) | 0.9 (3.38) | 2.4 (3.0) | 1.1 (4.1) | 1.2 (3.22) |
Una especie de tiburón de tamaño mediano (cercano a los 300cm de LT) que pertenece al género Ginglymostoma sobre la base de los siguientes caracteres: barbillas nasales largas, alcanzando la boca; ojos y abertura branquiales dorsolaterales; aletas pectorales, dorsales y anal con los ápices redondeados; segunda aleta dorsal y aleta anal mucho más pequeñas que la primera dorsal. Los siguientes caracteres la distinguen de la única otra especie del género descrita a la fecha: inserción de la primera aleta dorsal ligeramente anterior al origen de las pélvicas; los extremos posteriores de las aletas pélvicas, sin considerar en los machos los mixopterigios, alcanzan el origen de la segunda aleta dorsal, cuando están plegadas al cuerpo; distancia interdorsal corta, quedando el extremo posterior de la primera cerca del origen de la segunda; parte distal de la segunda aleta dorsal ligeramente anterior al origen del lóbulo dorsal de la aleta caudal, cuando está plegada al cuerpo; el extremo posterior de la aleta anal (sin estar plegada) alcanza el origen inferior de la aleta caudal; longitud prebranquial corta; distancia entre la axila de las aletas pélvicas y el origen de la aleta anal relativamente corta, igual a 1-1.3 veces la base de la primer aleta dorsal; barbillas nasales bien desarrolladas; ojos pequeños, su diámetro 0.7-1.4% de la LT y cabe más de 8 veces en la longitud interorbital; dentículos dérmicos con 5 a 6 quillas; dientes con una cúspide central alta, de menor tamaño que la base del mismo, en sus bordes flanquean de manera casi simétrica 4 o más cúspides secundarias obtusas, el número de ellos en la mandíbula superior e inferior es de 27 a 30, 33 a 35, respectivamente.
DescripciónEn la tabla 1 se muestran los datos morfométricos del holotipo (fig. 1) junto con el resto de la serie tipo. Cuerpo alargado, tronco ancho y robusto en la parte anterior; su máxima altura se presenta en la parte media, entre la axila de las aletas pectorales y las pélvicas, abarcando de 6.6 a 8.7 veces la LT. Región cefálica y prebranquial corta, la segunda comprendiendo de 5.5 a 6.8 veces la LT; hocico redondeado, ancho y muy corto, su longitud menor que la distancia interorbital; boca inferior pequeña, posicionada anterior a los ojos, su amplitud ligeramente mayor que la del hocico y con 2 barbillas carnosas localizadas en el margen anterior de las narinas; la longitud de las babillas es igual o mayor que la distancia preoral y caben 2 a 2.7 veces en la longitud del hocico; las narinas están conectadas con la boca, se presentan hendiduras nasorales; poseen 5 aberturas branquiales, las 2 últimas muy juntas y junto con la tercera están insertas por encima y detrás de las aletas pectorales (fig. 2); los ojos son ovalados y pequeños, su diámetro 0.7-1.4% de la LT y abarcan más de 8.5 veces la longitud interorbital; hay presencia de espiráculo de tamaño reducido, menor al diámetro ocular.
Presenta 2 aletas dorsales, la segunda ligeramente más pequeña que la primera, ambas con margen anterior ligeramente convexo y su margen distal con una ligera concavidad; esta característica es más marcada en ejemplares adultos, siendo en juveniles redondeada (fig. 3); la inserción de la primera dorsal está sobre o por delante del origen de las aletas pélvicas, su altura equivale a 1.2-2.9 veces la longitud del borde interno de las pélvicas, su extremo posterior alcanza el origen de la segunda dorsal dando como resultado un espacio interdorsal corto y representa 4.2-5.6% de LT; la segunda dorsal tiene su inserción muy por delante del origen de la aleta anal, al ser plegada su extremo posterior alcanza el origen del lóbulo dorsal caudal; aletas pectorales grandes, margen anterior recto y levemente convexo, extremo distal romo, región posterior ligeramente cóncavo; aleta anal de menor tamaño que la segunda dorsal, su inserción antecede el extremo posterior de la base de la segunda dorsal, mientras que el extremo posterior (sin estar plegada) alcanza el origen del lóbulo inferior de la aleta caudal, y al estar plegada sobrepasa esta distancia; el espacio entre la axila de las aletas pélvicas y el origen de la anal es pequeño y en ella comprende de 1 a 1.3 veces la longitud de la primera dorsal; el lóbulo dorsal de la aleta caudal representa 26.5-33.5% de LT.
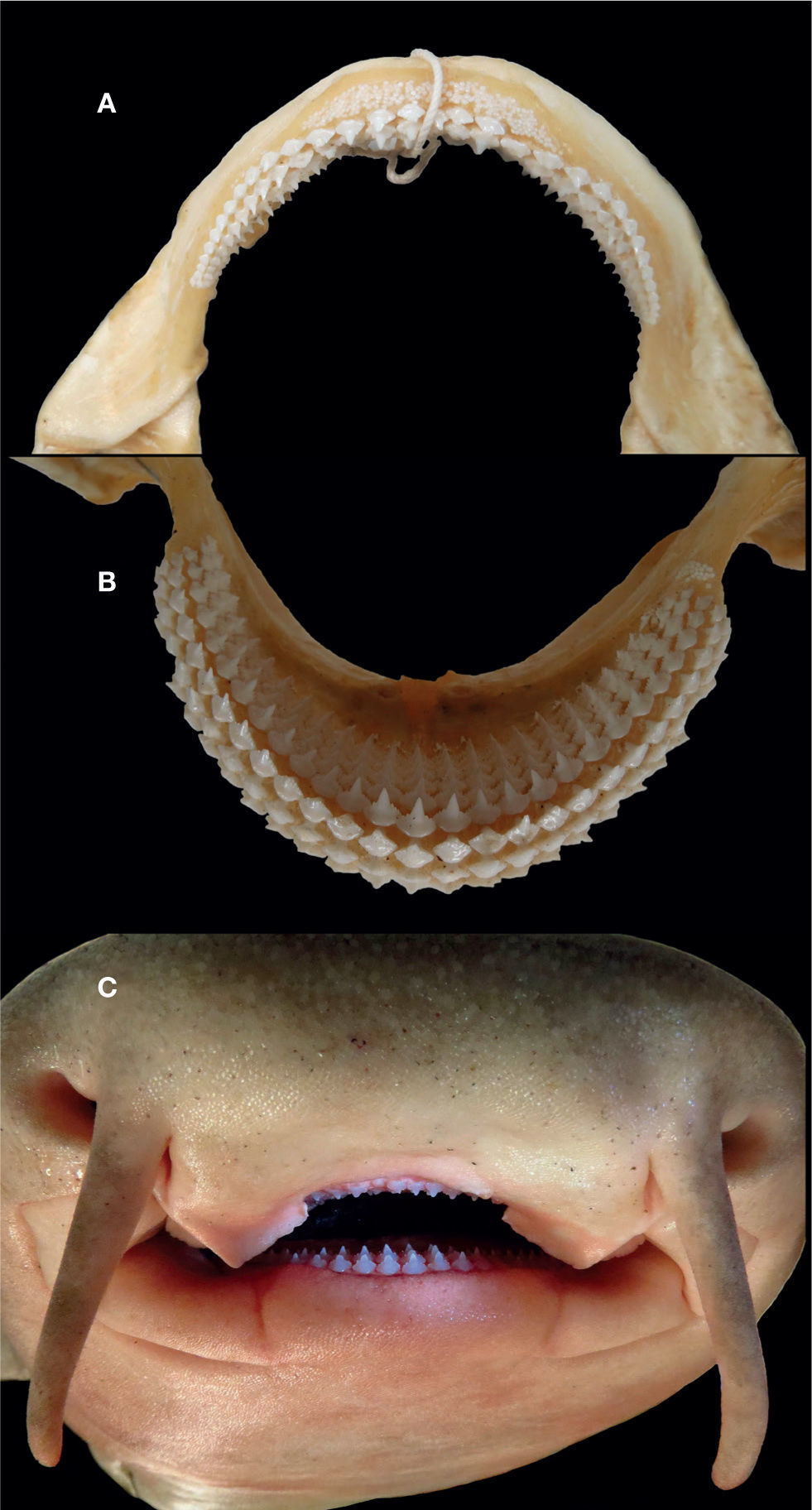
Los dientes son multicúspides y pequeños (fig. 4), presentan una cúspide central bordeada de manera simétrica de 4 o más cúspides laterales o secundarias; los dientes de ambas mandíbulas son muy similares, la fórmula dental obtenida de los ejemplares analizados es:
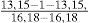
se observó una ligera variación, de uno a 3 dientes en ejemplares inmaduros. Los dentículos dérmicos son pequeños y tienen forma rómbica (fig. 5), su anchura es igual o mayor a su largo; su extremo terminal es angular; el margen anterior cuenta con marcadas muescas que corresponden con las quillas dorsales. En la parte dorsal de los dentículos están presentes de 5 a 6 quillas que corren en dirección paralela a su longitud, son obtusas y bajas, lo que los hace lisos al tacto, la quilla central es la más alta, la cual corre a lo largo de todo el dentículo, las restantes son más altas en la parte anterior del dentículo y disminuyen conforme se aproximan al extremo posterior.
ColoraciónEn vivo, el ejemplar adulto presenta una coloración parda y tiene variantes pálidas con tintes amarillos hasta oscuros. Los organismos inmaduros presentan un patrón de manchas oscuras en todo el cuerpo, cuyo tamaño es menor que aquel del diámetro ocular, excepto en la región ventral cefálica. Tanto en estados juveniles como en adultos la coloración del vientre es más clara, con tonalidades blancas a rosáceas, a la observada en el dorso de los organismos. La mayoría de los ejemplares preservados tienden a presentar una coloración dorsal parda con tintes verdosos y la región pélvica es, por lo general, de coloración amarillenta.
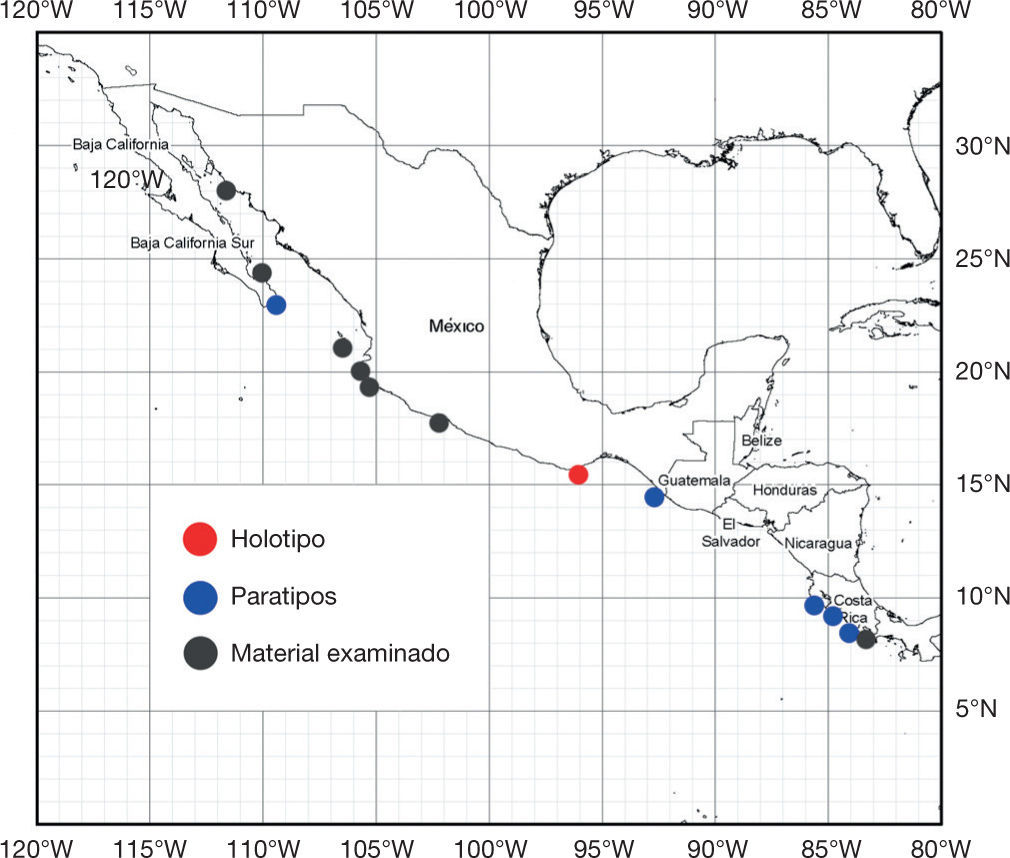
DistribuciónDesde el sur de la costa occidental de Baja California, México, hasta Perú, incluyendo el golfo de California; se considera un elemento endémico del Pacífico oriental tropical (fig. 6).
ParásitosEl género Ginglymostoma se encuentra parasitado por 21 especies de metazoarios a lo largo de su intervalo de distribución, incluyendo una especie de monogéneo, 15 especies de céstodos, 2 de nemátodos, 2 de copépodos y 1 de hirudíneo (Pollerspöck, 2014). La mayor parte de esos registros corresponden a la especie G. cirratum de la costa Atlántica. Sin embargo, para la que aquí se describe como nueva especie, Ginglymostoma unami sp. nov., se cuenta únicamente con los registros documentados por Caira y Euzet (2001), quienes identificaron 2 especies de céstodos, Pedibothrium brevispine y Pedibothrium manteri como parásitos de la válvula espiral en ejemplares recolectados en San José del Cabo, Baja California Sur, México. De manera interesante, los autores discutieron que la distribución anfiatlántica de los céstodos (dado que también habían sido registrados en tiburones en la costa Atlántica) era resultado de un evento vicariante posterior al surgimiento del istmo de Panamá, sugiriendo que estas especies de céstodos han permanecido sin cambios por ese periodo de tiempo y reconociendo la importancia de establecer diferencias genéticas entre individuos de Pedibothrium Linton, 1909 en ambos lados de la barrera geográfica. Los resultados de este trabajo, en cuanto al reconocimiento de una especie nueva de tiburón, refuerzan aún más la necesidad de obtener secuencias de ADN para los helmintos, pues existe la posibilidad de que éstas resulten ser especies crípticas (Nadler y Pérez-Ponce de León, 2011;1 Pérez-Ponce de León y Nadler, 2010). Claramente el evento vicariante determinó el evento de especiación del hospedero y es necesario explorar si éste causó el mismo efecto sobre las poblaciones de parásitos.
Resumen taxonómicoHolotipoCNPE-IBUNAM 18850, 1 organismo macho, adulto, con 155.7cm de LT, recolectado con red de enmalle de 6” de luz de malla, que operaba cercano en la zona rocosa frente a punta Salchii, a 7.5millas náuticas al E de Puerto Ángel. Localidad tipo: Punta Salchii, Puerto Ángel, Oaxaca, México, 15°40’ N, 96°21’ O, fecha de recolecta: 6 de septiembre de 2013; colectores: Fidencio Spíndola Ávila y Fidencio Spíndola Ramírez.
Paratipos6 ejemplares. 1) UCR 3011-01, 1 organismo macho en estado de madurez, 207.5cm LT, capturado el 18 de julio de 2014, por medio de anzuelo y carnada muerta (sardina) a fondo, frente al hotel Rancho Pez Vela, Golfito, Puntarenas, Costa Rica, 8°38’07.29” N, 83°13’05.35” O, a 7 m de profundidad; colectores: Federico Hampl Aguel, Christopher Hampl Millán y Claudio Iglesias Ginel; 2) CNPE-IBUNAM 9492, 1 organismo macho juvenil, 49.5cm LT, capturado el 19 de septiembre de 1993, por medio de anzuelo en San José del Cabo, Baja California Sur, México; 3) CPFES-I-155, 1 organismo hembra, embrión de 26.6cm de LT, como parte de una camada de una hembra capturada en Puerto Madero, Chiapas, México; colector: Francisco Sancho; 4) UCR 0880-1, 1 organismo macho, juvenil de 80.6cm LT, capturado el 11 de junio de 1975, en bahía Salinas, Guanacaste, Costa Rica, 11°03’ N, 85°41’ O; colector: Olmán Alán; 5) UCR 0025-16, 1 organismo hembra, embrión de 27.4cm LT, capturado el 7 de abril de 1966, con rotenona, en una poza de marea en playas del Coco, Guanacaste, Costa Rica, 10°33’ N, 85°42’ O; colector: Herbert Nanne; 6) UCR 0200-23, 1 organismo macho, neonato de 37.4cm LT, capturado el 10 de septiembre de 1967, con rotenona, en una poza de marea en playas Tamarindo, Guanacaste, Costa Rica, 10°17’ N, 85°51’ O; colector: Peter L. Weaver.
Material adicional examinado25 ejemplares completos y 3 mandíbulas. CNPE-IBUNAM s/núm. de catálogo (ex ITEMS 81-54), 1 organismo de sexo indeterminado, 532mm LT, capturado el 28 de marzo de 1981, en Las Bucas, Huatabampo, Sonora, México; CIICMYL-P311.01, 1 organismo macho, 182mm LT, capturado con red de arrastre el 29 de abril de 1959, en la isla Madre, Islas Marías, Nayarit, México; CIICMYL-P311.02, 1 organismo macho, 254mm LT, capturado el 29 de abril de 1959, en la isla Madre, Islas Marías, Nayarit, México; CICIMAR-CI 892, 13 organismos en estado embrionario, 5 machos, 6 hembras y 2 indefinidos, 14.2-20cm LT, colectados al lado E de isla Cerralvo, La Paz, Baja California Sur, México; ENCB-IPN-LEM 8079, 5 organismos embriones, 252-275cm LT, provenientes de una hembra capturada en Barra de Navidad, Jalisco, México; UCR 0755-01, 1 organismo macho en estado embrionario, 28.7cm LT, capturado el 4 de febrero de 1973, en playa Sámara, Guanacaste, Costa Rica, 09°52’ N, 85°31’ O, colector: Jorge A. Ramírez; UCR 128-29, 3 organismos, 1 macho (46.5cm LT) neonato y 2 hembras (27.5-52.7cm LT) capturados el 27 de enero de 1967, con rotenona en una poza de marea frente a playa Tamarindo, Guanacaste, Costa Rica, 10°18’ N, 85°50’ O, colectores: William A. Bussing, O. Blanco y C. Kalb; IGM-CMR-46, mandíbula de ejemplar adulto, capturado en julio de 1980, isla Isabela, Nayarit, México; CP-UMAR s/núm. de catálogo, mandíbula de 1 organismo macho adulto, de 220 LT, capturado el 20 de agosto de 2008, en Petacalco, Guerrero, México (figs. 3A, B); CP-UMAR s/núm. de catálogo, mandíbula de 1 organismo hembra, 140cm LT, capturado el 28 de mayo de 2011, en Chipehua, Oaxaca, México.
EtimologíaEl epíteto específico unami se refiere al acrónimo de la Universidad Nacional Autónoma de México (UNAM), en reconocimiento al papel trascendental que ésta tiene en la educación del pueblo mexicano.
Comentarios taxonómiosGinglymostoma unami sp. nov. difiere de su congénere, G. cirratum, por varios caracteres: la inserción de la aleta dorsal se presenta anterior al origen de las aletas pélvicas, mientras que la inserción de la segunda aleta dorsal se encuentra a nivel del origen de las aletas pélvicas, o posterior a éstas; en la nueva especie el extremo posterior de las aletas pélvicas, una vez plegadas, alcanzan o están cercanos al origen de segunda dorsal (sin incluir en los machos los mixopterigios); por su parte en G. cirratum estas no alcanzan la inserción de la segunda dorsal. El espacio interdorsal es otra característica que distingue a ambas especies, siendo más corto en G. unami sp. nov. (3.6-5.6% de LT), esto se debe a que en la nueva especie el extremo de la primera dorsal alcanza la inserción de la segunda dorsal, siendo más evidente en organismos maduros, mientras que en G. cirratum el extremo posterior de la segunda dorsal no alcanza la inserción de la segunda dorsal (5.4-9.5% de LT); en G. unami sp. nov. el extremo posterior de la segunda dorsal alcanza o está ligeramente delante del inicio del lóbulo superior de la aleta caudal, siendo más evidente en ejemplares adultos, mientras que en G. cirratum el extremo posterior no alcanza el origen de dicha aleta; sin estar plegada la aleta anal, en G. unami sp. nov. su extremo alcanza o sobrepasa el origen de la inserción inferior de la aleta caudal, lo que no sucede en G. cirratum; la distancia que hay entre la axila de las aletas pélvicas con respecto al inicio de la aleta anal también distingue a ambas especies, siendo menor en G. unami sp. nov., donde esta distancia comprende menos de 1.2 veces la base de la primer aleta dorsal, mientras que en G. cirratum comprende más de 1.4 veces; en conjunto las características anteriores remarcan en G. cirratum un cuerpo más grácil que en G. unami sp. nov., donde se hace el tronco más robusto y corto, e incluso la región prebranquial es más corta (14.5-18% de LT) en comparación con G. cirratum (12-15% de LT).
La nueva especie tiene dientes con mayor número de cúspides secundarias dispuestas simétricamente con respecto a la cúspide central, además, sus extremos no son agudos, sino bajos y romos (fig. 4), en cambio, Luer, Blum y Gilbert (1990) observaron un menor número de cúspides laterales con extremos agudos característicos de los dientes de G. cirratum. Además, los dentículos dérmicos en G. unami sp. nov. son rómbicos con 5 a 6 quillas (fig. 5), en comparación con los de G. cirratum, donde son más elongados y presentan menor número de quillas; estas diferencias coinciden con lo documentado por Castro (2011a). Cabe señalar que en las descripciones y diagnosis de G. cirratum que se basan en ejemplares colectados en el Atlántico (por ejemplo, Baughman y Springer, 1950; Bigelow y Schroeder, 1948; Garman, 1913) se distinguen con claridad las diferencias enunciadas con anterioridad para esta especie.
Por último, la distribución de ambas especies es disyunta, estando la nueva especie que se describe restringida al Pacífico oriental tropical (fig. 6), mientras que G. cirratum se encuentra en costas del Atlántico. De hecho, Castro (2011a) había sugerido que las poblaciones de Ginglymostoma del Atlántico y del Pacífico provenían de un ancestro común y que habían quedado aisladas desde hace más de 4millones de años al ocurrir el último cierre del istmo de Panamá. Diferencias morfológicas entre ambas poblaciones ya habían sido reconocidas por otros autores como Castro-Aguirre, Espinosa-Pérez y González-Acosta (2010) y Castro (2011a), los cuales designaban a la población del Pacífico como Ginglymostoma sp. Este patrón de separación geográfica también ya había sido evidenciado recientemente por medio de marcadores moleculares (Karl et al., 2012), sin embargo, la especie no había sido propiamente descrita ni nombrada. La distribución de especies hermanas de peces en ambas costas del continente americano ha sido también observado en otros grupos e incluso en otras especies de tiburones como en Carcharhinus porosus (Ranzani, 1839) (Atlántico) y Carcharhinus cerdale Gilbert, 1898 (Pacífico) (Castro, 2011b).

Clave para la identificación de las especies de la familia Ginglymostomatidae, adaptación deCompagno (2002)
- 1a.
Barbillas nasales cortas, no alcanzan el borde anterior de la boca; labio inferior no trilobulado y sin surcos oralabiales conectando el borde del borde del mismo con los surcos labiales inferiores; segunda aleta dorsal y aleta anal de tamaño similar a la primera dorsal; aleta caudal corta, menos de 20% de LT (océano Índico occidental).....................................Pseudoginglymostoma brevicaudatum
- 1b.
Barbillas nasales largas, sobrepasando el borde anterior de la boca; labio inferior trilobulado y con surcos oralabiales conectando el borde del mismo con los surcos labiales inferiores; segunda aleta dorsal y aleta anal mucho más pequeñas que la primera dorsal; aleta caudal larga, más de 20% de LT.....................................2
- 2a.
Ojos y aberturas branquiales ubicados lateralmente; dientes laterales más o menos comprimidos y dispuestos en series imbricadas; aletas pectorales, dorsales y anal con los ápices angulares, las pectorales estrechas y falcadas (océanos Índico y Pacífico occidental y central).....................................Nebrius ferrugineus
- 2b.
Ojos y aberturas branquiales ubicados dorso-lateralmente; dientes laterales no comprimidos y no en series imbricadas; aletas pectorales, dorsales y anal con los ápices redondeados, las pectorales amplias y no falcadas.....................................3
- 3a.
Región prebranquial 12.0-15.0% de LT; la distancia entre la axila de las aletas pélvicas y el origen de la aleta anal comprende más de 1.4 veces la base de la primer aleta dorsal; el extremo de la primera dorsal no alcanza la inserción de la segunda dorsal; espacio interdorsal 5.4-9.5% de LT; el extremo posterior de la segunda dorsal alcanza o está ligeramente delante del inicio del lóbulo superior de la aleta caudal; el extremo de la aleta anal no alcanza el origen de la inserción inferior de la aleta caudal; dentículos dérmicos elongados (más largos que anchos), con menos de 5 (océano Atlántico).....................................Ginglymostoma cirratum
- 3b.
Región prebranquial 14.5-18.0% de LT; la distancia entre la axila de las aletas pélvicas y el origen de la aleta anal comprende menos de 1.2 veces la base de la primer aleta dorsal; el extremo de la primera dorsal alcanza la inserción de la segunda dorsal; espacio interdorsal 3.6-5.6% de LT; el extremo posterior de la segunda dorsal no alcanza el inicio del lóbulo superior de la aleta caudal; el extremo de la aleta anal alcanza o sobrepasa el origen de la inserción inferior de la aleta caudal; dentículos dérmicos oblongos, no elongados (más anchos que largos), con 5 a 6 quillas (océano Pacífico oriental).....................................Ginglymostoma unami sp. nov.
La descripción que se hace en este trabajo de Ginglymostoma unami sp. nov. contribuye con el inventario taxonómico de los elasmobranquios de México que a la fecha cuenta con 211 especies, de las cuales 109 son de tiburones (Del Moral y Pérez-Ponce de León, 2013). La incorporación de datos moleculares al trabajo taxonómico de rutina, adicionalmente al estudio detallado de los organismos que están depositados en colecciones biológicas, redundará seguramente en el descubrimiento de más especies de elasmobranquios que son habitantes de los mares mexicanos.
Material comparativo examinado: Ginglymostoma cirratumCinco especímenes completos, más 1 mandíbula. CIICYML 311.03, 1 organismo hembra, 50.5cm LT, capturado con red de arrastre el 7 de enero de 1964, en la Laguna de Términos, Ciudad del Carmen, Campeche, México; UCR 0351-27, 1 organismo hembra, 30. 7cm LT, capturado el 23 de abril de 1969, con redes de arrastre a una profundidad de 35-55 m, al NE de Gorda Bank, Caribe de Honduras; UCR 0252-22, 1 organismo hembra, 33.1cm LT, capturado 30 de mayo de 1968, con redes de arrastre a una profundidad de 20-24 m, entre Puerto Limón y Tortuguero, Limón, Mar Caribe, Costa Rica. colectores: William A. Bussing y R. T. Nishimoto; UADY s/catálogo, 1 ejemplar hembra, recolectado en Yucalpetén, Yucatán, México; ENCB-IPN-LEM 8147, 1 organismo adulto 100.2cm LT, recolectado en el Atlántico de México; IGM-CMR-45, mandíbula de 1 organismo juvenil, obtenido del mercado de La Viga, Distrito Federal, México.
AgradecimientosSe agradece el apoyo en las diferentes fases del trabajo a los integrantes del laboratorio de pesquerías de la UMAR: Dr. Vicente Anislado-Tolentino; así como al laboratorio de bioestadísticas y pesquerías de la FES-Zaragoza: Dr. Isaías H. Salgado-Hugarte y M. en C. Óscar Uriel-Mendoza. A los Biólogos Armando Martínez y Jonathan De la Cruz por el apoyo en las diversas fases del proyecto. A la M. en C. Berenit Mendoza-Garfias por la asistencia y preparación de muestras en el microscopío electrónico de barrido (Laboratorio de Microscopía Electrónica del Instituto de Biología, UNAM). A los diversos curadores de las colecciones de referencia por permitir la revisión de los ejemplares: Dra. Lizbeth Chumba-Segura, Dra. Mirna I. López, Dra. Ana Fabiola Guzmán-Camacho, M. en C. Héctor Espinosa-Pérez, Dr. José De la Cruz-Agüero, Dr. Felipe Amezcua-Linares, Dr. Jesús Alvarado-Ortega, Ing. Francisco Sancho y Biol. José Antonio Martínez-Pérez. Agradecemos muy en especial el apoyo brindado por los integrantes y autoridades del Museo de Zoología y de la Escuela de Biología, Universidad de Costa Rica, en el traslado, medición, fijación, depósito y resguardo de uno de los paratipos: Carlos Garita, Miguel Marrero, Fiorella Vásquez, Gustavo Arias, Rita Vargas, Kimberly García, Odalisca Breedy, Ana Rosa Ramírez y Carolina Méndez. En este mismo sentido se agradece y se hace el reconocimiento a Federico Hampl-Aguel, Christopher Hampl-Millán y Claudio Iglesias-Ginel quiénes capturaron el espécimen más grande de la serie tipo. El primer autor agradece al Posgrado en Ciencias Biológicas por la formación en el posgrado y al Conacyt por la beca otorgada para desarrollar sus estudios.