





Se realizó un estudio faunístico de la comunidad de las arañas de 4 oasis de Baja California Sur. Los organismos fueron recolectados en ambientes xérico y mésico con trampas de caída y captura manual y por medio de golpeo y con redes de manta. Se capturaron 34 familias, 130 géneros y 243 especies que representan el 63.2% de las registradas para la península de Baja California. Un género y 11 especies son citados por vez primera para México, y 13 géneros y 18 especies son nuevos registros para Baja California Sur. Las familias Salticidae (36), Araneidae (29), Theridiidae (27) y Gnaphosidae (24) fueron las de mayor riqueza específica. En el ambiente xérico se registraron 32 familias, 125 géneros y 215 especies, y en el mésico 26 familias, 96 géneros y 157 especies, con 128 especies compartidas. Ochenta y siete especies fueron únicas en el ambiente xérico y 29 en el mésico. La complementariedad entre ambientes fue del 47.5%. En el suelo se registraron 28 familias, 89 géneros y 144 especies, y en la vegetación 16 familias, 45 géneros y 119 especies. Ambos estratos compartieron 19 especies, 125 especies se registraron solo del suelo y 100 en el matorral con una complementariedad de 92,2%.
A spider inventory from 4 oases at the xeric and mesic environments was undertaken. Ground and vegetation spiders were captured with pit fall traps and collected by sweeping and beating. Thirty four families, 130 genera and 243 species were captured which represent 63.2% of the species recorded for the Baja California Peninsula. One genus and 11 species are cited for the first time for Mexico, 13 genera and 18 species are new records for Baja California Sur. Salticidae (36), Araneidae (29), Theridiidae (27), and Gnaphosidae (24) families had the highest specific richness. Thirty two families, 125 genera, and 215 species are reported for the xeric environment and 26 families, 96 genera, and 157 species for the mesic environment. Both environments shared 128 species; 87 and 29 species were exclusive to xeric shrub and mesic vegetation, respectively. Complementarity between xeric and mesic environment was 47.5%. Twenty eight families, 89 genera, and 144 species, and 16 families, 45 genera and 119 species were captured in soil and vegetation strata, respectively with a complementarity of 92.2%.
Las zonas áridas del norte de México ocupan el 65% del territorio nacional y son altamente diversas debido a su compleja fisiografía originada durante el Cenozoico (Cartron, Ceballos y Felger, 2005). Por su actual posición latitudinal presentan alta y constante radiación solar (Ferrusquía-Villafranca, González-Guzmán y Cartron, 2005), escasez e irregularidad de lluvias (125-400mm promedio de los máximos anuales), humedad ambiental variable (Cartron et al., 2005; Cloudsley-Thompson, 1983), aunado con una irregular carencia de agua y temperatura extrema (5-12°C ->28°C promedio de los máximos anuales); estos factores han contribuido a la prevalencia del ambiente árido que las caracteriza. Inmersos en estas regiones se encuentran numerosos cuerpos de agua que emergen del subsuelo y arroyos de temporal que dan origen a los oasis, ambientes de vital importancia para el establecimiento de asentamientos humanos y como hábitat de una biota relicta que contrasta con la que se halla en las zonas circundantes (Arriaga, 1997; Ezcurra, Felger, Russell y Equihua, 1988; Grismer, 2002; Grismer y McGuire, 1993).
En la península de Baja California se han encontrado alrededor de 184 oasis (Maya, Coria y Domínguez, 1997), de los cuales 60 son considerados como oasis típicos por presentar un cuerpo de agua dulce con vegetación asociada (Rodríguez-Estrella, Blázquez y Lobato, 2005). Aunque estos sitios representan el 1% de la superficie total de la península, son considerados como ambientes únicos en México, porque presentan características biogeográficas, ecológicas y evolutivas particulares semejantes a una isla, que representan unidades ecológicas discretas (Grismer y McGuire, 1993). Además, porque albergan una considerable diversidad biológica importante de poblaciones raras, endémicas o relictas, y exclusivas de estos ambientes (Luja-Molina, 2011). Por estas razones, algunos de estos oasis han sido incorporados en la lista internacional RAMSAR de humedales de importancia internacional, de acuerdo con el convenio del mismo nombre que fue firmado en Ramsar, Irán. A pesar de su gran importancia y peculiaridad, poco se conoce de la riqueza específica y composición de las comunidades de organismos de estos sitios, entre los que se encuentra el megadiverso grupo de las arañas. Estudios preliminares señalan que de las 384 especies de arañas registradas en la península de Baja California (Berrian, 2014; Jiménez, 2011), tan solo se registraron 61 en los oasis, dominando el gremio de las errantes; asimismo, se han encontrado variaciones de las comunidades de arañas que se asociaron a diferencias en la riqueza florística de cada oasis (Jiménez, Palacios y Tejas, 1997; Llinas-Gutiérrez y Jiménez, 2004). Por otro lado, se destaca la urgencia por describir y corroborar especies ya descritas como, por ejemplo, Correa-Ramírez, Jiménez y García de León (2010), con base en análisis taxonómicos y moleculares, encontraron que lo que estaba definido como la especie Pardosa sierra Banks, 1898 (Familia Lycosidae), en realidad es un complejo de 3 especies, lo que llevó también a la conclusión de que P. sierra es endémica de los oasis de la península de Baja California, en particular de sus ambientes mésicos (Correa-Ramírez et al., 2010).
Asimismo, se ha comprobado que existe una relación estrecha de las arañas de los oasis con diferentes especies animales. Específicamente, se ha encontrado que organismos de diversas familias de arañas como Dictynidae, Anyphaenidae, Salticidae, Uloboridae, Tetragnathidae, Miturgidae y Philodromidae son presas de la avispa lodera parasitoide [Trypoxylon (Trypargilum) tridentatum tridentatum Packard 1867], lo cual sucede con mayor frecuencia en la zona mésica (Domínguez y Jiménez, 2008). Por otro lado, Nieto-Castañeda y Jiménez-Jiménez (2009) al estudiar 2 especies de arañas endémicas sintópicas del género Syspira (Miturgidae) (S. tigrina Simon 1895 y S. longipes Simon 1895), presas de algunos roedores (Álvarez-Castañeda, Correa-Ramírez y Trujano-Álvarez, 2006), encontraron que la coexistencia entre ambas está relacionada por las diferencia en tallas corporales y sus patrones de actividad temporal, no difiriendo en su distribución espacial entre los ambientes mésico y xérico. Otro aspecto que resalta de las arañas de este género es que poseen ciclos de vida muy peculiares, ya que presentan un solapamiento muy marcado en tallas corporales de diferentes estadios, resultado quizá de las diferentes tasas de crecimiento y el continuo reclutamiento de estos organismos, producto de ovoposición de las hembras durante todo el año. Estos patrones no han sido registrados para otras especies de arañas en las que han sido estudiados sus ciclos de vida (Nieto-Castañeda, Salgado-Ugarte y Jiménez, 2012).
Debido al panorama mencionado, es altamente probable que aún falte mucho por conocer y describir de la fauna aracnológica asociada a los oasis. Dada esta situación, el objetivo de este estudio es dar a conocer las especies de arañas asociadas a 4 oasis de Baja California Sur, 2 localizadas en la parte central y 2 en la región sur, particularizando en su distribución confinada en el gradiente mésico-xérico como una primera aproximación de la riqueza biótica que estos ambientes albergan. Debido a la gran cantidad de información generada durante este estudio, los análisis ecológicos generados serán publicados en otro manuscrito, ya que es complicado presentarlos en este mismo trabajo, dada su extensión.
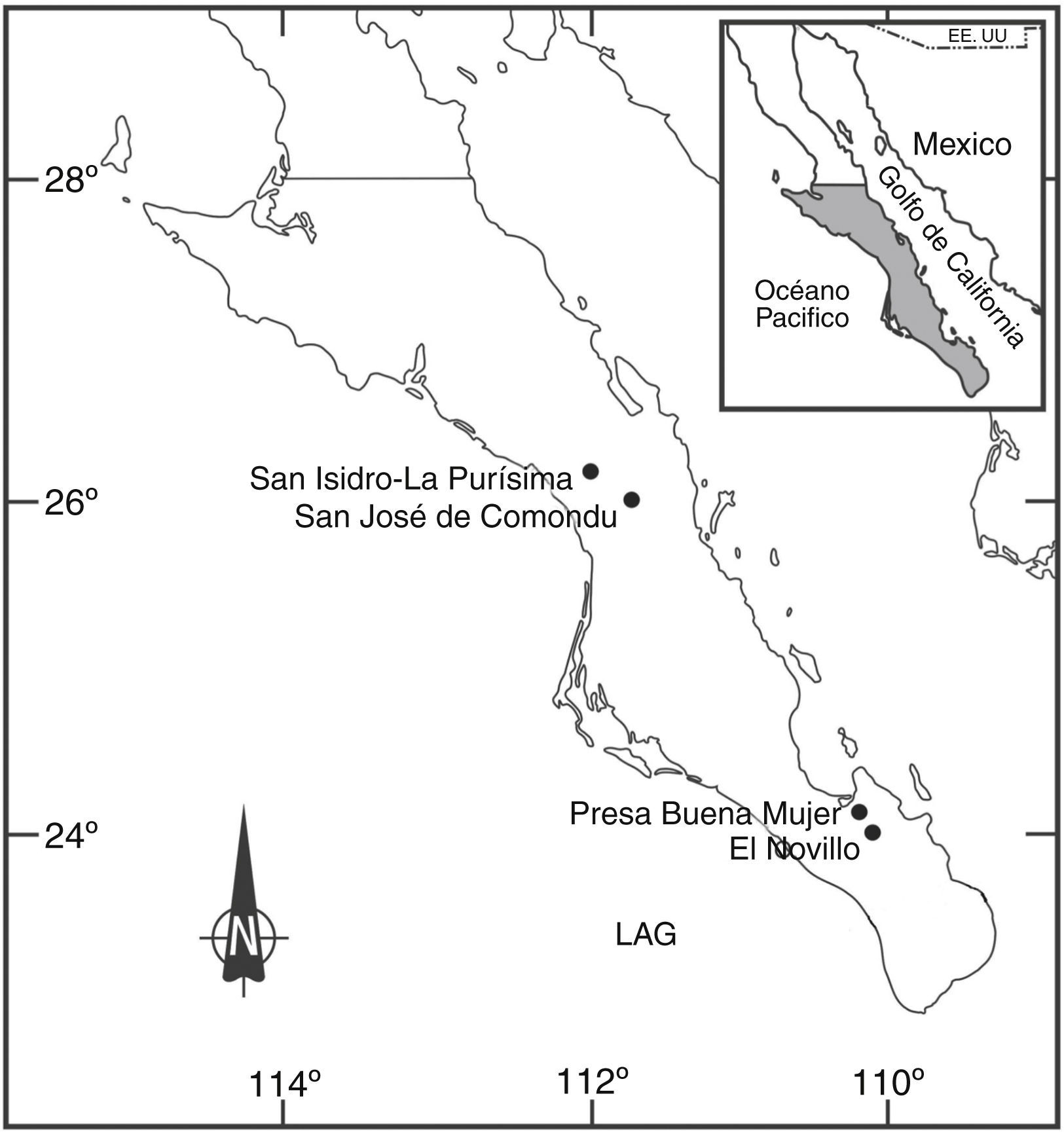
Materiales y métodosSe seleccionaron 4 oasis ubicados en Baja California Sur de acuerdo con su localización latitudinal con fines comparativos (fig. 1): 1) San Isidro-La Purísima (26o12’23” N, 112o02’54” O, 110m snm, superficie 2.25km2); 2) San José de Comondú (26o23’34” N, 111o49’13” O, 330m snm, superficie 0.88km2); 3) La Presa de la Buena Mujer (24°05’13” N, 110°11’19” O, 180m, superficie 0.86km2 y 4) arroyo El Novillo (23°55’40” N, 110°13’27” O, 220m) como parte de la subcuenca El Novillo (superficie 589.2km2). Los primeros 2 se localizan en la parte central del estado, el primero en el Llano Purísima Iray y el segundo en la sierra de la Giganta, en tanto que los 2 últimos se encuentran en la parte sur, en la región del Cabo. El clima de los 4 humedales es del tipo árido extremoso, con lluvias en verano y del tipo muy seco y semicálido (BWh) (García, 1981). El oasis de San Isidro-La Purísima forma parte de un arroyo con una cuenca de 4,966km2 de extensión. En su cabecera, bastante retirada del Pacífico, recolecta agua de un buen número de pequeños afluentes que descienden de la sierra vecina a bahía Concepción (Flores, 1998). El hábitat mésico está conformado por vegetación arbustiva Juncos acutus L. Prosopis sp., Vallesiana glabra (Cav.), Cryptostegia grandiflora (Roxb.) y arbórea como Washingtonia robusta H. Wendl y Phoenix dactylifera L. El hábitat xérico contiene comunidades de arbustos de tallos carnosos gruesos de aproximadamente 3m de altura, característicos del matorral sarcocaule como Jathropha cinerea (Ortega), Opuntia sp., Mammillaria sp. y Acacia brandegeana I. M. Jhtn. San José de Comondú está en la parte más superior del arroyo Comondú, aproximadamente a 22km al sureste de La Purísima y en el lado oeste de la sierra de la Giganta. Este arroyo está situado dentro de una cañada constituido por una cadena de pozas comunicadas por el cauce del arroyo en forma permanente (Flores, 1998). La vegetación dominante en el hábitat mésico es W. robusta, P. dactylifera y Xanthosoma sp., junto con cultivos de mango, aguacate, higos y cítricos. En el hábitat xérico dominan vegetación arbustiva característica del matorral sarcocaule, como Prosopis sp., J cinerea, A. brandegeana y V. glabra. El arroyo de temporal El Novillo se ubica en la sierra El Novillo o sierra las Calabazas y se asienta sobre suelos vertisoles, es decir, arcillosos con grandes cantidades de grava y rocas superficiales y xerosoles de escaso contenido de materia orgánica. La vegetación es arbórea del tipo selva baja caducifolia, con alturas de 3-5m como: J. cinerea, Cylindropuntia cholla Weber, Stenocereus thurberi (Engelm.), Haematoxylon brasiletto Karst., Justica californica Benth., Acasia sp. y Brusera sp., la vegetación mésica es escasa y es del tipo herbáceo. La presa de la Buena Mujer se ubica en la sierra de las Cruces al este de la ciudad de La Paz, donde se generan varias cuencas pequeñas, y es en una de ellas donde nace el arroyo El Cajoncito que alimenta a la presa (Inegi, 1981). La comunidad de matorral xerófilo es del tipo sarco-crasicaule dominado por cactáceas como Pachycereus pringlei (S. Watson), Machaerocereus gummosus Britton y Rose, Lophocercus schoottii Britton y Rose, Opuntia sp., Lemaireocereus thurberi Britton y Rose, Bursera sp y Prosopis sp., la vegetación mésica es escasa y está constituida principalmente por W. robusta (Domínguez, 2005; Jiménez-Jiménez y Palacios-Cardiel, 2010).

Se realizaron muestreos bimensuales, con duración de 2 días por localidad, de julio a diciembre del 2002 y de febrero a octubre del 2003 en los oasis San Isidro y en San José de Comondú, y de marzo a noviembre del 2004 en El Novillo y Presa Buena Mujer. En cada lugar se trazaron transectos al azar: 3 en San Isidro y San José de Comondú, y 2 en los oasis restantes. Cada transecto de 150m de longitud, se trazó perpendicular al cuerpo de agua, abarcando así los ambientes mésico y xérico de cada sitio. A lo largo de cada transecto se colocaron 15 trampas de caída a intervalos de 10m cada una, utilizando como conservador 250ml de anticongelante. Este tipo de trampa fue construida con 2 botellas de plástico, una de 600ml donde se depositó el conservante y otra de 2L; a ambas botellas se les hizo un corte transversal a unos 20cm de altura, para eliminar la parte del cuello, y solo el de la grande se utilizó como embudo para captar a las arañas que resbalaron para ser atrapadas en el recipiente más pequeño (Gurdebeke y Maelfait, 2002; Patrick y Hansen, 2013). Las trampas fueron enterradas a nivel del suelo y cubiertas con una malla metálica de 23×23cm, separada de la orilla del embudo a unos 3cm, suspendida por 4 patas metálicas, una en cada esquina, para apoyarse en el suelo. Cada 2 meses, el contenido se retiró y se coló del recipiente fijador, conservándolo en etanol al 70% para su transporte. Como complemento a las recolectas en el suelo, en cada transecto se hicieron capturas en el follaje de forma manual durante el día de 8:00 a 12:00h (4h/transecto), utilizando redes planas de manta de 1×1m, empleando una vara para golpear y sacudir la vegetación, para hacer caer y capturar a los organismos en un rango de 0.5-2m, durante aproximadamente 10-15min, para recolectarlos posteriormente utilizando aspiradores y pinceles. Estas recolectas fueron realizadas en un cuadro de 2×2 m en los mismos sitios donde fueron enterradas las trampas de caída.
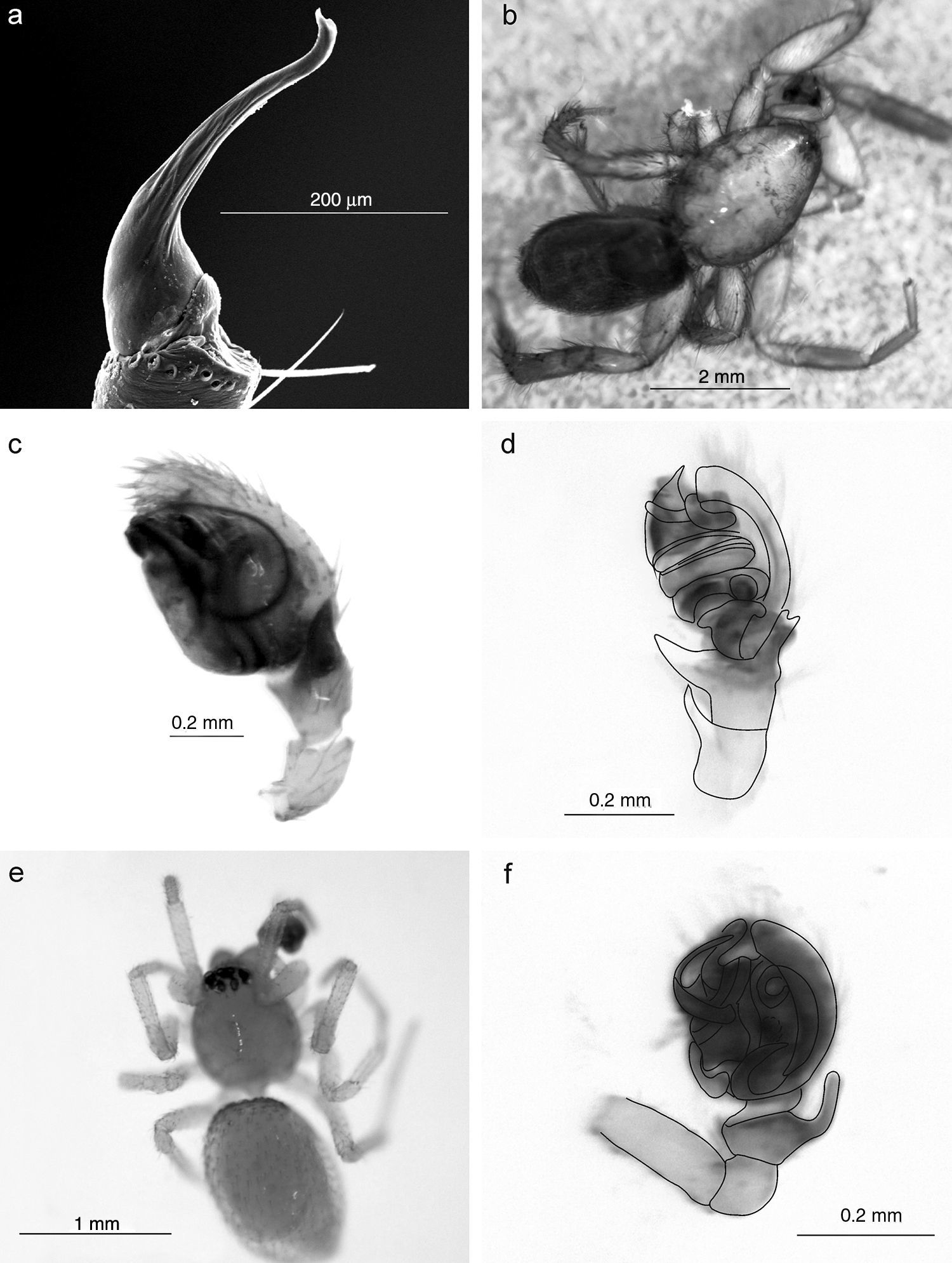
Todos los ejemplares (adultos y juveniles) recolectados fueron separados e identificados a nivel genérico de acuerdo con Ubick, Paquin, Cushing y Roth (2005) y a nivel específico con base en trabajos o revisiones taxonómicas de diversos autores (Griswold, 1987; Levi, 1953b, 1955, 1957, 1959a, 1959b, 1963a, 1963b, 1964; Platnick y Shadab, 1974, 1982, 1983, entre otros). Los juveniles fueron identificados a nivel genérico o específico cuando fue posible compararlos con los adultos. Los nombres de las especies fueron validados con base en el catálogo mundial de arañas (World Spider Catalog, 2015). Se ilustran con fotografías digitales únicamente los nuevos registros de géneros y especies para México. Las fotografías fueron hechas con una cámara digital DFC295, adaptada a un microscopio estereoscópico (MZ6, Leica Microsystems, Weztlar, Alemania) y el programa Helicon Focus 4.0 (www.heliconsoft.com) fue utilizado para editar las imágenes digitales. Las imágenes tomadas con un microscopio electrónico de barrido fueron elaboradas utilizando un microscopio modelo S-3000N, Hitachi, Tokyo, Japón. Todos los ejemplares fueron conservados en alcohol etílico al 70% y depositados en la colección aracnológica del Centro de Investigaciones Biológicas del Noroeste (CARCIB), La Paz, Baja California Sur, México. Para reconocer la complementariedad de la composición entre los ambientes mésico y xérico y en los estratos del suelo y vegetación, se utilizó el índice de Colwell y Coddington (1994) expresado como CAB=UAB/ SAB, donde SAB=a+b-c y UAB=a+b-2c; a=número de especies del sitio A, b=número de especies del sitio B y c=al número de especies en común en los sitios A y B, y refiere al grado de disimilitud en la composición de especies entre pares de biotas expresado en porcentaje de especies (figs. 2 y 3).


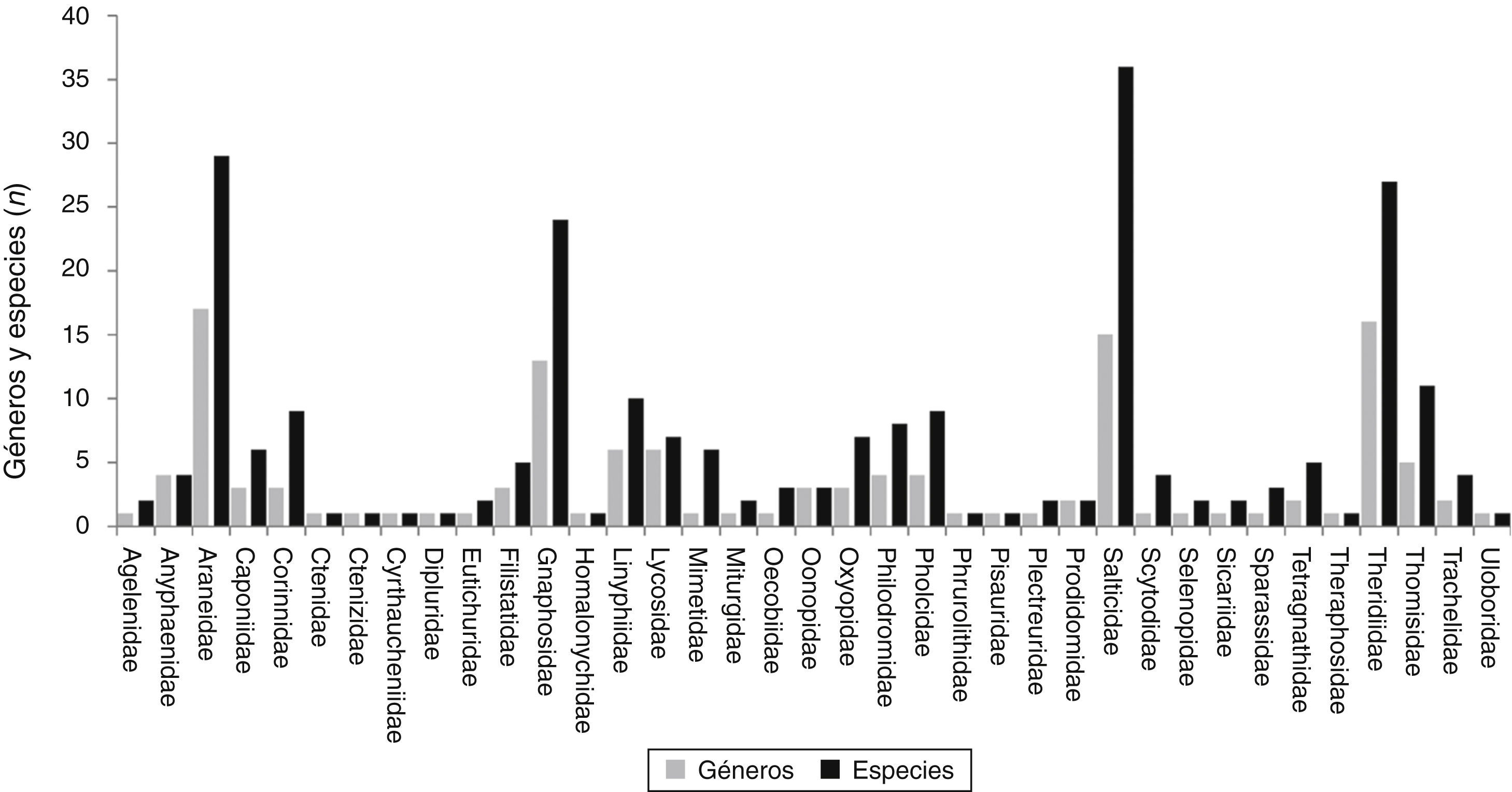
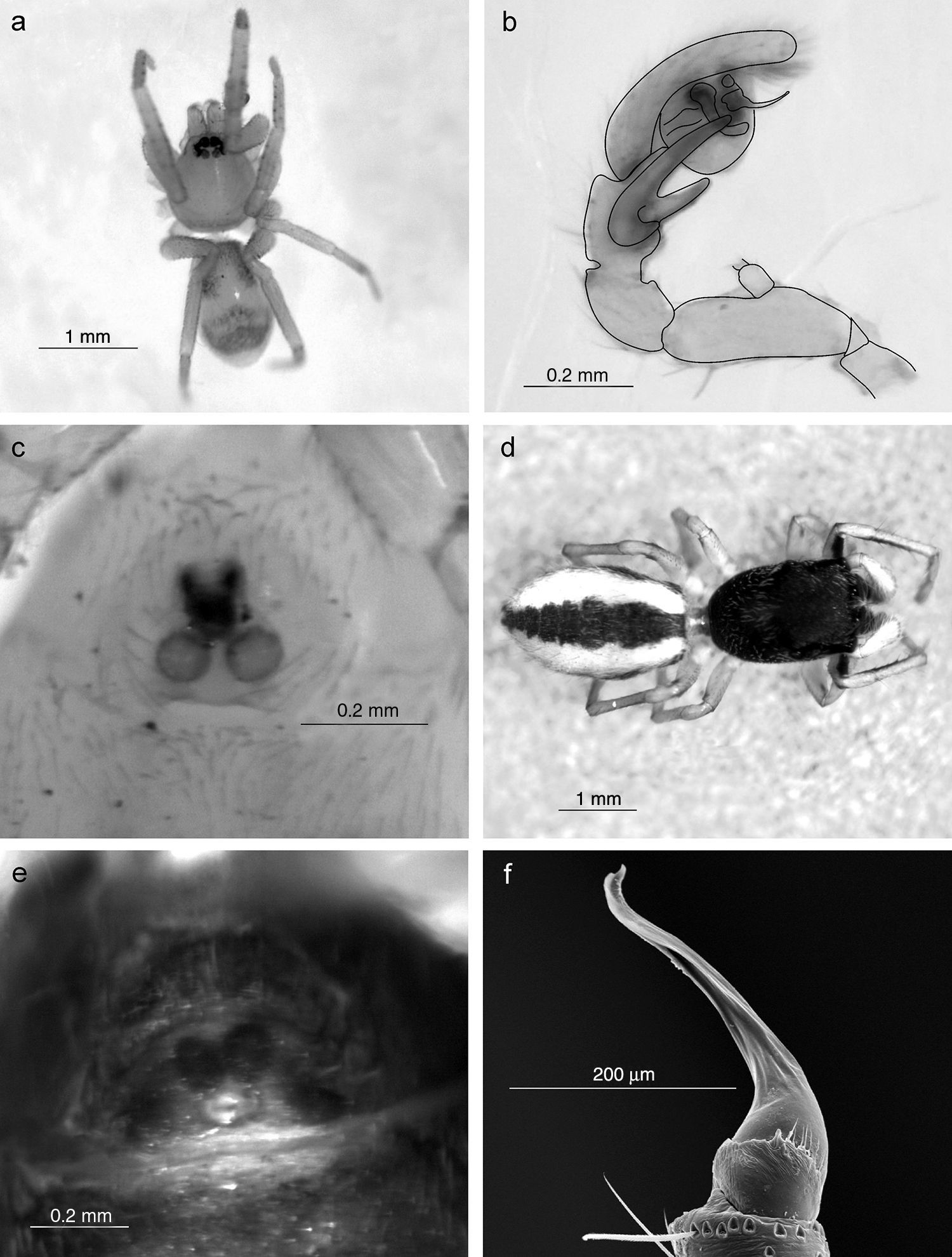
Se registraron un total de 243 especies de arañas pertenecientes a 130 géneros y 34 familias (Anexo). De este total, 41 morfoespecies (16.9%) solo fueron determinadas a nivel genérico por tratarse de ejemplares juveniles; 143 (58.8%) especies fueron identificadas a nivel específico, 59 (24.3%) fueron consideradas como especies no descritas y 4 se identificaron como especies afines a las descritas. Sin embargo, este material requerirá de un mayor análisis taxonómico para su confirmación. Un género (Scotinella Banks, 1911) y 11 especies son citados por primera vez para México; 13 géneros y 18 especies son nuevos registros para Baja California Sur (Anexo). Asimismo, se ilustra por primera vez al género Filistatoides F. O. P. Cambridge, 1899 (figs. 4f y 5a), citado por Jiménez (2004) como primer registro para México. Las familias con mayor número de géneros fueron Araneidae (17), Theridiidae (16), Salticidae (15) y Gnaphosidae (13), las 2 primeras tejedoras de redes orbiculares y redes espaciales, respectivamente, y las 2 últimas cazadoras o arañas errantes (Cardoso, Pekár, Jocqué y Coddington, 2011). Las familias con mayor riqueza de especies fueron Salticidae (36), Araneidae (29), Theridiidae (27), Gnaphosidae (24) y Corinnidae (15), lo que muestra que las saltadoras, tejedoras de redes orbiculares y espaciales, así como las corredoras son dominantes, tal como se ha registrado en la mayoría de los ecosistemas a nivel mundial (Cardoso et al., 2011). En contraste, las familias que registraron menor riqueza fueron Ctenidae, Ctenizidae, Cyrthaucheniidae, Dipluridae, Homalonychidae, Pisauridae, Theraphosidae y Uloboridae, cada una con un género y una sola especie (fig. 2).
 Scotinella sp.♂ habitus; b) pedipalpo vista prolateral; c) epigineo vista ventral; d) Sarinda cutleri ♀ habitus; e) epigineo vista ventral; f) Filistatoides sp. ♂ pedipalpo vista retrolateral.")
 Filistatoides sp. ♂ pedipalpo vista prolateral; b) Heser nilicola ♂ habitus; c) pedipalpo vista prolateral; d) Erigone barrowsi ♂ pedipalpo vista ventral; e) Idionella nesiotes ♂ habitus; f) pedipalpo vista prolateral.")
De acuerdo con lo registrado hasta el momento, los siguientes taxones, encontrados en los oasis de estudio, son considerados como endémicos para la península: Rothilena Maya-Morales y Jiménez, 2013 (Agelenidae) con sus 6 especies: R. griswoldi Maya-Morales y Jiménez, 2013; R. cochimi Maya-Morales y Jiménez, 2013; R. pilar Maya-Morales y Jiménez, 2013; R. golondrina Maya-Morales y Jiménez, 2013; R. sudcaliforniensis Maya-Morales y Jiménez, 2013; R. naranjensis Maya-Morales y Jiménez, 2013. Y 13 especies más: Creugas comondensis Jiménez, 2007; C. guaycura Jiménez, 2008 (Corinnidae); Filistatinella palaciosi Jiménez y Palacios-Cardiel, 2012 (Filistatidae); Hogna persimilis Banks, 1898; Pardosa sierra Banks, 1898 (Lycosidae); Oecobius hoffmannae Jiménez y Llinas, 2005 (Oecobiidae); Plectreurys arida Gertsch, 1958; P. bicolor Banks, 1898 (Plectreuridae); Lyssomanes pescadero Jiménez y Tejas, 1993 (Salticidae); Loxosceles baja Gertsch y Ennik, 1983; L. mulege Gertsch y Ennik, 1983 (Sicariidae); Misumenoides quetzaltocatl Jiménez, 1992 y Tmarus ehecatltocatl Jiménez, 1992 (Thomisidae).
Como nuevos registros a nivel genérico para Baja California Sur tenemos a Sarinda Peckham y Peckham, 1892 (García-Villafuente, 2009;) (Salticidae); Septentrina Bonaldo, 2000 (Corinnidae) (Bonaldo, 2000); Grammonota Emerton, 1881 (Banks, 1898; Bishop y Crosby, 1932; Ibarra-Núñez, Maya-Morales y Chamé-Vázquez, 2011); Erigone Audouin, 1826 (Gertsch y Davis, 1937); Idionella Banks, 1893 (Dondale, 1959; Ivie, 1967) (Linyphiidae); Pholcophora Banks, 1896 (Gertsch, 1977, 1982) (Pholcidae); Ummidia Thorell, 1875 (Hoffman, 1976; Ibarra-Núñez et al., 2011) (Ctenizidae); Coleosoma O. P. Cambridge, 1882 (Levi, 1959a, Hoffman, 1976); Chrosiothes, Simón, 1894 (Levi, 1964); Cryptachaea Archer, 1946 (Levi, 1955); Dipoena Thorell, 1869 (Levi, 1953a, b, 1963a); Thymoites Keyserling, 1884 (Levi, 1959b, Hoffman, 1976, Ibarra-Núñez et al., 2011); Hentziectypus Archer, 1946 (Levi,1959b) (Theridiidae); y Corythalia C. L. Koch, 1850 (Richman y Cutler, 2012) (Salticidae).
En cuanto a los nuevos registros de especies para Baja California Sur, documentamos a Metepeira arizonica Chamberlin e Ivie, 1942; M. minima Gertsch, 1936 (Piel, 2001); Neoscona utahana (Chamberlin, 1924) (Berman y Levi, 1971) (Araneidae); Castianeira similis (Banks, 1929); C. venusta (Banks,1898) (Reiskind, 1969) Meriola decepta Banks, 1895 (Platnick y Shadab, 1974) (Corinnidae); Drassyllus conformans Chamberlin, 1936; D. notonus Chamberlin, 1928 (Platnick y Shadab, 1982); Synaphosus syntheticus (Chamberlin, 1924) (Ovtsharenko, Levy y Platnick, 1994) (Gnaphosidae); Sassacus papenhoei, Peckham y Peckham, 1895 (Richman, 2008); Habronattus elegans (Peckham y Peckham, 1901); H. hallani (Gertsch, 1934) (Griswold, 1987); Corythalia penicillata (F. O. P. Cambridge, 1901) (Hoffman, 1976) (Salticidae); Cryptachae porteri (Banks, 1896) (Levi, 1959b); Theridion myersi Levi, 1957 (Levi, 1957); Wamba crispulus (Simon, 1895) (Levi, 1963b); Hentziectypus schullei (Gertsch y Mulaik, 1936) (Levi, 1955) (Theridiidae); y Xysticus lutzi Gertsch, 1935 (Schick, 1965) (Thomisidae).
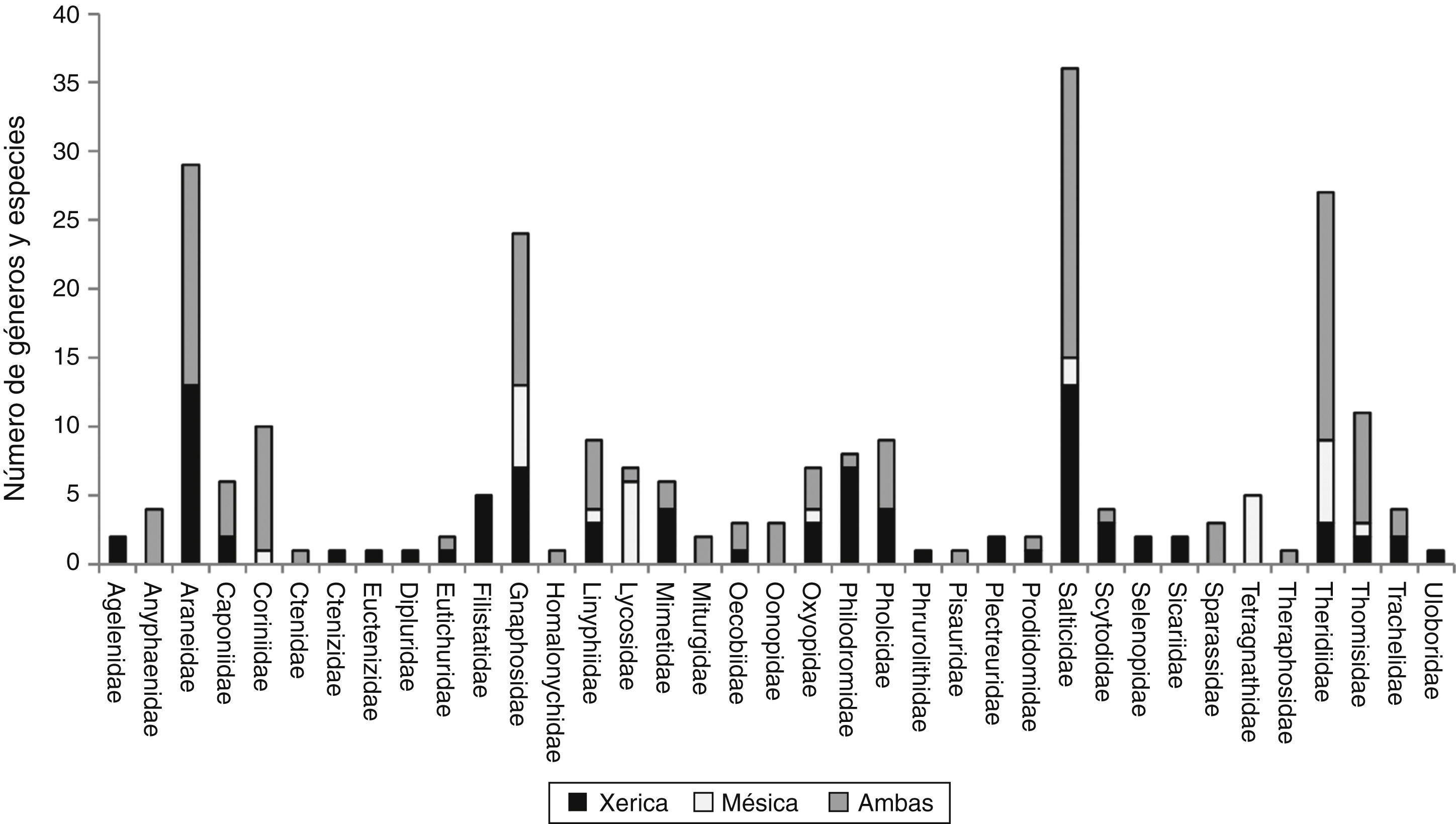
En el ambiente xérico se encontró un total de 215 especies, 125 géneros y 32 familias, y en el ambiente mésico se registraron 157 especies, 96 géneros y 26 familias; ambos ambientes compartieron 128 especies (fig. 3), siendo su complementariedad del 47.5%. En el estrato del suelo se registraron 144 especies, 89 géneros, 28 familias y en el estrato de vegetación 119 especies, 45 géneros y 16 familias, ambos estratos compartieron 19 especies, 125 especies se registraron solo del suelo y 100 de vegetación (Anexo), registrando una complementariedad del 92.2%.
DiscusiónCon este trabajo se incrementa cuantiosamente el conocimiento sobre la biodiversidad de arañas para México y para el estado de Baja California Sur, notándose lo mucho que falta por conocer de la misma, por el hecho de que un porcentaje considerable de la fauna se registra por primera vez, siendo la mayoría previamente documentada para EE. UU. y Canadá. Por ejemplo, a nivel genérico Scotinella Banks, 1911 (fig. 4a, b y c) citado para Canadá y Estados Unidos; asimismo, ratificamos la presencia de Sarinda Peckham y Peckham, 1892 (fig. 4d, e) registrado por García-Villafuente (2009), debido a que en el catálogo mundial de arañas (World Spider Catalog, 2015) no ha sido registrado para México, a pesar de que este género es conocido desde EE. UU. hasta Argentina. Los taxones Erigone barrowsi Crosby y Bishop, 1928 (fig. 5d), Idionella nesiotes (Crosby, 1924) (fig. 5e, f), Habronattus californicus (Banks, 1904) (fig. 6a, b), Sarinda cutleri (Richman, 1965) (fig. 4d, e), Emertonella emertoni (Bryant, 1933) (fig. 6c, d, e, f) y Euryopis mulaiki Levi, 1954 (fig. 7a, b), igualmente se registran solo para EE. UU.. Por su parte, Tigrosa helluo (Walckenaer, 1837) (fig. 7e), Dipoena nigra (Emerton, 1882) (fig. 7c, d) y Uloborus glomosus (Walckenaer, 1841) (fig. 7f) se citan para EE. UU. y Canadá, mientras que Theridion varians Hohn, 1823 tiene distribución Holártica y en América se registra solo en EE. UU. y Canadá. A la par destaca Heser nilicola (O. P. Cambridge, 1874) (fig. 5b, c), especie propia del Mediterráneo, las Islas Canarias, Burkina Faso y, aunque documentada para EE. UU., es considerada como introducida en América (Platnick y Shadab, 1983; World Spider Catalog, 2015).
 Habronattus californicus ♂ habitus; b) pedipalpo vista prolateral; c) Emertonella emertoni ♂ habitus vista lateral; d) habitus vista dorsal; e) pedipalpo vista retrolateral; f) epigineo vista ventral.")
 Euryopis mulaiki ♂ habitus; b) pedipalpo vista prolateral; c) Dipoena nigra ♂ habitus, d) pedipalpo vista prolateral; e) Tigrosa helluo ♂ pedipalpo vista ventral; f) Uloborus glomosus ♀ epigineo vista ventral.")
Para Baja California Sur, los 13 taxones nuevos registros a nivel genérico habían sido registrados previamente para 24 estados de la República, siendo Chiapas, Colima, Guerrero, Hidalgo, Nayarit, San Luis Potosí, Tabasco, Tamaulipas y Veracruz donde se distribuye cerca del 60% de estos registros para el estado. Concomitantemente, los 18 nuevos registros de especies para Baja California Sur también habían sido registrados en un 60% para Baja California, Chihuahua, Guerrero, Jalisco, Nuevo León, Oaxaca, San Luis Potosí, Sinaloa, Sonora y Tamaulipas. De acuerdo con Jiménez y Palacios-Cardiel (2012), el número de especies conocidas para el estado era de 208, y con este estudio se incrementó en un 8.7%. Es probable que esta gran diversidad esté relacionada con la evolución geomorfológica de la península de Baja California y de la región noroeste de México que produjo una transformación radical de un hábitat con vegetación mésica subtropical a un matorral xerófilo, generando hábitats únicos con gran variabilidad de microambientes (Arriaga, 1997; Axelrod, 1979). Esta transformación como resultado de millones de años, sin lugar a duda, creó los actuales refugios mésicos que hoy solo se encuentran en las porciones más altas de las cadenas montañosas, o en los oasis o humedales en medio del desierto (Arriaga, 1997; Grismer y McGuire, 1993). Es por ello que los nuevos registros de taxones de arañas de este trabajo también se distribuyan en otras entidades del interior de la república, donde la temperatura ambiental sea más fría y la precipitación mayor que en las zonas desérticas.
Por otra parte, en este trabajo se encontró una elevada proporción de especies no descritas (24.5%), destacando las de los géneros Mimetus (Mimetidae), Pholcophora (Pholcidae) y Scytodes (Scytodidae). Esto indica que existe una gran riqueza específica que falta por conocer y describir, sobre todo, en algunos grupos que pueden incrementar el porcentaje de endemismos conocido, como el caso del género Rothilena (Agelenidae), recientemente descrito para la península (Maya-Morales y Jiménez, 2013), además de las 13 especies citadas anteriormente. Otro aspecto importante es que la riqueza de especies en los oasis se incrementó en un 75% de lo registrado previamente por Llinas-Gutiérrez y Jiménez (2004). Este porcentaje también es mayor si lo comparamos con estudios llevados a cabo en otras zonas áridas de Norteamérica, como en el matorral desértico del desierto sonorense en Arizona, en el que Chew (1961) registró 4 familias, 12 géneros y 20 especies de arañas asociadas al follaje de Larrea divaricata. En el matorral del desierto chihuahuense Hatley y MacMahon (1980) recolectaron 40 especies de 11 familias en el cañón Verde en Cache Country en Utha; Bowen, Horner y Cook (2004) confirmaron 26 familias, siendo las Gnaphosidae las más diversas con 11 géneros y 26 especies; Broussard y Horner (2006) registraron un total de 24 familias, 46 géneros y 66 especies de arañas cursoriales en el Trans-Pecos al oeste de Texas; Lightfoot, Brantley y Allen (2008) encontraron 121 especies de arañas en el suelo en Nuevo México; y Richman, Brantley Hu y Whitehouse (2011) registraron un total de 24 familias y 117 especies de arañas recolectadas durante 30 años en la vegetación desértica de esta región. Probablemente la gran diferencia en la riqueza de especies encontrada en este trabajo comparada con otras zonas desérticas, sea porque se incluyeron especies de suelo y vegetación, mientras que en los trabajos mencionados solo incluyeron especies de un único estrato.
La alta riqueza de géneros y especies de arañas tejedoras de red de las familias Araneidae (16/29) y Theridiidae (16/27), tal como se esperaba, se debió a que fueron capturadas tanto en el follaje de la vegetación xerófila como en la mésica de los oasis y es muy semejante a lo registrado para el bosque mesófilo de montaña (Ibarra-Núñez et al., 2011) y las zonas tropicales (Silva y Coddington, 1996), pero difiere de lo obtenido por el trabajo de Richman, Brantley, Hu y Whitehouse (2011) para el desierto chihuahuense, donde estas familias estuvieron representadas por pocos géneros y especies (Araneidae 5/7, Theridiidae 6/11). Es probable que en nuestro caso, la heterogeneidad florística, disimilitud estructural y tipos de vegetación entre los oasis distribuidos en diferentes latitudes de Baja California Sur, y la vegetación circundante de matorral xerófilo (Arriaga, Díaz, Domínguez y León, 1997) hayan permitido el exitoso establecimiento de diversas familias de arañas, debido a la presencia de especies vegetales arbóreas y arbustivas como especies de Acacia sp. (huizache), Parkinsonia florida subsp. peninsulare (palo verde), Jatropha cinerea (lomboy), Leucaena microcarpa, Olneya tesota (palo fierro), Chloroleucon mangense var. leucospermum (palo ébano) y Prosopis articulata (mezquite), entre otras, así como especies mesófilas como Salix sp., que junto con las palmas Washingtonia robusta y Phoenix dactylifera y huertos de frutales, han favorecido la creación de ambientes más húmedos en medio del desierto (Arriaga et al., 1997). Estos ambientes tienen una estructura vegetal compleja que pueden ofrecer una gran variedad de microhábitats disponibles para el establecimiento de las arañas. En el caso de las arañas cazadoras errantes, Salticidae y Gnaphosidae también tuvieron alta riqueza en los oasis. Sin embargo, estas familias también han sido registradas como muy ricas en una localidad del matorral desértico del tipo sonorense al sur de la península (Jiménez y Navarrete, 2010) y en el desierto chihuahuense (Broussard y Horner, 2006; Richman et al., 2011). Esto podría indicar que sus especies podrían ser muy tolerantes ecológicamente al encontrarse tanto en el ambiente xérico como en el mésico (fig. 3).
Los valores de complementariedad de la comunidad de arañas del matorral xerófilo y de zonas mésicas (47.5%) indican que estas comunidades son muy distintas, sobre todo, considerando que en el gradiente xérico-mésico las familias Agelenidae, Ctenizidae, Cyrthaucheniidae, Dipluridae, Filistatidae, Plectreuridae, Selenopidae, Sicariidae y Uloboridae, se circunscriben solo al matorral xerófilo probablemente porque están mejor adaptadas a las condiciones secas y extremosas del ambiente árido, estando ausentes en la vegetación mésica (fig. 2). En cambio, en el ambiente húmedo, 4 especies de Lycosidae y 5 de Tetragnathidae fueron registradas solamente en la vegetación mesófila, que se desarrolla en estos ambientes generados por la presencia de los cuerpos de agua. Las 127 especies restantes que estuvieron presentes a todo lo largo del gradiente xérico-mésico de los oasis estudiados, posiblemente son especies euritópicas (con amplios rangos de tolerancia ambiental) que pueden transitar o permanecer en ambos ambientes (fig. 2). En el estrato del suelo se encontró un mayor número de especies (144) y familias (29) que en la vegetación (119 y 18, respectivamente) debido a la alta riqueza de Gnaphosidae, Linyphiidae y Lycosidae. En cuanto a la familia Salticidae (arañas errantes) y Theridiidae (arañas tejedoras de redes) fueron también muy diversas en ambos estratos con una riqueza similar de especies. En cuanto a los índices de complementariedad de los estratos, en estudios previos realizados en el bosque mesófilo de montaña Sorensen (2003) registró un valor del 58.1% e Ibarra-Núñez et al. (2011) obtuvieron un 76.2%, mientras que en este trabajo se registró un 92.2%. Esto puede demostrar que el estrato de vegetación comparte pocas especies con el estrato del suelo y que ambos estratos tienen un alto grado de especificidad en el hábitat, debido probablemente a las diferencias en las condiciones ambientales proporcionadas por los mismos.
La gran riqueza encontrada en este estudio nos da un muy buen indicio del éxito de las comunidades de arañas de estos oasis, lo cual está potencialmente relacionado con la coexistencia de diferentes gremios tróficos, que como grupo les permite ocupar y tolerar una gran variedad de microhábitats en el ambiente xérico y mésico, debido a su alta capacidad de dispersión, sus diversas estrategias tróficas, y quizá a que los oasis aún no están tan severamente perturbados por la actividad antropogénica, entre otros muchos otras factores que sin duda se requiere abordar.
A Ibeth Posada y Carlos A. Frieven por su asistencia en la recolecta de material en campo, a Gerardo Hernández del CIBNOR por la edición de las fotografías, a Diana Dorantes por la edición del resumen en inglés y a los revisores anónimos y editor asociado por sus valiosos comentarios y sugerencias al manuscrito. Este proyecto fue apoyado por el Consejo Nacional de Ciencia y Tecnología (Semarnat-Conacyt C01-0052).
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.