








Los patrones de reclutamiento de peces aportan información valiosa para entender la dinámica de sus poblaciones. El presente trabajo analiza variaciones espaciales y temporales en las abundancias de reclutas de 4 peces arrecifales (Stegastes partitus, Chromis cyanea, Gramma loreto y Scarus taeniopterus) en Banco Chinchorro, Caribe mexicano, mediante censos visuales realizados en hábitats de cordillera y parche de febrero de 2006 a febrero de 2007. La evaluación del hábitat se basó en las coberturas del fondo marino registradas en vídeo-transectos y en la complejidad del sustrato. En las especies estudiadas se detectó una tendencia de mayor reclutamiento en junio, julio y agosto. Usando pruebas pareadas de Ansari-Bradley y análisis de redundancia, se determinó que las mayores abundancias de reclutas de Chromis cyanea y Gramma loreto estuvieron asociadas al hábitat de cordillera, el cual presentó valores comparativamente altos de profundidad y complejidad. Por otra parte, Scarus taeniopterus y Stegastes partitus no mostraron una preferencia clara para la selección del hábitat. Aunque se requiere de estudios más amplios en términos de número de hábitats y especies, los resultados de la presente investigación resaltan la importancia de las cordilleras coralinas para el reclutamiento y, consecuentemente, para el mantenimiento de las poblaciones de peces arrecifales.
Fish recruitment patterns provide valuable information for understanding their population dynamics. This paper analyzes temporal and spatial variations in recruits abundance of 4 reef fishes (Stegastes partitus, Chromis cyanea, Gramma loreto and Scarus taeniopterus) at Chinchorro Bank, Mexican Caribbean, by means of visual censuses conducted in spur and patch habitats from February 2006 to February 2007. Habitat assessment was based on sea floor coverage recorded by video transects and on substratum complexity. The species studied showed a tendency towards higher recruitment rates in June, July and August. Using Ansari-Bradley pairwise tests and redundancy analysis, higher abundances of Chromis cyanea and Gramma loreto recruits were found to be associated mainly with spur habitats, which showed relatively high depth and complexity values. On the other hand, Scarus taeniopterus and Stegaste partitus showed no clear preference in terms of habitat selection. Although further studies on more habitats and species are needed, the results of this research highlight the relevance of coral spurs for recruitment and, consequently, for the maintenance of reef fish populations.
Una de las comunidades más diversas en los sistemas arrecifales es la constituida por los peces, cuyo ciclo vital presenta 2 fases: la larval pelágica, la cual les permite dispersarse, y la adulta demersal, relativamente sedentaria (Choat y Bellwood, 1991). En las poblaciones marinas, la adición de individuos a una población local generalmente ocurre entre la fase larval pelágica y la fase juvenil temprana béntica o demersal, después del asentamiento. El estudio de este proceso poblacional, denominado reclutamiento, es clave para entender un amplio rango de fenómenos ecológicos, desde la dinámica demográfica hasta la estructura genética de las metapoblaciones (Caley et al., 1996). Los índices de reclutamiento tienden a ser muy irregulares tanto temporal como espacialmente (Hogan, Thiessen, Sale y Heath, 2012; Paddack y Sponaugle, 2008; Vallès, Kramer y Hunte, 2008), por lo cual, en algunas ocasiones es difícil establecer claramente patrones de variabilidad. Entre los principales factores asociados con la abundancia y distribución de nuevos reclutas en los arrecifes se han identificado la abundancia de coespecíficos Anderson, Carr y Hixon, 2007; Webster, 2003), las tasas diferenciales de sobrevivencia después del asentamiento (Carr y Hixon, 1995; Wellington, 1992) y los atributos del hábitat (Booth y Beretta, 1994; Paddack y Sponaugle, 2008; Tolimieri, 1995, 1998a). En cuanto a las variaciones temporales, estudios previos realizados en arrecifes del Caribe no han detectado un patrón claro de estacionalidad (Robertson, Schober y Brawn, 1993; Sponaugle y Cowen, 1996). En aquellos casos en los que los patrones de reclutamiento se explican en buena medida con base en las características del hábitat (Holbrook, Forrester y Schmitt, 2000; Risk, 1997; Stier y Osenberg, 2010; Tolimieri, 1998a, 1998b), el sustrato arrecifal puede concebirse como una compleja matriz en la cual los peces encuentran alimento y refugio contra los depredadores. Por consiguiente, resulta razonable que diferentes tipos de hábitats provean diversos grados de protección y disponibilidad de nutrientes, con mayor abundancia de reclutas en hábitats «preferenciales», lo cual se manifestaría de diferentes maneras de acuerdo con la escala espacial de cada estudio (Caselle y Warner, 1996; Tolimieri, 1995, 1998a, 1998b). Conocer la relación entre los diversos tipos de hábitats y las tasas de reclutamiento de las especies ícticas que habitan los arrecifes es un aspecto particularmente importante para prever cambios en las dinámicas poblacionales producidos por alteraciones en los hábitats de zonas coralinas.
Algunos autores han propuesto que las oscilaciones en los patrones de asentamiento y reclutamiento de peces arrecifales pueden tener un fuerte componente estocástico (Doherty, 2002; Tolimieri, 1995), mientras que otros plantean que podrían deberse a cambios en ciertas conductas reproductivas relacionadas con ciclos anuales o estacionales (Ault y Johnson, 1998; Bouchon-Navaro, Louis y Bouchon, 1997), e incluso se las ha asociado a ciclos de mayor influencia en la conducta social de las especies, tales como el diario y el lunar (Galzin, 1987; Shapiro, 1991). Según algunos autores, lo más probable es que la abundancia local esté determinada tanto por las tasas de reclutamiento como por las interacciones locales, aunque la magnitud del efecto de cada uno de estos factores aún no es del todo clara (Chesson, 1998; Pitchford, James y Brindley, 2005).
En el mar Caribe se encuentra la segunda barrera arrecifal más grande del mundo, cuya porción norte está ubicada frente a las costas del estado mexicano de Quintana Roo. Banco Chinchorro se encuentra entre los sistemas arrecifales más importantes de la región; en 1996 fue declarado Reserva de la Biosfera debido a su diversidad de flora y fauna (SEMARNAT, 1996). Para el adecuado manejo de esta, como de toda área marina protegida, es preciso contar con una robusta base científica en cuanto a las dinámicas de reclutamiento de sus comunidades ícticas, que permitan identificar sitios y/o temporadas prioritarias para enfocar adecuadamente los esfuerzos de conservación. No obstante, en la actualidad no hay estudios publicados sobre el reclutamiento de los peces de Banco Chinchorro, como tampoco los hay con respecto a otros arrecifes del Caribe mexicano. Por ello, la presente investigación se planteó como objetivo analizar las variaciones tanto espaciales como temporales, en el reclutamiento de 4 especies ícticas representativas de los sistemas arrecifales de la región: Chromis cyanea (C. cyanea), Gramma loreto (G. loreto), Scarus taeniopterus (S. taeniopterus) y Stegastes partitus (S. partitus), en sitios localizados en 2 tipos de hábitats de la Reserva de la Biosfera Banco Chinchorro.
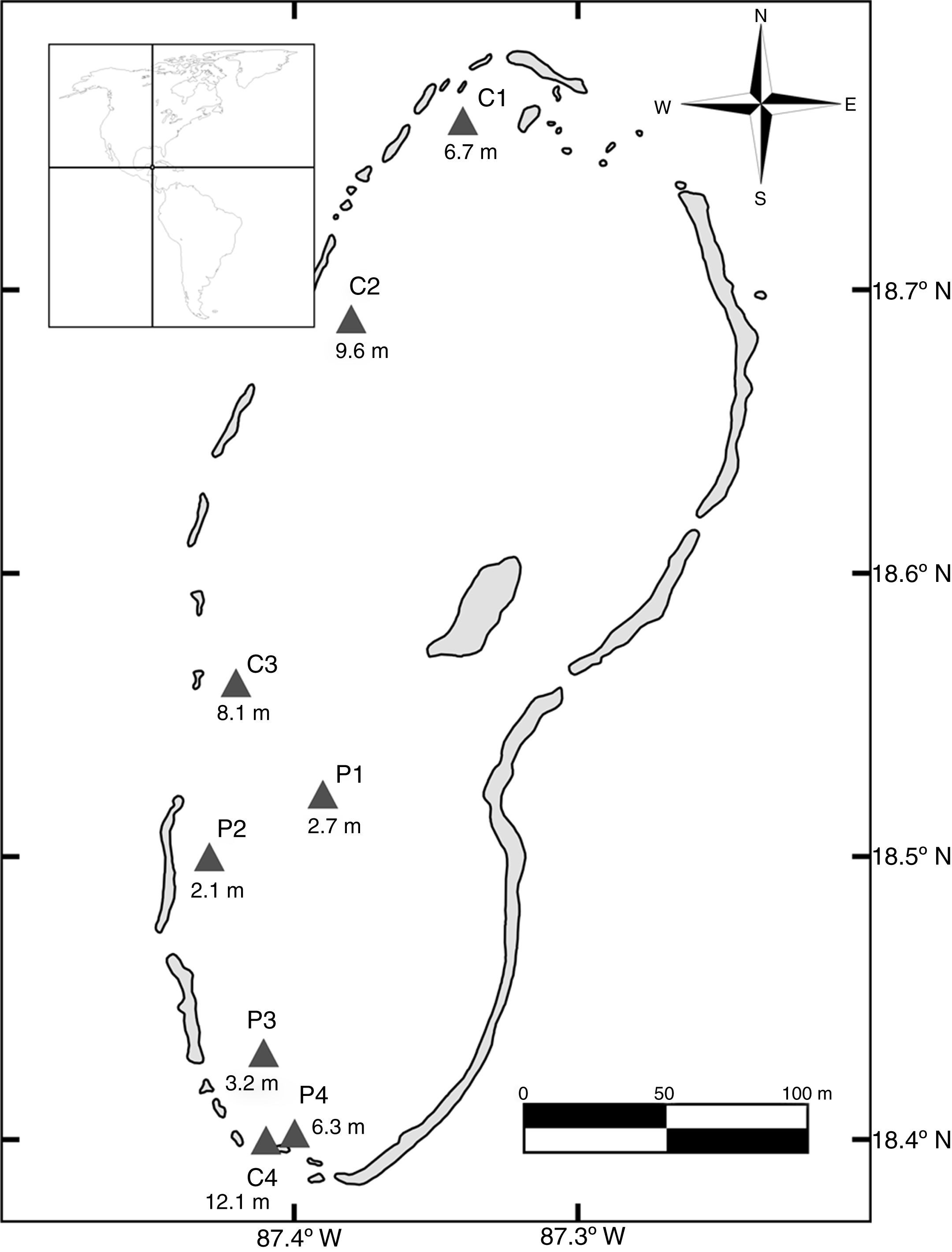
Materiales y métodosBanco Chinchorro es un arrecife tipo plataforma (Chávez e Hidalgo, 1988), con un área total de 814.2km2 y un gradiente batimétrico que va desde 1m en la parte norte hasta 25m en la parte sur (González, Torruco, Liceaga y Ordaz, 2003); se ubica a 30.8km de Mahahual y está separado de la costa por un canal de 1,000m de profundidad máxima (Wells, 1988). Geográficamente se localiza en las coordenadas 18°35’ N, 87°22’ O (fig. 1). Este sistema está rodeado por una barrera arrecifal de forma elíptica irregular cuyo perímetro es de 115km (Jordán y Martín, 1987). La laguna arrecifal tiene una superficie de aproximadamente 533km2 (González et al., 2003), con diversos tipos de hábitats que van desde cordilleras y macizos hasta parches coralinos y arenales (INE-SEMARNAT, 2000).

Las 4 especies ícticas estudiadas (C. cyanea, G. loreto, S. taeniopterus y S. partitus) son típicas de zonas arrecifales. Varias características hacen de estas especies buenos modelos de estudio: se las ha registrado como dominantes numéricamente en el Sistema Arrecifal Mesoamericano, no presentan comportamiento críptico y sus reclutas son fácilmente identificables bajo el agua atendiendo aspectos específicos de talla y coloración. Para tener la certeza de incluir solo organismos reclutados durante los meses de muestreo, con base en la información publicada sobre las tallas máximas de reclutas se censaron únicamente individuos menores a 3.5cm para los casos de C. cyanea y S. taeniopterus, y menores a 3.0 y 2.5cm para los casos de G. loreto y S. partitus, respectivamente (Almada-Villela, Sale, Gold-Bouchot y Kjerfve, 2003). Para la adecuada identificación de los reclutas de S. taeniopterus se tomó en cuenta que estos presentan una coloración diferente a la de los adultos; además, la distinción de esta especie respecto de otras similares se basó en la presencia de bordes oscuros en la aleta caudal (Humann y DeLoach, 2002).
Los muestreos se llevaron a cabo en 8 sitios de sotavento seleccionados con base en su desarrollo coralino, 4 en hábitats de cordilleras (C1, C2, C3, C4) y 4 en hábitats de parche (P1, P2, P3, P4; fig. 1). La metodología utilizada para el muestreo de datos de reclutas se basó en censos visuales con equipo scuba y transectos en banda de 30m de largo por 1m de ancho (Almada-Villela et al., 2003); este método es el recomendado cuando se desea obtener estimaciones cuantitativas de las abundancias (Sanderson y Solonsky, 1986). En cada uno de los sitios se realizaron 6 réplicas separadas lateralmente entre sí por una distancia mínima de 5m (Almada-Villela et al., 2003; English, Wilkinson y Baker, 1997). Los censos se realizaron entre las 9:00 y las 16:00h, durante fechas próximas al plenilunio, ya que en el asentamiento de varias especies se ha observado un pico en torno a esta fase lunar (Robertson, 1992). Los datos de riqueza y abundancia se registraron en hojas de acrílico; adicionalmente, se registró la profundidad y la temperatura de cada sitio. Temporalmente los muestreos se realizaron durante 9 meses a lo largo de un ciclo anual (feb-06, abr-06, jun-06, jul-06, ago-06, oct-06, nov-06, ene-07, feb-07).
Se estimaron los porcentajes de coberturas bénticas mediante la técnica de vídeo-transectos (Aronson, Edmunds, Precht, Swanson y Levitan, 1994; Aronson y Swanson, 1997), los cuales fueron filmados usando como guía los transectos en banda trazados para los censos, por lo que en cada sitio se grabaron en total 6 vídeo-transectos, cada uno abarcando un área del sustrato de 30m de longitud por aproximadamente 0.5m de ancho. El análisis de los vídeos se llevó a cabo seleccionando 40 fotogramas distribuidos a intervalos de tiempo iguales a la duración del vídeo dividida por 40. En cada fotograma se identificó el tipo de sustrato debajo de 13 puntos distribuidos uniformemente en un monitor, según el método –modificado– de Aronson et al. (1994). De esta manera, en cada vídeo se identificaron los sustratos correspondientes a un total de 520 puntos, lo cual constituye un muestreo suficiente para obtener caracterizaciones firmes de cualquier ambiente arrecifal (Hernández-Landa, 2004). Las categorías que se utilizaron para agrupar los tipos de coberturas bénticas fueron: arena, algas, esponjas, rocas y escombro, y 3 grupos coralinos (hexacorales, hidrocorales y octocorales), con base en el trabajo de Rodríguez-Zaragoza (2007).
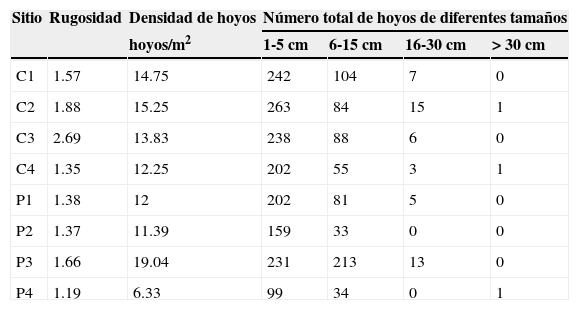
Para evaluar la complejidad del sustrato se utilizó la rugosidad, así como el número y diámetro de los hoyos. La rugosidad fue estimada utilizando una cadena delgada de acero de 10m de longitud, la cual fue colocada al lado de cada transecto a los 0, 10 y 20m, siguiendo el relieve del sustrato. En cada caso, se determinó el índice de rugosidad dividiendo la longitud total de la cadena (10m) entre la longitud real, es decir, entre la distancia ocupada por dicha cadena extendida sobre el sustrato (Luckhurst y Luckhurst, 1978). En cada sitio se obtuvieron 18 medidas de rugosidad. El número y diámetro de los hoyos en el sustrato marino fue evaluado mediante cuadrantes de 1m2, ubicados a 0, 10, 20 y 30m sobre los transectos, para tener un total de 24 cuadrantes por sitio. Las categorías para la clasificación de los hoyos con base en sus diámetros se establecieron de acuerdo con Roberts y Ormond (1987), agrupando las 2 últimas categorías, de modo que se utilizaron 4 categorías: 1-5, 6-15, 16-30 y>30cm.
Análisis estadísticosSe calculó la abundancia promedio y densidad (individuos/m2) en cada uno de los sitios y meses muestreados. Dado que los datos obtenidos no fueron paramétricos y que el modelo de muestreo resultó ser no balanceado, se utilizaron pruebas pareadas de Ansari-Bradley tanto para el contraste de abundancias entre los hábitats de cordillera y parche como entre los diversos sitios y meses, con el propósito de identificar diferencias significativas específicas (Ansari y Bradley, 1960; Hollander y Wolfe, 1973). Para los casos en los que fue necesario, las medianas fueron igualadas de acuerdo con lo descrito por Sprent (1989). Las pruebas se llevaron a cabo mediante la función ansari.test integrada en el paquete Stats versión 2.15.3 para el programa R (Ihaka y Gentleman, 1996). Debido a que la matriz correspondiente a S. taeniopterus presentó una cantidad de ceros comparativamente alta en los sitios y meses estudiados, no se incluyeron los datos de esta especie en las pruebas pareadas de Ansari-Bradley para evitar resultados no confiables.
Para detectar si uno o más factores ambientales explican de forma significativa la variación observada en las abundancias, se realizaron análisis multivariables basados en distancias para modelos lineales usando el programa DISTLM versión 5.0 (Anderson, 2004). Tanto para los análisis espaciales como para los temporales se usaron los datos brutos sin transformar ni estandarizar, con valores de ji-cuadrado como medidas de distancia y 9,999 permutaciones. Mediante un análisis de ordenación se evaluó la relación de la abundancia de los reclutas con las coberturas bénticas y las variables ambientales. Previamente se llevó a cabo un análisis de correspondencia sin tendencia usando el programa CANOCO versión 4.0 (Leps y Smilauer, 2003). Como los resultados de dicho análisis indicaron que la longitud máxima de los gradientes fue menor a 3, mediante el programa mencionado se llevó a cabo un análisis de redundancia (RDA; Ter Braak y Smilauer, 2002). Debido a que los análisis de RDA llevados a cabo con los datos originales presentaron altos factores de inflación, se decidió eliminar 2 variables, una de cada grupo de datos: de los porcentajes de coberturas bénticas se eliminó el grupo de las algas, y de las variables de complejidad del sustrato se eliminó la categoría de hoyos mayores a 30cm.
ResultadosSe registraron en total 3,440 reclutas en el área de estudio. Las mayores densidades se registraron en los sitios con cordilleras (fig. 2). Tanto C. cyanea como G. loreto registraron mayores abundancias promedio en el hábitat de cordillera, mientras que las otras 2 especies no mostraron una clara preferencia de hábitat (fig. 3). Los reclutas de G. loreto y S. partitus se observaron en todos los sitios; C. cyanea estuvo presente en casi la totalidad de los sitios, con excepción de P2. En contraparte, S. taeniopterus se registró únicamente en 2 sitios (P3 y C3; fig. 3). Con las pruebas de Ansari-Bradley entre hábitats se observó que tanto C. cyanea como G. loreto presentaron niveles de reclutamiento significativamente diferentes entre ambos hábitats (p<0.01), mientras que S. partitus no presentó diferencias en este sentido. Las pruebas pareadas a nivel de sitios en ninguna de las 3 especies revelaron un patrón claro.
 de cada especie en los sitios de muestreo. C: cordillera; P: parche.")

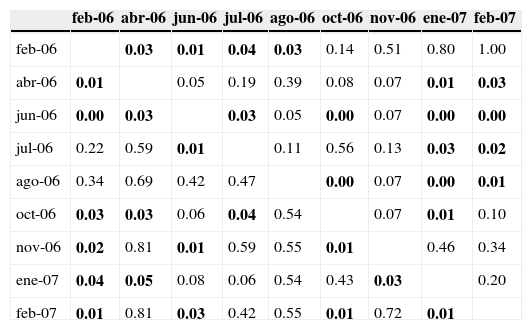
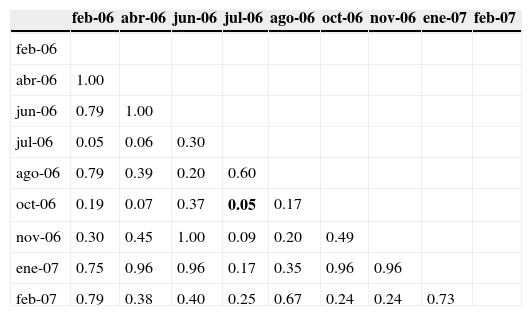
El valor más alto de densidad promedio de reclutas se observó en el mes de agosto, y en noviembre se registró el más bajo (fig. 4); para las 4 especies el reclutamiento máximo correspondió a alguno de los meses de verano, entre junio y agosto (fig. 5). Mientras que G. loreto y S. partitus fueron reclutadas durante todo el año, C. cyanea y S. taeniopterus no lo fueron, ya que la primera no fue registrada en noviembre, y la segunda solo se registró en 4 de los 9 meses. No obstante, en los resultados de las comparaciones pareadas de abundancias entre meses para C. cyanea, G. loreto y S. partitus no se aprecia un patrón significativo de estacionalidad (tablas 1 y 2).
 para las 4 especies durante los meses de muestreo.")

Resultados de las pruebas pareadas de Ansari-Bradley entre meses.
| feb-06 | abr-06 | jun-06 | jul-06 | ago-06 | oct-06 | nov-06 | ene-07 | feb-07 | |
|---|---|---|---|---|---|---|---|---|---|
| feb-06 | 0.03 | 0.01 | 0.04 | 0.03 | 0.14 | 0.51 | 0.80 | 1.00 | |
| abr-06 | 0.01 | 0.05 | 0.19 | 0.39 | 0.08 | 0.07 | 0.01 | 0.03 | |
| jun-06 | 0.00 | 0.03 | 0.03 | 0.05 | 0.00 | 0.07 | 0.00 | 0.00 | |
| jul-06 | 0.22 | 0.59 | 0.01 | 0.11 | 0.56 | 0.13 | 0.03 | 0.02 | |
| ago-06 | 0.34 | 0.69 | 0.42 | 0.47 | 0.00 | 0.07 | 0.00 | 0.01 | |
| oct-06 | 0.03 | 0.03 | 0.06 | 0.04 | 0.54 | 0.07 | 0.01 | 0.10 | |
| nov-06 | 0.02 | 0.81 | 0.01 | 0.59 | 0.55 | 0.01 | 0.46 | 0.34 | |
| ene-07 | 0.04 | 0.05 | 0.08 | 0.06 | 0.54 | 0.43 | 0.03 | 0.20 | |
| feb-07 | 0.01 | 0.81 | 0.03 | 0.42 | 0.55 | 0.01 | 0.72 | 0.01 |
Arriba de la diagonal se muestran las probabilidades relativas a la especie C. cyanea, y abajo de la diagonal, para la especie G. loreto. Con negritas se presentan los valores significativos (p<0.05).
Resultados de las pruebas pareadas de Ansari-Bradley entre meses. Probabilidades correspondientes a la especie S. partitus.
| feb-06 | abr-06 | jun-06 | jul-06 | ago-06 | oct-06 | nov-06 | ene-07 | feb-07 | |
|---|---|---|---|---|---|---|---|---|---|
| feb-06 | |||||||||
| abr-06 | 1.00 | ||||||||
| jun-06 | 0.79 | 1.00 | |||||||
| jul-06 | 0.05 | 0.06 | 0.30 | ||||||
| ago-06 | 0.79 | 0.39 | 0.20 | 0.60 | |||||
| oct-06 | 0.19 | 0.07 | 0.37 | 0.05 | 0.17 | ||||
| nov-06 | 0.30 | 0.45 | 1.00 | 0.09 | 0.20 | 0.49 | |||
| ene-07 | 0.75 | 0.96 | 0.96 | 0.17 | 0.35 | 0.96 | 0.96 | ||
| feb-07 | 0.79 | 0.38 | 0.40 | 0.25 | 0.67 | 0.24 | 0.24 | 0.73 |
Con negritas se presentan los valores significativos (p<0.05).
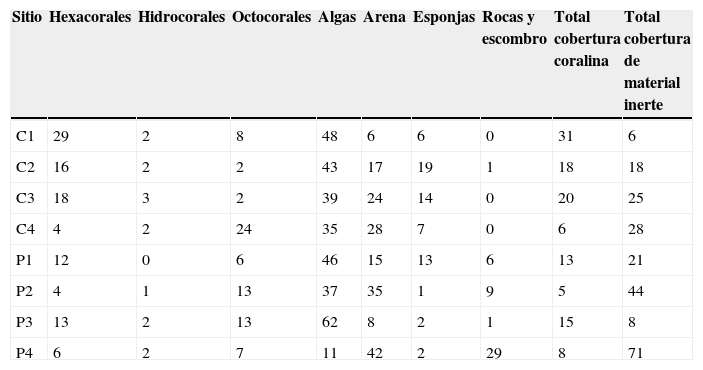
Se analizó un área total de aproximadamente 720m2 de fondo oceánico durante todo el ciclo. En promedio, el hábitat de parche presentó una profundidad de 3.6±1.87m, una rugosidad de 1.4±0.23, y 12.2±6.95 hoyos, con 5 a 15% de coberturas coralinas (fig. 1; tablas 3 y 4). Comparativamente, el hábitat de cordillera mostró mayores valores de profundidad (9.2±2.35m), rugosidad (1.8±0.49) y número de hoyos (14.0±4.18), así como de coberturas coralinas (6 a 31%; fig. 1; tablas 3 y 4). Los mayores porcentajes de cobertura béntica en 7 de los 8 sitios correspondieron al grupo de las algas (con coberturas de entre el 35 y el 62%). Los hidrocorales, así como el sustrato formado por rocas y escombro, registraron generalmente las menores coberturas.
Porcentajes promedio redondeados de las coberturas bénticas en los 8 sitios.
| Sitio | Hexacorales | Hidrocorales | Octocorales | Algas | Arena | Esponjas | Rocas y escombro | Total cobertura coralina | Total cobertura de material inerte |
|---|---|---|---|---|---|---|---|---|---|
| C1 | 29 | 2 | 8 | 48 | 6 | 6 | 0 | 31 | 6 |
| C2 | 16 | 2 | 2 | 43 | 17 | 19 | 1 | 18 | 18 |
| C3 | 18 | 3 | 2 | 39 | 24 | 14 | 0 | 20 | 25 |
| C4 | 4 | 2 | 24 | 35 | 28 | 7 | 0 | 6 | 28 |
| P1 | 12 | 0 | 6 | 46 | 15 | 13 | 6 | 13 | 21 |
| P2 | 4 | 1 | 13 | 37 | 35 | 1 | 9 | 5 | 44 |
| P3 | 13 | 2 | 13 | 62 | 8 | 2 | 1 | 15 | 8 |
| P4 | 6 | 2 | 7 | 11 | 42 | 2 | 29 | 8 | 71 |
Datos promedio de las variables de complejidad del hábitat para los 8 sitios estudiados.
| Sitio | Rugosidad | Densidad de hoyos | Número total de hoyos de diferentes tamaños | |||
|---|---|---|---|---|---|---|
| hoyos/m2 | 1-5cm | 6-15cm | 16-30cm | >30cm | ||
| C1 | 1.57 | 14.75 | 242 | 104 | 7 | 0 |
| C2 | 1.88 | 15.25 | 263 | 84 | 15 | 1 |
| C3 | 2.69 | 13.83 | 238 | 88 | 6 | 0 |
| C4 | 1.35 | 12.25 | 202 | 55 | 3 | 1 |
| P1 | 1.38 | 12 | 202 | 81 | 5 | 0 |
| P2 | 1.37 | 11.39 | 159 | 33 | 0 | 0 |
| P3 | 1.66 | 19.04 | 231 | 213 | 13 | 0 |
| P4 | 1.19 | 6.33 | 99 | 34 | 0 | 1 |
El análisis de regresión espacial indicó que el conjunto de las variables ambientales explica el 47% de la variación en las abundancias, tomando el factor temporal como covariable (seudo-F=2.98, p<0.01). Por otra parte, en el análisis de regresión temporal, con la variación espacial como covariable, se observó que la variación múltiple de las abundancias no está determinada significativamente por el factor temporal (seudo-F=1.03, p<0.32).
Ninguna de las pruebas aplicadas a los 2 ejes canónicos obtenidos en los RDA mostraron significatividad (p>0.05). Los 2 primeros ejes del RDA que relaciona las abundancias con los porcentajes de coberturas acumularon el 95.80% de la varianza, mientras que en el RDA de las abundancias y la complejidad del sustrato el porcentaje de la varianza acumulado por los 2 primeros ejes fue 95.40%. Las coberturas de hidrocorales, hexacorales y esponjas se asociaron a los sitios con hábitat de cordillera, mientras que las coberturas de las categorías rocas y escombro, y arena, se relacionaron mayormente con los sitios de parche, a excepción de P3 (fig. 6). En el RDA también se aprecia que C. cyanea se relacionó con las esponjas, y S. partitus con sitios donde las coberturas de rocas y escombro fueron altas (fig. 6). El análisis de las variables de complejidad del hábitat muestra que la profundidad es la variable que más influyó en el reclutamiento de las especies. Así, G. loreto y C. cyanea se relacionaron con sitios profundos, mientras que S. partitus y S. taeniopterus mostraron afinidad con sitios someros (fig. 7).

Resultados gráficos del análisis de redundancia basado en las matrices de abundancias de las especies y de coberturas bénticas.
C. cyanea: Chromis cyanea; G. loreto: Gramma loreto; S. taeniopterus: Scarus taeniopterus; S. partitus: Stegastes partitus; Hex: hexacorales; Hid: hidrocorales; Oct: octocorales; Are: arena; Esp: esponjas; Roc: rocas y escombro.
; Prof: profundidad; Rug: rugosidad; 1-5: número total de hoyos de entre 1 y 5 cm; 6-15: número total de hoyos de entre 6 y 15 cm; 16-30: número total de hoyos de entre 16 y 30 cm.")
Gráfica del análisis de redundancia basado en las matrices de abundancias de las especies y de variables de la complejidad arrecifal durante el ciclo anual.
C. cyanea: Chromis cyanea; G. loreto: Gramma loreto; S. taeniopterus: Scarus taeniopterus; S. partitus: Stegastes partitus; Hoyos: densidad de hoyos (hoyos/m2); Prof: profundidad; Rug: rugosidad; 1-5: número total de hoyos de entre 1 y 5 cm; 6-15: número total de hoyos de entre 6 y 15 cm; 16-30: número total de hoyos de entre 16 y 30 cm.
Las altas abundancias y densidades de reclutas registradas en los sitios con hábitat de cordillera podrían deberse a las significativamente mayores tasas de reclutamiento que presentaron C. cyanea y G. loreto en este tipo de hábitat, las cuales son especies que suelen constituir cardúmenes numerosos (DeLoach y Humann, 1999). En el caso de C. cyanea se han registrado mayores reclutamientos en zonas donde existen altas abundancias de organismos de la misma especie (Anderson et al., 2007). El asentamiento gregario es un fenómeno común en invertebrados marinos (Burke, 1986) y peces de arrecife (Hixon y Webster, 2002); entre las ventajas de esta característica se ha mencionado que los residentes coespecíficos pueden servir como una señal de hábitat disponible, y que la agregación puede reducir el riesgo de depredación (Anderson et al., 2007).
Aunque los valores de densidad y abundancia de reclutas de las especies estudiadas presentaron sus máximos en los meses de verano, ninguna de ellas mostró diferencias significativas asociadas directamente a alguno de estos meses. Otros estudios realizados en el Caribe tampoco han detectado estacionalidad significativa, sino una alta variabilidad en los picos de reclutamiento de diversas especies, los cuales suelen presentarse entre mayo y noviembre (Booth y Beretta, 1994; Tupper y Hunte, 1994; Vallès et al., 2008). La relación observada entre los meses de verano y los picos de reclutamiento podría deberse a las variaciones en la temperatura del mar, la cual, en el presente estudio, varió en un rango de 5.9°C. Sponaugle, Grorud-Colvert y Pinkard (2006)y Wilson y Meekan (2002) han registrado que la temperatura influye en el crecimiento larval, lo cual podría resultar en picos de reclutamiento.
Las regresiones multivariables también indicaron que los cambios temporales no explican de manera significativa la variación múltiple de las abundancias, con la variación espacial como covariable. Este resultado refuerza la idea de que en el Atlántico tropical occidental el reclutamiento de la mayoría de los taxones ocurre a lo largo del año (Vallès et al., 2008), por lo cual, los patrones estacionales son menos marcados que los registrados para la Gran Barrera Arrecifal (Robertson y Kaufmann, 1998). Tanto S. partitus como G. loreto tuvieron una presencia continua a lo largo del ciclo anual, con un máximo de reclutamiento en agosto.
Los sitios con hábitat de cordillera, junto con P3, tendieron a presentar los mayores valores de rugosidad, densidad y número de hoyos de 3 categorías, probablemente debido a sus comparativamente altas coberturas de hexacorales e hidrocorales. Gratwicke y Speight (2005) plantean que la rugosidad y los porcentajes de coberturas de sustratos duros están correlacionados, por lo que áreas con mayor cobertura de sustrato duro son también más complejas topográficamente. Los valores más bajos en las variables mencionadas entre los sitios con hábitat de cordillera se registraron en C4, y en P4 en el caso de los sitios con hábitat de parche, ambos ubicados en el extremo sur del arrecife.
Al igual que el presente trabajo, estudios previos han detectado que las características del hábitat béntico local tienen el potencial para influir en el asentamiento de los peces arrecifales debido a la selección del hábitat (Holbrook et al., 2000; Munday, 2002; Schmitt y Holbrook, 2000; Sponaugle et al., 2002), independientemente de los procesos a gran escala que afectan las abundancias larvales (Holbrook et al., 2000). Esta selección del hábitat puede explicarse tomando en cuenta que las larvas de los peces arrecifales, durante estadios tardíos, presentan nado activo (Fisher, 2005; Fisher, Bellwood y Job, 2000), tienen bien desarrolladas las capacidades sensoriales (Kingsford et al., 2002; Leis y McCormick, 2002) y pueden responder a un amplio intervalo de señales asociadas con el hábitat béntico, particularmente en zonas arrecifales (Leis y Carson-Ewart, 1999; Gutiérrez, 1998). No obstante, en ciertos casos las variaciones espaciales en el reclutamiento se han relacionado principalmente a una distribución estocástica del espacio disponible (Doherty, 2002; Villegas-Sánchez, Rivera-Madrid y Arias-González, 2010).
La complejidad del sustrato (número y tamaño de los hoyos, rugosidad y profundidad) suele influir de manera relevante en la abundancia de peces pequeños (Nemeth, 1998; Roberts y Ormond, 1987). De entre todas las variables de complejidad arrecifal que se analizaron en el presente estudio, solo la profundidad mostró relación con el reclutamiento de peces, lo que evidencia que dicha variable participa en la estructuración de la comunidad. La escasa relación entre el reclutamiento y las demás variables de la complejidad estudiadas podría deberse a que la selección que los peces arrecifales hacen del hábitat opera a múltiples escalas espaciales (Kingsford et al., 2002).
Las comparaciones a nivel de hábitat indicaron que C. cyanea y G. loreto prefirieron los sitios con cordilleras, y según el análisis de ordenación, estas especies mostraron una relación positiva con la cobertura de esponjas. A este respecto, numerosos estudios han registrado una fuerte fidelidad de reclutas de ciertas especies de peces arrecifales a microhábitats específicos y preferencias de hábitat (Holbrook et al., 2000; Munday, 2002; Schmitt y Holbrook, 2000; Sponaugle et al., 2002). En cuanto a las especies S. partitus y S. taeniopterus, no se detectaron preferencias claras por alguno de los hábitats muestreados.
Se observó a S. partitus y G. loreto en todos los sitios, aunque como se menciona anteriormente, G. loreto se registró con mayores abundancias en las cordilleras, a menudo debajo de repisas o justo dentro de cavidades y fisuras de corales, tal como documentan DeLoach y Humann (1999). La especie S. partitus presentó su mayor valor de abundancia promedio en el sitio más somero estudiado, con hábitat de parche, seguido por los sitios con hábitat de cordillera. En el resultado del análisis de RDA se puede observar una tendencia de esta especie a reclutarse en sitios con altos porcentajes de cobertura de rocas y escombro, así como de arena. Este resultado es compatible con los de Vallès et al. (2008), quienes observaron que S. partitus prefiere parches discretos de hábitats físicamente complejos rodeados por sustrato plano para maximizar el campo de visión (idea original de Rilov, Figueira, Lyman y Crowder, 2007), aunque tiene la capacidad de reclutarse en hábitats con muy diversas características (Danilowicz, Tolimieri y Sale, 2001; Sponaugle y Cowen, 1996), lo cual explica su presencia en todos los sitios.
Puesto que la composición del sustrato es relativamente estable en el tiempo y fácil de medir, resulta una característica del hábitat béntico conveniente para el estudio de patrones de reclutamiento en especies ícticas (Vallès et al., 2008). No obstante, la densidad y abundancia de reclutas puede variar considerablemente año tras año (Williams y Sale, 1981), y a diferentes escalas espaciales se pueden presentar diferentes patrones (Caselle y Warner, 1996; Levin, 1994; Tolimieri, 1995), por lo tanto, es necesario dar continuidad a este tipo de investigaciones para obtener descripciones consistentes de las tendencias en el reclutamiento. Los resultados del presente estudio evidencian una relación directa entre altas tasas de reclutamiento de peces y ciertas condiciones ambientales, particularmente en sitios profundos (en términos espaciales) y meses de verano (en términos temporales). Ya que en el hábitat de cordillera se registraron los mayores valores de densidad y abundancia, se propone considerar los lugares con este tipo de hábitats como sitios prioritarios para la conservación de las poblaciones de peces arrecifales como C. cyanea y G. loreto.
Agradecemos al personal de la Reserva de la Biosfera Banco Chinchorro el apoyo proporcionado para llevar a cabo los muestreos, en especial a Felipe Fonseca-Peralta, José Castro-Gutiérrez y María del Carmen García-Rivas. También agradecemos a Gaspar Quezada su colaboración en las salidas al campo y a los compañeros de laboratorio que nos apoyaron durante el trabajo de campo: Juan Carlos Fuentes, Fabián Rodríguez-Zaragoza y Gilberto Acosta-González.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.