









A survey of psocopterans in the Zapotitlán Salinas Valley, Puebla, was conducted from October 1994 to October 1995. A total of 582 specimens were collected, representing 25 species, belonging to 15 genera and 8 families (18.38% of the species recorded in the state of Puebla). Six species constitute 67% of the total of specimens collected, and 9 species represent 3.4% of the same total. Twelve species were associated to thorn scrub, 16 were found on izotal, and 6 species were found on Bursera spp. trees. It was found that both richness of species and relative abundance are related to the rainfall regime of the area (r=0.595 and r=0.739, respectively).
Se llevó a cabo un censo de psocópteros en el valle de Zapotitlán Salinas, Puebla, de octubre de 1994 a octubre de 1995. Se recolectaron 582 ejemplares, que corresponden a 25 especies, en 15 géneros y 8 familias, lo que constituye el 18.38% del total de especies registradas en el estado de Puebla. Seis especies representan el 67% del total de ejemplares recolectados y 9 el 3.4% del mismo total; 12 especies se encontraron asociadas con el matorral espinoso, 16 especies se encontraron en el izotal y 6 especies se encontraron en árboles de Bursera spp. Se encontró que tanto la riqueza como la abundancia de especies se encuentran relacionadas con el régimen de lluvias del área (r=0.595 y r=0.739, respectivamente).
In Mexico, arid and semi-arid zones occupy about 50% of the territory, from the north to the center of the country, including the coastal plains of Baja California and Sonora (Rzedowski, 1978; Briones, 1994; González-Medrano, 2004). It has been recently pointed out that the Mexican deserts concentrate a sizable proportion of biological richness, as well as a high percentage of endemisms, both of flora and fauna, which accentuates as the distribution becomes more tropical (Valiente-Banuet, 1990), offering a sharp contrast with the idea that arid zones are uninteresting, species poor areas. Among the Mexican arid and semiarid zones, the region of Tehuacán-Cuicatlán, in southern Puebla and northwestern Oaxaca, stands out with its biological richness, particularly by the endemisms of its flora (Dávila et al., 1993), its fossil deposits of the Lower Cretaceous, the archaeological remains of different pre-hispanic settlements, and by its cultural richness, merits that contributed to raise the area to the status of Biosphere Reserve (Semarnat, 1998). It is considered a megadiversity center (Dávila, 1997) and one of the most diverse arid zones of North America (Casas et al., 2001).
The valley of Zapotitlán Salinas is found within that protected natural area. It is a semi-arid zone in SE Puebla (18°20' N, 97°28' W), at the NE limit of the Valley of Tehuacán (Zavala-Hurtado, 1982). It has an area of 86.76sq. km, and constitutes a physiographic unit of the floristic province of Tehuacán-Cuicatlán (Rzedowski, 1978). The vegetation of the area is considered as a relict of the thorn scrub that once dominated the whole valley, and although it shows little perturbation, the anthropogenic pressures and the marginal character of the inhabitants of the area represent a serious threat to the stability of the ecosystem (Osorio et al., 1996). The area has archaeological vestiges of the Mixtec and Zapotec cultures, as well as geologic outcrops, fossil beds, and a variety of soil types (Byers, 1967; Valiente-Banuet et al., 2000; Arias et al., 2001). The valley of Tehuacán-Cuicatlán has been an area of interest for floristic, phytogeographic and ethnobotanical research; about 10% of the Mexican flora is found there, including 365 endemic species, and 808 species useful for humans, in an area of 10 000sq. km (Smith, 1965; Villaseñor et al., 1990; Dávila et al., 1993; Casas et al., 2001; Paredes-Flores et al., 2007). Such floristic richness is comparable to other areas of the country, with different sizes and vegetation, such as the state of Querétaro, Sierra de Manantlán and Chamela Biological Station, both in the state of Jalisco, and even Los Tuxtlas Biological Station, in Veracruz (Dávila et al., 2002). The regional fauna, however, is less well known than the vascular flora; the vertebrates have been relatively well studied: amphibians and reptiles (Canseco, 1996), birds (Arizmendi and Espinosa, 1996), and bats (Rojas-Martínez and Valiente-Banuet, 1996), as well as studies on the human settlements on the Valley (Byers, 1967).
As for the arthropods, particularly the insects, little has been done and there is little documented information (Dávila et al., 2002). Occasional observations indicate that there is a high diversity of scorpions, spiders, ants and termites, but the only groups that have been documented are Hemiptera (Brailovsky et al., 1994, 1995), Hymenoptera (Rico-Gray et al., 1998; Ríos-Casanova et al., 2004; Guzmán-Mendoza and Castaño-Meneses, 2007; Guzmán-Mendoza et al., 2010, 2012; Vásquez-Bolaños et al., 2011), and Lepidoptera, in particular with reference to the edible larvae of Paradirphia fumosa (Saturniidae) (Velázquez et al., 2006). With respect to psocopterans, a total of 125 and 139 species have been recorded in Puebla and Oaxaca respectively, with 17 and 22 endemic species respectively for each state (Mockford and García-Aldrete, 1996; García-Aldrete, pers inf.), but there has not been an attempt of systematic psocid collecting in the Tehuacán-Cuicatlán valley, so our objective was to survey the psocid fauna in Zapotitlán Salinas, to analyze its relation to the different vegetation types and to determine their phenology.
Material and methodsPsocids were collected monthly by direct sampling, in 9 localities, from October 1994 to October 1995, except for July 1995 (due to logistic problems), along the highway Tehuacán-Huajuapan de León, and on the road to the village of Santa Ana Teloxtoc, between 18°17'−18°25' N, 97°26'−97°34' W (Fig. 1), with an accumulated effort during the collecting period of 108 man/hours. At the end 12 of the 13 samples that had been planned were carried out. Altitude in the study area ranges from 1 300 to 1 700m asl. The weather is dry with summer rains (BSohw”(w)(e)g), temperature throughout the year varies between 18–22°C, the rainfall is about 400mm a year, and there is a well defined “canícula” in the middle of the rainy season (Zavala-Hurtado, 1982). The vegetation is arid tropical scrub (Leopold, 1950), which has been classified in different units according to its physonomy (Zavala-Hurtado, 1982; Osorio et al., 1996). This work follows the classification of Zavala-Hurtado (1982): 1, “thorn scrub”, constituted mostly by thorny leguminous shrubs, mixed with Agave spp., small cacti and low trees. 2, “tetechera”, defined by dominance of Neobuxbaumia columnar cacti. 3, “izotal”, with dominance of Yucca periculosa Baker and Beaucarnea gracilis Lem., and 4, “cardonal”, with dominance of the cactus Cephalocereus columna-trajani (Karw. Ex Pfeiff.) P. V. Heath. Besides, we established a unit called “Bursera spp.”, to include Bursera spp. trees, not considered in the classification, but sampled for psocids.

In each locality, the different vegetation units were sampled systematically, taking care that the same group of plants were not sampled in a period of less than 3 months. Psocids were collected by directly examining tree trunks and columnar cacti, and beating branches and trunks over a collecting cloth, from where they were taken with a mouth aspirator to be preserved in 80% ethyl alcohol. The identification was made by examination of head, wings, legs and genitalia mounted on slides in Canada Balsam. For the mounting technique see García-Aldrete (1990). The specimens collected are deposited in the National Insect Collection (CNIN), Departamento de Zoología, Instituto de Biología, UNAM.
The richness and relative abundance values correspond to number of species and specimens collected. Species diversity of the area was estimated with the Simpson index (1 - Σpi2, cf. Moreno [2001]) using Microsoft Excel 2007. To establish the efficiency of the collecting, the species accumulation curve was obtained using PRIMER v6 (Clarke and Gorley, 2006) with Jacknife 1 estimator, which is based on the number of species seen only in 1 sample (Moreno, 2001). The slope of the curve was calculated from Clench's equation (Soberón and Llorente, 1993). The relation between specific richness with rainfall, as well as relative abundance of species with rainfall, was determined by the Pearson correlation coefficient (Zar, 2010). Both calculations were performed with STATISTICA 7.0 (StatSoft, 2004).
ResultsComposition of the fauna. During the period, 582 specimens were collected, representing 25 species in 15 genera and 8 families (Table 1). A total of 13 species (52%) have already been described, and 12 (48%) are new to science. The suborder Trogiomorpha is the less represented, with only 1 species each in the genera Rhyopsocus and Psyllipsocus. The suborder Troctomorpha is represented by 4 species, all in the genus Liposcelis. The suborder Psocomorpha is the most diverse, with 19 species in 12 genera. The family Psocidae is the largest and more diverse, with 8 genera and 10 species, and the most diverse genera are Liposcelis and Lachesilla, with 4 species each. None of the 25 species collected had been recorded before in Zapotitlán Salinas, and 11 of them are new state records: the 4 species of Liposcelis, Asiopsocus tehuacanus García-Aldrete and Casasola, Valenzuela sp. 2, Blaste sp. 2, Hyalopsocus sp., Indiopsocus bisignatus Banks, Steleops monticola García-Aldrete and Trichadenotecnum desolatum (Chapman), with which the psocid fauna of Puebla is raised to 136 species. A. tehuacanus García-Aldrete and Casasola is endemic to the area, and Lachesilla tehuautlensis García-Aldrete only occurs in the Tehuacán-Cuicatlán Valley and in Huautla, Morelos, some 130km E of the type locality, ca. Zapotitlán Salinas (García-Aldrete and Casasola, 1995; García-Aldrete, 2000).
Species of Psocoptera from Zapotitlán Salinas Valley, Puebla
| Number of specimens | Thorn Scrub | Izotal | Bursera spp. | |
|---|---|---|---|---|
| TROGIOMORPHA | ||||
| Psoquillidae | ||||
| 1 Rhyopsocus concavus García-Aldrete | 4 | * | ||
| Psyllipsocidae | ||||
| 2 Psyllipsocus oculatus Gurney | 16 | * | ||
| TROCTOMORPHA. | ||||
| Liposcelididae | ||||
| 3 Liposcelis bostrychopila Badonnel | 36 | * | * | |
| 4 Liposcelis sp. 1 | 1 | * | ||
| 5 Liposcelis sp. 2 | 94 | * | * | * |
| 6 Liposcelis sp. 3 | 4 | * | * | |
| PSOCOMORPHA | ||||
| Asiopsocidae | ||||
| 7 Asiopsocus tehuacanus García-Aldrete and Casasola González | 11 | * | ||
| Caeciliusidae | ||||
| 8 Valenzuela totonacus (Mockford) | 27 | * | ||
| 9 Valenzuela sp. 1 | 1 | * | ||
| 10 Valenzuela sp. 2 | 2 | * | ||
| Lachesillidae | ||||
| 11 Lachesilla fuscipalpis Badonnel | 13 | * | ||
| 12 L. palmicola García-Aldrete | 52 | * | ||
| 13 L. tectorum Badonnel | 1 | * | ||
| 14 L. tehuautlensis García-Aldrete | 56 | * | * | |
| Peripsocidae | ||||
| 15 Peripsocus sp. | 13 | * | ||
| Psocidae | ||||
| 16 Blaste posticata Banks | 17 | * | * | |
| 17 Blaste sp. 1 | 2 | * | * | |
| 18 Blaste sp. 2 | 32 | * | ||
| 19 Blastopsocus sp. | 26 | * | * | |
| 20 Hyalopsocus sp. | 4 | * | * | |
| 21 Indiopsocus bisignatus Banks | 11 | * | ||
| 22 Metylophorus sp. | 1 | * | ||
| 23 Ptycta sp. | 27 | * | ||
| 24 Steleops monticola García-Aldrete | 124 | * | ||
| 25 Trichadenotecnum desolatum (Chapman) | 7 | * | ||
| TOTAL | 582 | 13 | 16 | 5 |
Distribution of the abundance. Figure 2 shows the species accumulation curve for the collecting period. The estimated value of species richness, based on the nonparametric estimator Jacknife 1 (31), was higher than the observed value (25). This means that 80% of the local real richness was collected; however, the slope of the line (0.75) indicates that we did not reach the asymptotic phase of the curve (Jiménez-Valverde and Hortal, 2003), which means that richness is probably larger than what is presented here. The most abundant species were Steleops monticola, Liposcelis sp. 2, Lachesilla tehuautlensis and L. palmicola (Fig. 3). These 4 species constitute 56% of the total collected, and on the other hand, 10 species constitute less than 2% of the total collected. Four taxa are rare, as they were represented by a single individual during the collecting period, 2 in the thorn scrub (Liposcelis sp. 1 and Metylophorus sp.), and 2 in izotal (Valenzuela sp. 1 and Lachesilla tectorum).


With respect to the relation of the psocid fauna with the vegetation unit, the thorn scrub and the izotal presented the highest diversity and species abundance (Fig. 3). In the thorn scrub, 13 species were found associated with this, and S. monticola, L. tehuautlensis, Blaste sp. 2, L. bostrychopila and Ptycta sp., were the most abundant. Also, Rhyopsocus concavus, Liposcelis sp. 1, Peripsocus sp., Blaste sp. 2, Metylophorus sp., Ptycta sp. and S. monticola were exclusive to this vegetation type; most of the individuals were found on branches and foliage; S. monticola was found mostly on the surface of trunks and branches. 16 species were found associated to the izotal, living mostly on dead, hanging leaves of Yucca. The most abundant species were Liposcelis sp. 2, Lachesillapalmicola and Valenzuela totonacus, and P. oculatus, V. totonacus, Valenzuela sp. 1, Valenzuela sp. 2, L. fuscipalpis, L. palmicola, L. tectorum, Hyalopsocus sp. and T. desolatum were exclusive to this biotope. Five species were associated to Bursera trees, both on foliage and trunks; A. tehuacanus was the most abundant and was found exclusively here. Blastopsocus sp. and Liposcelis sp. 2 also recorded high abundances; 5 species were shared between thorn scrub and izotal, 2 species were shared between the thorn scrub and Bursera trees, and 3 species were shared between izotal and Bursera. No species were found on Tetechera and Cardonal.
Using the Simpson index, species richness for species in this locality was 0.89. The psocid fauna of Zapotitlán Salinas represents 18.38% of the psocid fauna recorded in Puebla, which makes it an area of relative high diversity for this group of insects.
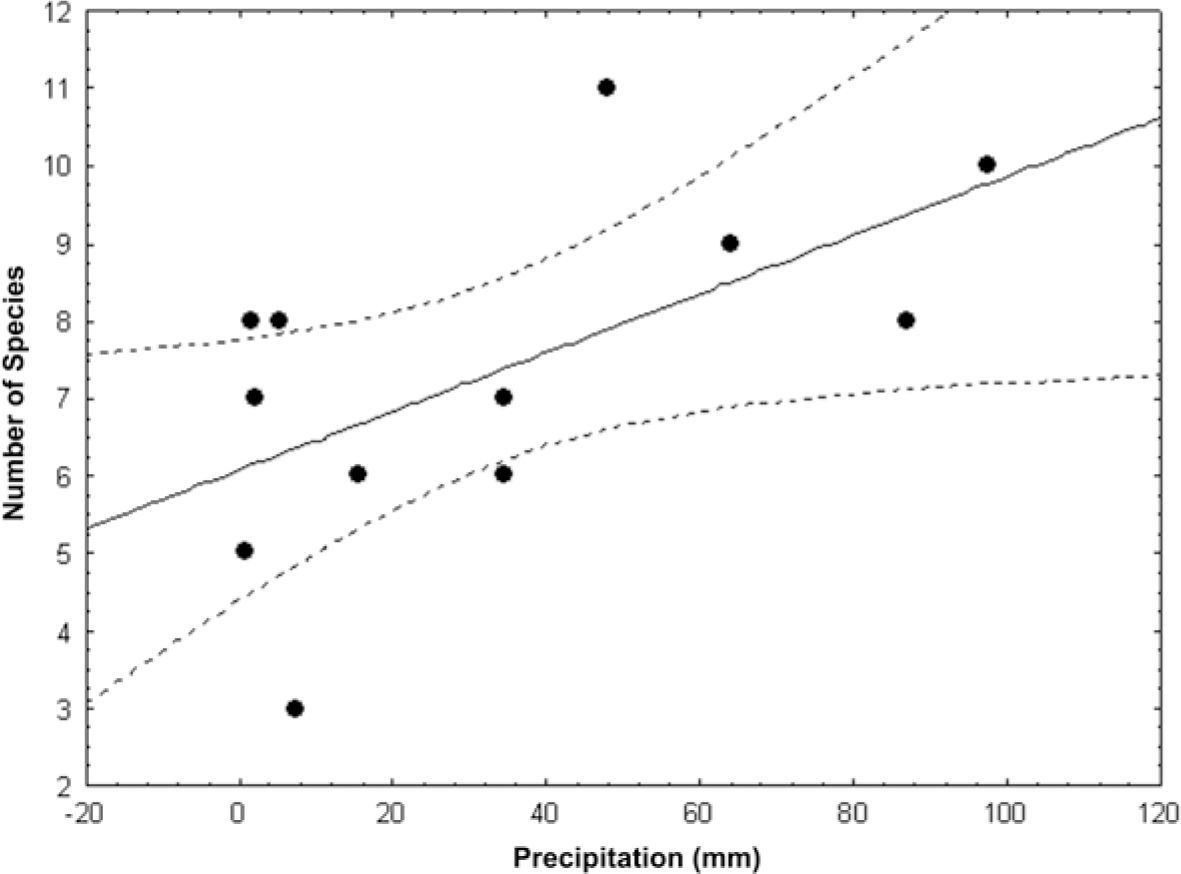
Phenology. Besides the considerable number of species found in thorn scrub, izotal and Bursera spp. trees, changes in presence and number of individuals per species were observed throughout the study period (Tables 2–4). These changes seem to be related to the rainfall regime of the area (Figs. 4, 5). The coefficient r for number of species and pluvial precipitation was 0.595, while for relative abundance the same parameter was r=0.739. In both cases, a positive correlation between the variables considered is observed, meaning that species richness and abundance increase with precipitation. Considering only the most abundant species, their relation to precipitation was (coefficient r in parentheses): L. bostrychopila (0.481), L. tehuautlensis (0.468), Liposcelis sp. 2 (0.379), S. monticola (0.064), and L. palmicola (−0.049). In the first 4 cases, the correlation is positive also, i.e., the abundance of these increase in the rainy season. In contrast, for L. palmicola the correlation is negative, which means that their abundance decrease during the rain season.
Species and number of individuals per species in thorn scrub, during the collecting period
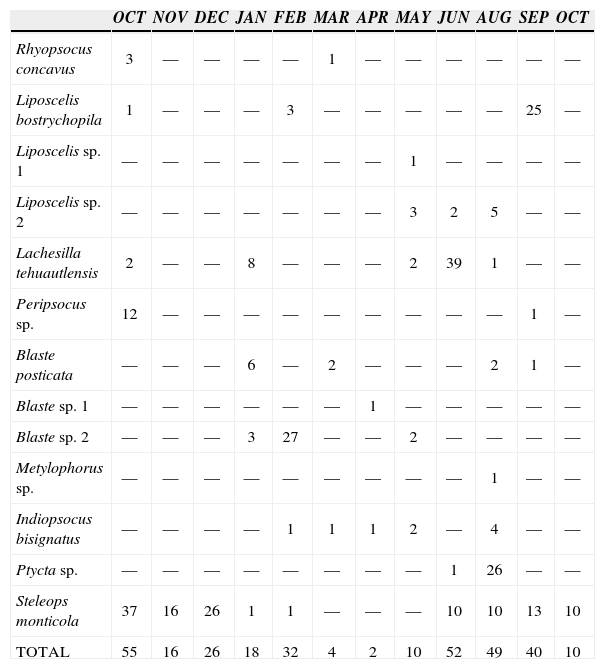
| OCT | NOV | DEC | JAN | FEB | MAR | APR | MAY | JUN | AUG | SEP | OCT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhyopsocus concavus | 3 | — | — | — | — | 1 | — | — | — | — | — | — |
| Liposcelis bostrychopila | 1 | — | — | — | 3 | — | — | — | — | — | 25 | — |
| Liposcelis sp. 1 | — | — | — | — | — | — | — | 1 | — | — | — | — |
| Liposcelis sp. 2 | — | — | — | — | — | — | — | 3 | 2 | 5 | — | — |
| Lachesilla tehuautlensis | 2 | — | — | 8 | — | — | — | 2 | 39 | 1 | — | — |
| Peripsocus sp. | 12 | — | — | — | — | — | — | — | — | — | 1 | — |
| Blaste posticata | — | — | — | 6 | — | 2 | — | — | — | 2 | 1 | — |
| Blaste sp. 1 | — | — | — | — | — | — | 1 | — | — | — | — | — |
| Blaste sp. 2 | — | — | — | 3 | 27 | — | — | 2 | — | — | — | — |
| Metylophorus sp. | — | — | — | — | — | — | — | — | — | 1 | — | — |
| Indiopsocus bisignatus | — | — | — | — | 1 | 1 | 1 | 2 | — | 4 | — | — |
| Ptycta sp. | — | — | — | — | — | — | — | — | 1 | 26 | — | — |
| Steleops monticola | 37 | 16 | 26 | 1 | 1 | — | — | — | 10 | 10 | 13 | 10 |
| TOTAL | 55 | 16 | 26 | 18 | 32 | 4 | 2 | 10 | 52 | 49 | 40 | 10 |
Species and number of individuals per species in izotal, during the collecting period
| OCT | NOV | DEC | JAN | FEB | MAR | APR | MAY | JUN | AUG | SEP | OCT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Psyllipsocus oculatus | — | — | 4 | 3 | 5 | — | — | — | — | 2 | 2 | — |
| Liposcelis bostrychopila | — | — | 2 | — | — | 5 | — | — | — | — | — | — |
| Liposcelis sp. 2 | — | 4 | 2 | — | 2 | 18 | 6 | 19 | 4 | 6 | 10 | 10 |
| Liposcelis sp. 3 | — | — | — | — | — | — | — | 1 | — | — | 1 | — |
| Valenzuela totonacus | — | — | 1 | 10 | — | — | — | — | — | — | 12 | 4 |
| Valenzuela sp. 1 | — | — | — | 1 | — | — | — | — | — | — | — | — |
| Valenzuela sp. 2 | — | — | 2 | — | — | — | — | — | — | — | — | — |
| Lachesilla fuscipalpis | 8 | — | — | — | — | 1 | 2 | — | — | — | — | 2 |
| Lachesilla palmicola | — | 7 | 4 | 15 | — | — | — | 4 | — | 8 | 6 | 8 |
| Lachesilla tectorum | — | — | — | — | — | — | — | — | — | 1 | — | — |
| Lachesilla tehuautlensis | — | — | — | — | — | — | — | — | — | — | — | 4 |
| Blaste posticata | — | — | — | 2 | — | — | — | — | — | 4 | — | — |
| Blaste sp. 1 | — | — | — | — | — | — | — | — | — | 1 | — | — |
| Blastopsocus sp. | — | — | — | — | — | — | — | — | — | — | 17 | — |
| Hyalopsocus sp. | — | — | 4 | — | — | — | — | — | — | — | — | — |
| Trichadenotecnum desolatum | — | — | — | — | — | — | — | — | — | — | — | 7 |
| TOTAL | 8 | 11 | 19 | 31 | 7 | 24 | 8 | 24 | 4 | 22 | 48 | 35 |
Species and number of individuals per species in Bursera spp. trees during the collecting period
| OCT | NOV | DEC | JAN | FEB | MAR | APR | MAY | JUN | AUG | SEP | OCT | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Liposcelis sp. 2 | — | — | — | — | — | — | 1 | — | 2 | — | — | — |
| Liposcelis sp. 3 | — | — | — | — | — | — | 1 | — | 1 | — | — | — |
| Asiopsocus tehuacanus | — | — | — | — | 1 | — | 3 | — | 7 | — | — | — |
| Blastopsocus sp. | — | — | — | — | — | — | — | — | 9 | — | — | — |
| Indiopsocus bisignatus | — | — | — | — | — | — | 1 | — | 1 | — | — | — |
| TOTAL | 0 | 0 | 0 | 0 | 1 | 0 | 6 | 0 | 20 | 0 | 0 | 0 |
 Samples. (—) Trend line.")
 Samples. (—) Trend line.")
Analyzing each vegetation unit, we found that, in the thorn scrub (Table 2), 55 specimens were collected in October 1994, 32 specimens were taken in February 1995 and 52 specimens were collected in June 1995; June and September correspond to the rain season, and October corresponds to the end of it, but still with remnant moisture. The most frequent species were B. posticata, Blaste sp. 2, I. bisignatus, L. tehuautlensis, Liposcelis sp. 2 and S. monticola, this present almost throughout the year. There are 2 periods of increase in number of individuals during the year: between December and March, in the dry season, and from May to October, in the rainy season. B. posticata and Blaste sp. 2 showed high abundance values in the dry season, while Liposcelis sp. 2, L. tehuautlensis, I. bisignatus and S. monticola were more abundant in the rain season (Table 2). The alternate succession of these species could represent a strategy to avoid competition. In izotal (Table 3), the highest number of specimens were collected in January and September (31 and 48 respectively). Liposcelis sp. 2, L. fuscipalpis, L. palmicola,P. oculatus and V. totonacus were the most frequent species and 8 species were collected only once in the year (Valenzuela spp. 1 and 2, L. tehuautlensis and L. tectorum, and Blaste sp. 1, Blastopsocus sp., Hyalopsocus sp., and T. desolatum). The same was found in the thorn scrub, there are 2 periods of high abundance in the year, during the dry and rain seasons. P.oculatus and L. palmicola are most abundant towards the end of the rain season and beginning of the dry season, and Liposcelis sp. 2 is most abundant at the end of the dry season and beginning of the rain season.V. totonacus and L. fuscipalpis are most abundant during the rainy season. Finally, in Bursera trees (Table 4) 6 specimens were collected in April and 20 in June. In this case, only A. tehuacanus was the most frequent species; however, all species were found only towards the end of the dry season and at the beginning of the rainy season.
DiscussionThe results constitute the first account of systematic psocid collecting in an arid zone in Mexico. They are a little surprising as, given the nature of the area, we expected a lower species richness, which, as it is, and even if the species accumulation curve did not reach the asymptotic phase, represents close to 20% the species richness of the state, which for the small area sampled, it is considerable. It is of interest that 11 of the 25 species collected represent new records for the state of Puebla, and that 9 species were new to science, a fact that points out that the psocid census in Mexico is far from being complete; after almost 40 years of collecting throughout the country, it is most probable that the present figure of 746 species for the country could be grossly underestimated.
The principal habitats of psocids are: 1, living foliage; 2, dead foliage; 3, ground litter; 4, bark of trees; 5, rock surface, and 6, human habitation (García-Aldrete, 1990). The psocids collected in Zapotitlán Salinas fall in categories 1, 2 and 4; the species collected in izotal correspond to dead foliage, those in thorn scrub correspond to living foliage and bark of trees, and those in Bursera correspond to bark of trees; it is of interest to point out that most of the species in these habitats were exclusive to them (cf. Table 1).
The most important species, in number of individuals were, in thorn scrub, the corticicolous S. monticola and the folicolous L. tehuautlensis, both least abundant in the dry season, and in izotal, the most important species were Liposcelis sp. 2 and L. palmicola, both folicolous, with the former being most abundant in the dry season and the latter being most abundant in the rain season; the above probably indicates that these species avoid competition. An identical situation was found by Herrera-Fuentes (1986), who studied the population fluctuation of Lachesilla alpha García-Aldrete and Liposcelis sp., on dead hanging leaves of Furcraea parmentieri at Cerro del Ajusco, Mexico D. F. Her study reported that both species alternate abundance, in relation to the rainfall regime of the area, the peaks of abundance of Liposcelis sp., occur during the dry season, while the peak of abundance of L. alpha occur during the rain season.
JACG thanks the Biology Department (UAM-Iztapalapa), José Alejandro Zavala Hurtado, Pedro Luis Valverde, Fernando Vite, and Héctor Moya, for valuable support during the period of field work. We also thank Harry Brailovsky (Instituto de Biología, UNAM) for critical comments on a previous draft of this work and two anonymous reviewers for their valuable comments and suggestions to improve the manuscript. ANGA thanks Instituto de Biología, UNAM, for supporting his research over the years.