



Las pteridofitas han sido escasamente estudiadas en el norte de México, ya que existen sierras donde se desconocen las especies que habitan en ellas. En el presente estudio se determinaron las especies de pteridofitas de 2 sierras del estado de Durango, México (El Sarnoso y Mapimí), se calculó el índice de diversidad beta para comparar la diversidad de estas plantas en ambas áreas y se representó la ubicación de cada especie en mapas de distribución geográfica. Se registraron 28 especies distribuidas en 4 familias y 9 géneros. El coeficiente de similitud fue de 0.89, con 25 especies en la sierra El Sarnoso y 28 en la de Mapimí. Se destacó la presencia de la especie indicadora de impacto ambiental antropogénico Pellaea ternifolia var. ternifolia en áreas de extracción de material rocoso. Otras especies registradas fueron Pleopeltis polypodioides var. michauxianum (=Polypodium polypodioides var. michauxianum), Adiantum capillus-veneris, Cheilanthes notholaenoides y Selaginella lepidophylla. El conocimiento de estas especies puede marcar la pauta para comenzar a elaborar programas de conservación en ambas sierras, ya que si no se controla la extracción de materiales rocosos, el hábitat se irá degradando progresivamente, afectando la supervivencia de las especies de flora y fauna que habitan en ellas.
Pteridophytes have been inadequately studied in northern México, resulting in large, remote areas where the species of pteridophytes that inhabit them are unknown. We surveyed pteridophytes in 2 mountain areas in the state of Durango, México (El Sarnoso and Mapimí). We plotted location of each species or subspecies and then calculated beta diversity to compare between-area diversity for this group of plants. A total of 28 species grouped in 4 families and 9 genera were recorded. The coefficient of similarity was 0.89, with 25 species in El Sarnoso and 28 in Mapimí. An important finding was the presence of an indicator species of anthropogenic environmental impact, Pellaea ternifolia var. ternifolia in areas of rock extraction. Other recorded species were Pleopeltis polypodioides var. michauxianum (=Polypodium polypodioides var. michauxianum), Adiantum capillus-veneris, Cheilanthes notholaenoides, and Selaginella lepidophylla. Knowledge of regional biodiversity, including pteridophytes, can support the development of conservation programs in these mountains. Without effective conservation programs in place, and if the removal of rock materials is not controlled, the habitat of these mountains will decay progressively, affecting the survival of flora and fauna species that inhabit these areas.
En la actualidad el alto grado de impacto antropogénico sobre la biodiversidad está desencadenando altas tasas de extinción de especies en lo que se considera como el sexto evento de extinciones masivas en la historia de la vida en el planeta (Dirzo y Raven, 2003). Esta crisis de la biodiversidad, que se manifiesta en una creciente degradación y agotamiento de los ecosistemas, ha promovido en las últimas décadas un creciente interés científico, social, económico y ético por encontrar estrategias y criterios científicos sobre los cuales basar la conservación de la biodiversidad (Halffter, 2000; Morrone, 2000; Novacek, 2008).
Las licofitas y los helechos son plantas vasculares que tradicionalmente se conocen como «pteridofitas», debido a que ambos grupos presentan reproducción por esporas (Schuettpelz y Pryer, 2008). Son plantas bioindicadoras que poseen características morfológicas, ciclos de vida y requerimientos de microhábitats que las predisponen a ser vulnerables a las alteraciones ambientales (Arcand y Ranker, 2008; De la Sota, 1973; Lindenmayer, Margules y Botkin, 2000; Moran, 1995; Page, 1985). Comparándolas con las angiospermas, este grupo de plantas posee patrones de distribución que están relacionados con factores abióticos, por ejemplo, dispersión anemocórica, y presentan relaciones coevolutivas a través de vectores bióticos (Barrington, 1993; Kessler, 2010; Moran, 2008). Además, su antigüedad, monofilia y estabilidad morfológica son indicadores para establecer patrones biogeográficos (Arana, Morrone, Ponce y Oggero, 2011). Por otra parte, estas plantas se consideran extremadamente vulnerables a la extinción; sin embargo, la fragmentación del hábitat y la deforestación favorecen su diversidad, debido a que se establecen los nichos ecológicos que estas plantas requieren (Arcand y Ranker, 2008).
Las pteridofitas son plantas diversas, ya que se estima la existencia de 15,000 especies, de las cuales ya han sido descritas más de 12,000 (Moran, 2008). En México se han documentado 1,014 especies (10% de la diversidad mundial), de las cuales 188 (18%) son endémicas (Martínez-Salas y Ramos, 2014). En general, la alta diversidad y abundancia de las pteridofitas en nuestro país se debe a su ubicación geográfica, así como a su geología, relieve, tipos de suelo y sus múltiples tipos de vegetación (Riba, Pérez-García y Orozco-Segovia, 1993). En particular, las zonas áridas del norte de México se consideran especialmente importantes en plantas vasculares no solo por su diversidad, sino por el alto grado de endemismos (González-Medrano y Chiang-Cabrera, 1988; Mickel y Smith, 2004; Rzedowski, 1978). Sin embargo, a la fecha solo se han realizado inventarios de pteridofitas en algunas regiones de los estados del norte como Chihuahua (Estrada-Castrillón y Villarreal-Quintanilla, 2010; Knobloch y Correll, 1962), Sonora (Shreve y Wiggins, 1964), Nuevo León (Aguirre-Claverán, 1983), Coahuila (Alba-Ávila, 2011) y Durango en la Reserva de la Biosfera La Michilía (González-Elizondo, González-Elizondo y Cortés-Ortiz, 1993). En el estado de Durango existen sierras y cadenas montañosas donde se desconocen las especies de pteridofitas que habitan en ellas. Un ejemplo son las sierras El Sarnoso y Mapimí y sus alrededores, ya que debido a su importancia ecológica existen planes a medio plazo para proponer estas sierras como un área natural protegida estatal (Orona-Espino y Estrada-Rodríguez, 2012). En el presente estudio se determinaron las especies de este grupo de plantas, se calculó el índice de diversidad beta para comparar la diversidad entre ambas sierras y se representó la ubicación de cada especie o subespecie utilizando sistemas de distribución geográfica.
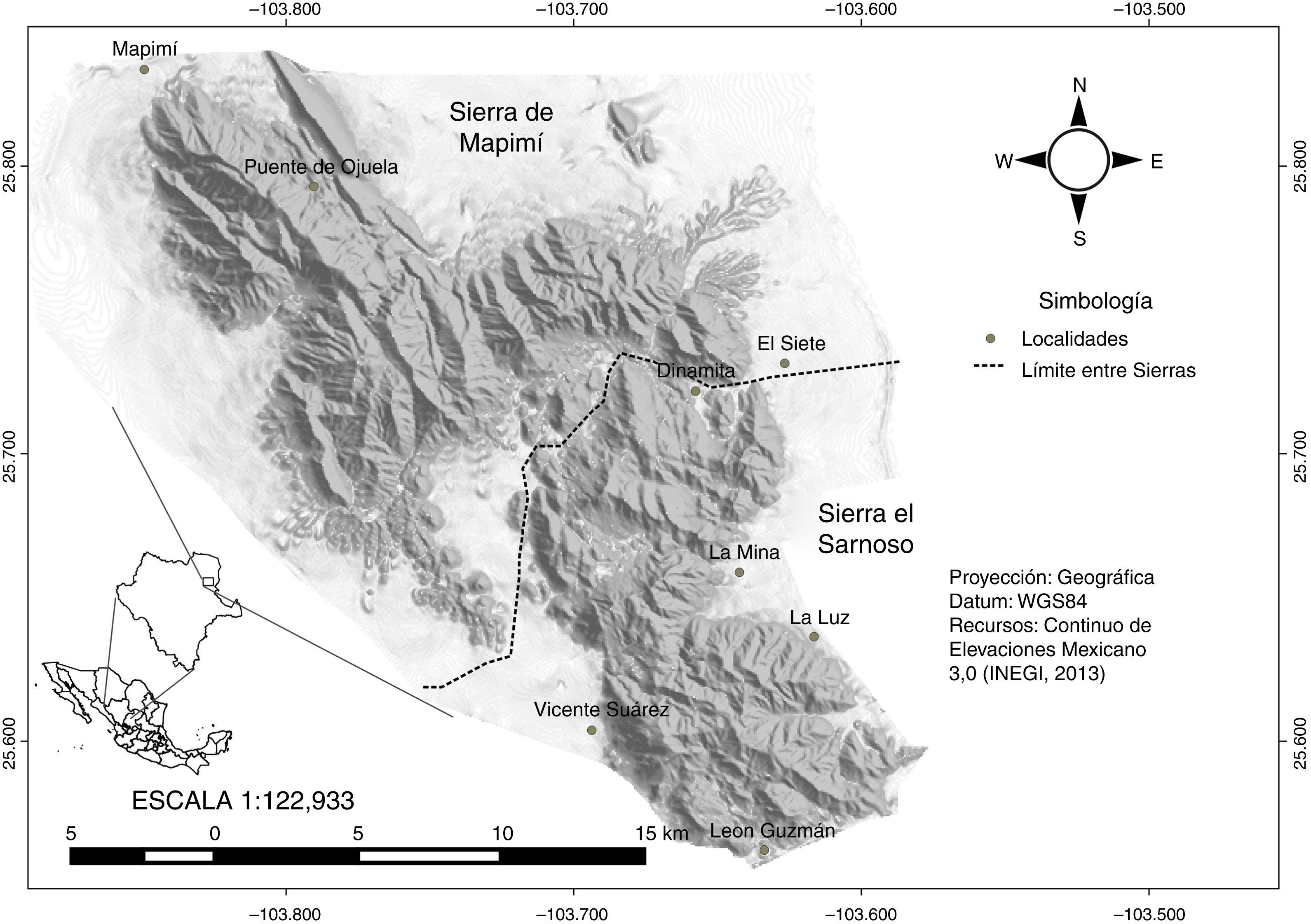
Materiales y métodosTrabajo de campoLas principales localidades en las que se llevó a cabo el muestreo de pteridofitas en la sierra de El Sarnoso fueron Dinamita, La Luz, León Guzmán y Mina; en la sierra de Mapimí se visitó Mapimí, Vicente Suárez, Puente de Ojuela y El Siete. Todas estas localidades fueron consideradas para la realización del estudio técnico justificativo de ambas sierras (Orona-Espino y Estrada-Rodríguez, 2012; fig. 1). Para cada localidad se realizaron 4 visitas en verano-otoño de 2012 y primavera-invierno de 2013. Se recorrieron 50 transectos de longitud y dirección variable, aproximadamente 3±1km, en donde se realizó una búsqueda exhaustiva de pteridofitas. Para cada individuo de pteridofita observado se registró su ubicación geográfica mediante GPS (Garmin 12XL) y se tomó fotografía. Se colectó un total de 84 individuos, los cuales se colocaron en una prensa botánica y se preservaron de acuerdo con la metodología propuesta por Arreguín-Sánchez en Lot y Chiang (1986).
Trabajo de laboratorio.")
La determinación taxonómica se basó en las claves de Mickel y Smith (2004). El cotejo de cada individuo se realizó con base en colecciones botánicas formales en los siguientes herbarios: Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional (ENCB); Departamento de Botánica, Herbario Universidad Autónoma Agraria Antonio Narro (UAAAN) y Herbario de la Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León (CFNL). Como acciones de intercambio institucional, se depositaron individuos de las especies recolectadas en los 3 herbarios antes mencionados. El arreglo taxonómico de las pteridofitas se basó en el Catálogo de Autoridades Taxonómicas de Helechos y Plantas Afines (pteridofitas) de México (Conabio, 2008). Finalmente, los ejemplares fueron montados en láminas de papel bristol color blanco de 28x40cm, se etiquetaron y guardaron en pliegos de papel manila color ámbar y fueron depositados en los herbarios ENCB, UAAAN y CFNL.
Análisis de datosPara estimar la diversidad beta, riqueza específica compartida, de las pteridofitas de la sierra El Sarnoso y Mapimí, se utilizó el coeficiente de similitud de Jaccard debido a su confiabilidad en el análisis de datos de presencia/ausencia (Sánchez y López, 1988). Esta información también se representó mediante un dendrograma utilizando un análisis de conglomerados jerárquicos para agrupar las localidades según su similitud en cuanto a especies de pteridofitas. Finalmente, para realizar los mapas de distribución de las pteridofitas se empleó el programa ARCGIS 9.3.1. Se utilizó el polígono «El Sarnoso» (Romero-Méndez y Martínez-Ríos, 2009) para delimitar el área de mapeo. Se utilizaron puntos geográficos colectados mediante GPS de cada organismo y se importaron en formato de tablas al programa transformándolos en formato «shapefile», a los cuales se les aplicaron geoprocedimientos como el «Definition Query» para cada una de las especies. Para la elaboración de los mapas se utilizaron imágenes de satélite de World Imagery como fondo y se exportaron a imagen jpg.
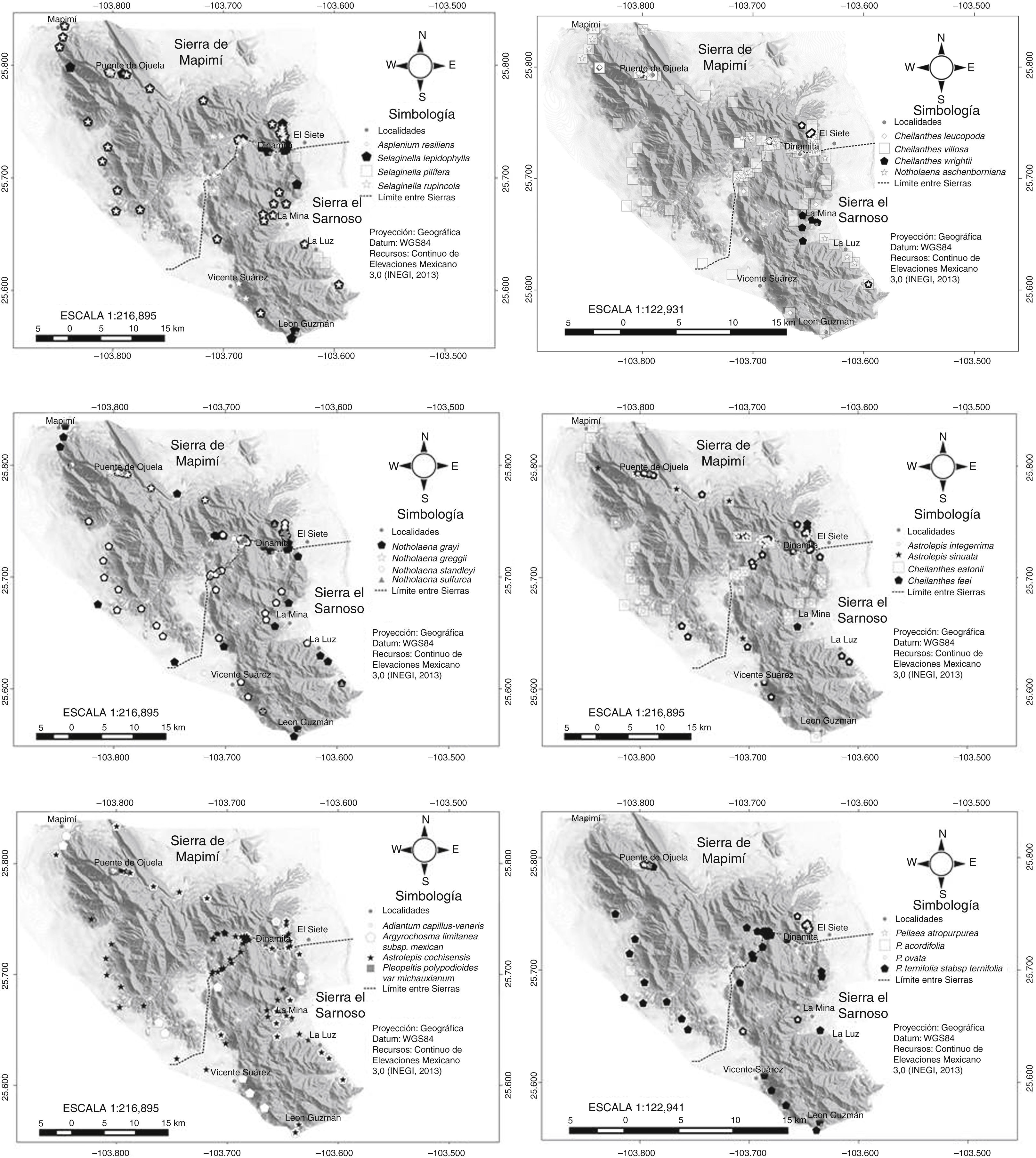
ResultadosSe observó que la flora pteridológica de las sierras El Sarnoso y Mapimí está conformada por 4 familias, 9 géneros, 28 especies y 3 variedades (tabla 1). La familia con mayor número de especies fue Pteridaceae (22), seguida de Selaginellaceae (3 especies) y, por último, Polypodiaceae y Aspleniaceae. Los géneros que presentaron un mayor número de especies fueron Cheilanthes, Pellaea y Notholaena. En la figura 2 se ubican geográficamente las diferentes especies de pteridofitas encontradas en ambas sierras, y en la figura 3 se muestran imágenes de estas plantas.
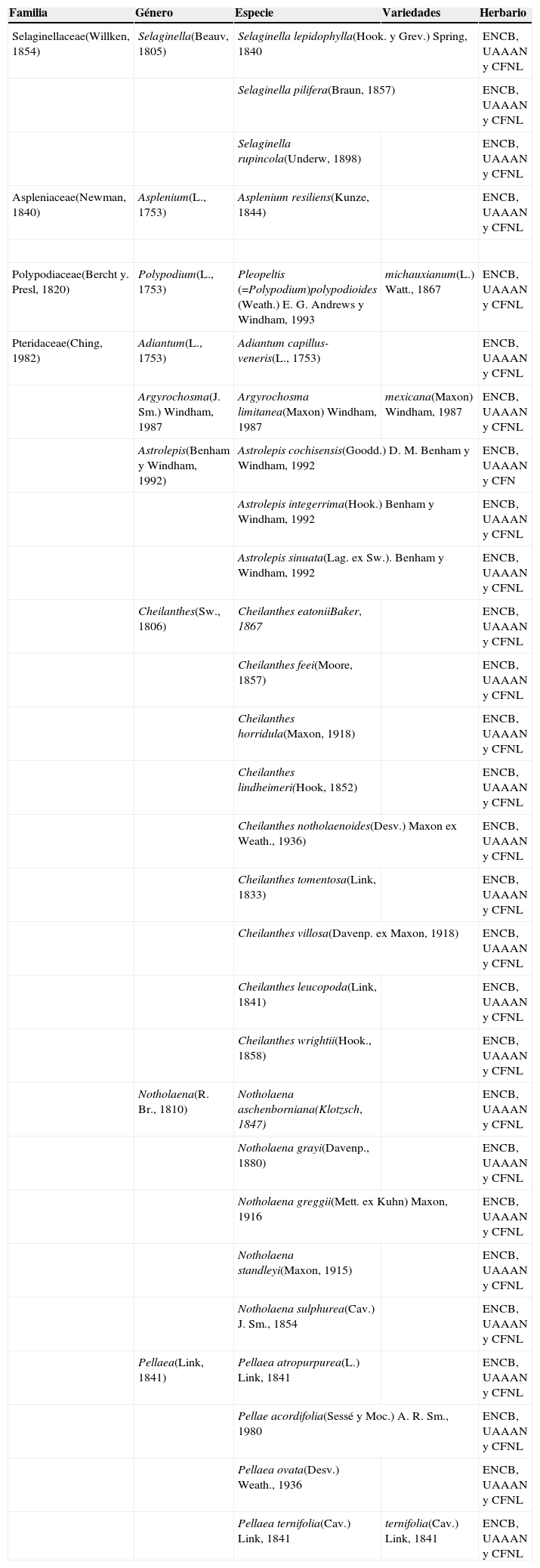
Listado de las pteridofitas de las sierras El Sarnoso y Mapimí (clasificación según Conabio, 2008) y herbarios de referencia donde fueron cotejadas y depositadas.
| Familia | Género | Especie | Variedades | Herbario |
|---|---|---|---|---|
| Selaginellaceae(Willken, 1854) | Selaginella(Beauv, 1805) | Selaginella lepidophylla(Hook. y Grev.) Spring, 1840 | ENCB, UAAAN y CFNL | |
| Selaginella pilifera(Braun, 1857) | ENCB, UAAAN y CFNL | |||
| Selaginella rupincola(Underw, 1898) | ENCB, UAAAN y CFNL | |||
| Aspleniaceae(Newman, 1840) | Asplenium(L., 1753) | Asplenium resiliens(Kunze, 1844) | ENCB, UAAAN y CFNL | |
| Polypodiaceae(Bercht y. Presl, 1820) | Polypodium(L., 1753) | Pleopeltis (=Polypodium)polypodioides (Weath.) E. G. Andrews y Windham, 1993 | michauxianum(L.) Watt., 1867 | ENCB, UAAAN y CFNL |
| Pteridaceae(Ching, 1982) | Adiantum(L., 1753) | Adiantum capillus-veneris(L., 1753) | ENCB, UAAAN y CFNL | |
| Argyrochosma(J. Sm.) Windham, 1987 | Argyrochosma limitanea(Maxon) Windham, 1987 | mexicana(Maxon) Windham, 1987 | ENCB, UAAAN y CFNL | |
| Astrolepis(Benham y Windham, 1992) | Astrolepis cochisensis(Goodd.) D. M. Benham y Windham, 1992 | ENCB, UAAAN y CFN | ||
| Astrolepis integerrima(Hook.) Benham y Windham, 1992 | ENCB, UAAAN y CFNL | |||
| Astrolepis sinuata(Lag. ex Sw.). Benham y Windham, 1992 | ENCB, UAAAN y CFNL | |||
| Cheilanthes(Sw., 1806) | Cheilanthes eatoniiBaker, 1867 | ENCB, UAAAN y CFNL | ||
| Cheilanthes feei(Moore, 1857) | ENCB, UAAAN y CFNL | |||
| Cheilanthes horridula(Maxon, 1918) | ENCB, UAAAN y CFNL | |||
| Cheilanthes lindheimeri(Hook, 1852) | ENCB, UAAAN y CFNL | |||
| Cheilanthes notholaenoides(Desv.) Maxon ex Weath., 1936) | ENCB, UAAAN y CFNL | |||
| Cheilanthes tomentosa(Link, 1833) | ENCB, UAAAN y CFNL | |||
| Cheilanthes villosa(Davenp. ex Maxon, 1918) | ENCB, UAAAN y CFNL | |||
| Cheilanthes leucopoda(Link, 1841) | ENCB, UAAAN y CFNL | |||
| Cheilanthes wrightii(Hook., 1858) | ENCB, UAAAN y CFNL | |||
| Notholaena(R. Br., 1810) | Notholaena aschenborniana(Klotzsch, 1847) | ENCB, UAAAN y CFNL | ||
| Notholaena grayi(Davenp., 1880) | ENCB, UAAAN y CFNL | |||
| Notholaena greggii(Mett. ex Kuhn) Maxon, 1916 | ENCB, UAAAN y CFNL | |||
| Notholaena standleyi(Maxon, 1915) | ENCB, UAAAN y CFNL | |||
| Notholaena sulphurea(Cav.) J. Sm., 1854 | ENCB, UAAAN y CFNL | |||
| Pellaea(Link, 1841) | Pellaea atropurpurea(L.) Link, 1841 | ENCB, UAAAN y CFNL | ||
| Pellae acordifolia(Sessé y Moc.) A. R. Sm., 1980 | ENCB, UAAAN y CFNL | |||
| Pellaea ovata(Desv.) Weath., 1936 | ENCB, UAAAN y CFNL | |||
| Pellaea ternifolia(Cav.) Link, 1841 | ternifolia(Cav.) Link, 1841 | ENCB, UAAAN y CFNL | ||


 Selaginella lepidophylla; 2) S. pilifera; 3) S. rupincola; 4) Asplenium resiliens; 5) Pleopeltis polypodioides var. michauxianum; 6) Adiantum capillus-veneris; 7) Argyrochosma limitanea var. mexicana; 8) Astrolepis cochisensis; 9) A. integerrima; 10) A. sinuata; 11) Cheilanthes eatonii; 12) C. feei; 13) C. horridula; 14) C. lindheimeri; 15) C. notholaenoides; 16) C. tomentosa; 17) C. villosa; 18) C. leucopoda; 19) C. wrightii; 20) Notholaena aschenborniana; 21) N. grayi; 22) N. greggii; 23) N. standleyi; 24) N. sulphurea; 25) Pellaea atropurpurea; 26) P. cordifolia; 27) P. ovata; 28) P. ternifolia var. ternifolia.")
Pteridofitas de las sierras El Sarnoso y Mapimí, Durango, México. 1) Selaginella lepidophylla; 2) S. pilifera; 3) S. rupincola; 4) Asplenium resiliens; 5) Pleopeltis polypodioides var. michauxianum; 6) Adiantum capillus-veneris; 7) Argyrochosma limitanea var. mexicana; 8) Astrolepis cochisensis; 9) A. integerrima; 10) A. sinuata; 11) Cheilanthes eatonii; 12) C. feei; 13) C. horridula; 14) C. lindheimeri; 15) C. notholaenoides; 16) C. tomentosa; 17) C. villosa; 18) C. leucopoda; 19) C. wrightii; 20) Notholaena aschenborniana; 21) N. grayi; 22) N. greggii; 23) N. standleyi; 24) N. sulphurea; 25) Pellaea atropurpurea; 26) P. cordifolia; 27) P. ovata; 28) P. ternifolia var. ternifolia.
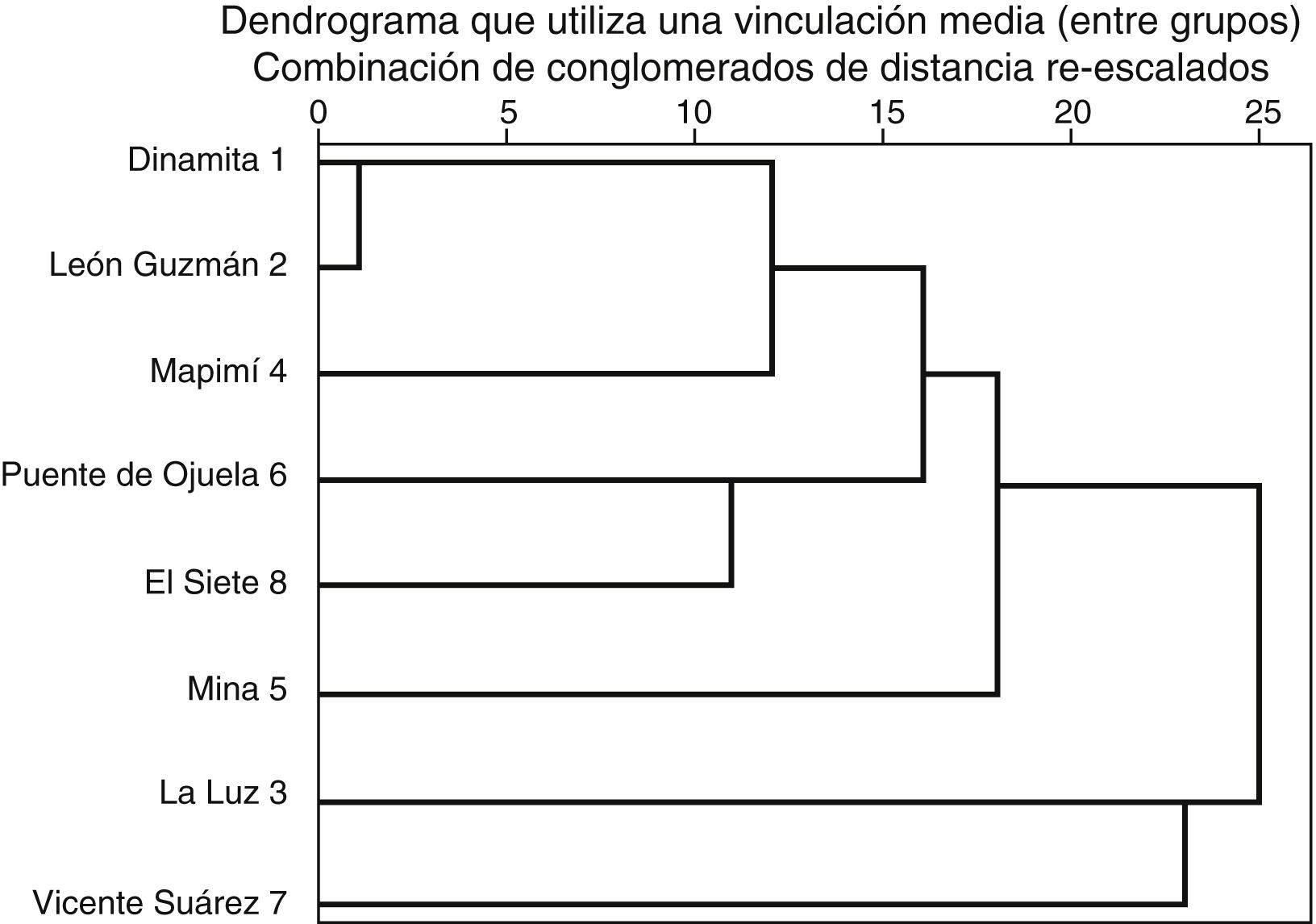
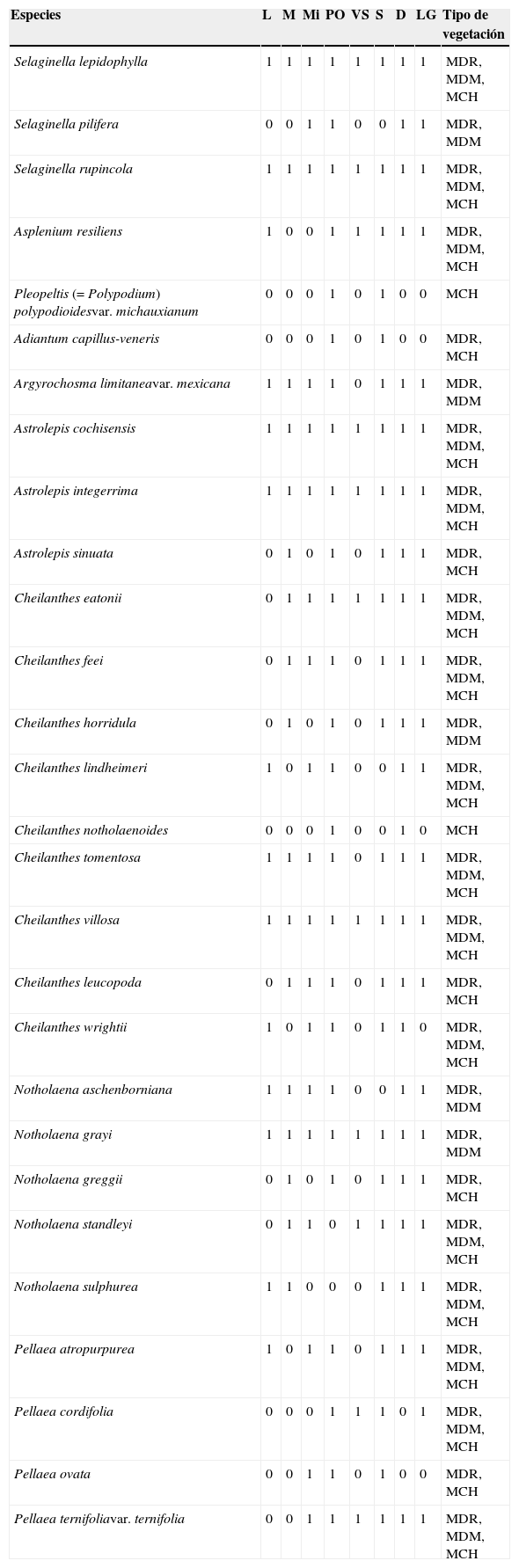
El índice de similitud de Jaccard entre la sierra El Sarnoso (Dinamita, La Luz, León Guzmán y Mina) y la sierra de Mapimí (Mapimí, Vicente Suárez, Puente de Ojuela y El Siete) fue de 0.89, es decir, que comparten más del 50% de las especies de pteridofitas. La localidad que presentó mayor número de especies fue Puente de Ojuela (28) y la que mostró menos fue Vicente Suárez (11). Las 28 especies citadas en el presente estudio se localizaron en la sierra de Mapimí, mientras que en la sierra El Sarnoso se localizaron 25 especies. En el cladograma obtenido mediante el análisis de conglomerados jerárquicos desarrollado a partir de la agrupación de especies por localidades, se observaron 7 grupos (fig. 4). Las localidades comprendidas por Dinamita y León Guzmán poseen una alta homogeneidad en cuanto a la diversidad de especies. Los tipos de vegetación presentes en estas localidades fueron el matorral desértico rosetófilo (MDR), el matorral desértico micrófilo (MDM) y el matorral chaparral (MCH) de acuerdo con Orona-Espino y Estrada-Rodríguez (2012); por su parte las localidades de Mapimí y La Mina presentaron una homogeneidad media en cuanto a diversidad de pteridofitas, encontrándose 3 tipos de vegetación en Mapimí (MDM, MDR y MCH) y solo 2 en La Mina (MDM y MDR). Finalmente, la localidad de Vicente Suárez y La Luz resultaron ser más heterogéneas en cuanto a la diversidad de especies que el resto de las localidades, registrándose solo el MDM en Vicente Suárez y MDR+MDM en la Luz (tabla 2).

Especies de pteridofitas por localidad y tipo de vegetación en que se observaron.
| Especies | L | M | Mi | PO | VS | S | D | LG | Tipo de vegetación |
|---|---|---|---|---|---|---|---|---|---|
| Selaginella lepidophylla | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Selaginella pilifera | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | MDR, MDM |
| Selaginella rupincola | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Asplenium resiliens | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Pleopeltis (=Polypodium) polypodioidesvar. michauxianum | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | MCH |
| Adiantum capillus-veneris | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | MDR, MCH |
| Argyrochosma limitaneavar. mexicana | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | MDR, MDM |
| Astrolepis cochisensis | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Astrolepis integerrima | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Astrolepis sinuata | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | MDR, MCH |
| Cheilanthes eatonii | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Cheilanthes feei | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | MDR, MDM, MCH |
| Cheilanthes horridula | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | MDR, MDM |
| Cheilanthes lindheimeri | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | MDR, MDM, MCH |
| Cheilanthes notholaenoides | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | MCH |
| Cheilanthes tomentosa | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | MDR, MDM, MCH |
| Cheilanthes villosa | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Cheilanthes leucopoda | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | MDR, MCH |
| Cheilanthes wrightii | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | MDR, MDM, MCH |
| Notholaena aschenborniana | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | MDR, MDM |
| Notholaena grayi | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM |
| Notholaena greggii | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | MDR, MCH |
| Notholaena standleyi | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
| Notholaena sulphurea | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | MDR, MDM, MCH |
| Pellaea atropurpurea | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | MDR, MDM, MCH |
| Pellaea cordifolia | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | MDR, MDM, MCH |
| Pellaea ovata | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | MDR, MCH |
| Pellaea ternifoliavar. ternifolia | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | MDR, MDM, MCH |
Localidades (D:Dinamita; L:La Luz; LG:León Guzmán; M:Mapimí; Mi:Mina; PO:Puente de Ojuela; S:El Siete; VS:Vicente Suárez); 0:ausente; 1:presente; MCH:Matorral chaparral; MDM:Matorral desértico micrófilo; MDR:Matorral desértico rosetófilo.
Las especies indicadoras proveen información cuantitativa sobre el estado de grandes sistemas (Hunsaker y Carpenter, 1990). En el presente estudio se pudo observar que en las instalaciones de caleras y marmoleras en la sierra de El Sarnoso, cuyas actividades han provocado la pérdida del hábitat tras el desmonte de la vegetación y la extracción de material como caliza y mármol, la planta P. ternifolia var. ternifolia actúa como pionera en la colonización de las áreas impactadas auxiliando en el inicio del proceso de sucesión vegetal primaria. Esto fue señalado por Rodríguez, Pacheco y Zavala-Hurtado (2008) en San Jerónimo Amanalco, Texcoco, Estado de México, quienes observaron bancos de esporas de esta especie en sitios modificados drásticamente por desmonte, incendio y áreas de cultivo abandonadas. Encontrar esta especie en las sierras El Sarnoso y Mapimí es consecuencia de las actividades antropogénicas inmoderadas que van produciendo una degradación progresiva del ambiente.
Los géneros con mayor número de especies en el presente trabajo fueron Cheilanthes (9) y Notholaena (6). Estrada-Castrillón y Villarreal-Quintanilla (2010) encontraron en el centro del estado de Chihuahua a estos mismos géneros (19 y 9 especies, respectivamente) en tipos de vegetación de desierto (matorral xerófilo), bosque de encinos, pastizales y bosque de pino. En las sierras El Sarnoso y Mapimí se registró un total de 4 familias, 9 géneros, 28 especies y 3 variedades de pteridofitas. En la sierra de Jimulco, Torreón, Coahuila (ubicada a 28km de las sierras El Sarnoso y Mapimí), Alba-Ávila (2011) registró 25 especies en vegetación de matorral (xerófilo, micrófilo y rosetófilo), chaparral y bosque de pino-encino, un piso altitudinal de 3,120 m snm y un área de 625 km2. Las sierras El Sarnoso y Mapimí presentan solo 3 tipos de vegetación (MDM, MDR y MCH) con elevaciones de 2,500 m snm y una superficie de 645.6 km2 (Dgetenal, 1978). Hunsaker y Carpenter (1990) y Whittaker, Willis y Field (2001) consideran que el número de especies tiende a incrementar conforme aumenta el tamaño del área.
Las sierras El Sarnoso y Mapimí comparten el 89% de las especies de pteridofitas según la similitud de Jaccard (0.89). Aunque Mapimí mostró una mayor riqueza de especies (28) que El Sarnoso (25) es posible que estos números se incrementen si se amplía el esfuerzo de muestreo. La similitud de especies entre ambas sierras se debe a que hace 65 millones de años constituían una sola formación rocosa, pero debido a un evento geológico quedaron separadas por el surgimiento del Cañón de las Piedras Encimadas, formándose así microhábitats muy similares. Esto explica por qué este grupo de plantas ocupan nichos ecológicos relativamente limitados en microhábitats en zonas áridas (Smith et al., 2006).
La especie Cheilanthes notholaenoides fue registrada en pocas localidades; sin embargo, Ramírez-Cruz et al. (2009) la documentaron en el Parque Nacional Los Mármoles, estado de Hidalgo como una especie de distribución amplia, registrándose en distintos tipos de bosques (encino, pino, pino-encino, táscate), matorral espinoso y vegetación secundaria. En el caso de las sierras El Sarnoso y Mapimí, esta especie se ha adaptado a crecer en grietas de las rocas donde se almacena el agua y existen los nutrientes necesarios para su desarrollo (Menéndez, Revilla, Bernard, Gotor y Fernández, 2006). La dispersión de sus esporas es anemocórica (por el viento), lo que facilita la colonización de otras áreas (Smith et al., 2006); además, algunas especies tienen frondas micrófilas, lo que les permite reducir la superficie de pérdida de agua (Blasco-Zumeta, 1998). Debido a que algunas especies de este grupo de plantas son indicadoras de deterioro ambiental; por ejemplo, P. ternifolia var. ternifolia, es importante considerarlas para monitorizar áreas vulnerables al impacto antropogénico. Asimismo, pueden marcar la pauta para comenzar a elaborar programas de conservación, en este caso para las sierras El Sarnoso y Mapimí, debido a que si no se controla la extracción de materiales rocosos, en un futuro se irá degradando el hábitat de estas sierras, lo que a su vez afectará la supervivencia de las especies de flora y fauna que habitan en ellas.
A E. Estrada-Castillón, J. A. Villarreal-Quintanilla, N. A. Salas-Muro, M. Armendáriz-Herrera, M. A. Favila, J. Bautista por su ayuda en el trabajo de campo. A M. A. Zavalza-Ávila por su ayuda en la elaboración de mapas. A Arreguín-Sánchez (ENCB), J. A. Villarreal-Quintanilla (UAAAN) y A. E. Estrada-Castillón (CFNL) por su apoyo en los herbarios. A C. W. Barrows por su ayuda en la revisión del abstract. Esta investigación fue desarrollada bajo el permiso de colecta SGPA/DGVS/08589/12 emitido por la Semarnat. La presente investigación fue financiada por el municipio de Lerdo, Durango, y comprende una parte del macro proyecto «Estudio técnico justificativo para declarar área natural protegida a la sierra El Sarnoso, municipio de Lerdo, Durango, México» Clave DGO-2008-CO4-96172.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.