



Se analizó la representatividad del inventario de plantas arbustivas de un área natural protegida de Coahuila, México, con los siguientes objetivos: describir los tipos de sesgos de muestreo; definir la heterogeneidad ambiental de la reserva captada en las recolectas botánicas, y determinar el nivel de resolución espacial que respalda la información recolectada. Los sesgos fueron significativos (por distancia a carreteras, χ2=199.01; p<0.01; por rango altitudinal, χ2=391.02; p<0.01, y por tipo de vegetación, χ2=518.13; p<0.01). La muestra del inventario exhibe diferencias significativas entre el grupo testigo y un diseño aleatorizado para 5 variables bioclimáticas (temperatura media anual [Bio1]: U=117,900; p<0.01; isotermalidad [Bio3]: U=189,650; p<0.01; precipitación del mes más húmedo [Bio13]: U=134,330; p<0.01; estacionalidad de la precipitación [Bio15]: U=175,720; p<0.01 y la precipitación del trimestre más frío [Bio19]: U=146,550; p<0.01). Respecto a las variables ambientales y distancias a carreteras, el cubrimiento de esta infraestructura dentro del área puede contribuir a una buena caracterización del área natural protegida. La información proveniente de las recolectas realizadas es adecuada para escalas pequeñas de análisis (1 y 0.5°).
The representativeness of the inventory of shrub species of a natural protected area in Coahuila, Mexico was analyzed, with the following objectives: to describe the types of sampling bias; to define the environmental heterogeneity of the botanical records; to determine the level of spatial resolution that supports the information collected. Biases were significant (χ2=distance road 199.01; p<0.01, by altitudinal range χ2=391.02; p<0.01 and vegetation χ2=518.13; p<0.01). The inventory sample exhibits significant differences between the control group and a randomized design for 5 bioclimatic variables (mean annual temperature (Bio1): U=117,900; p<0.01; isothermality (Bio3): U=189.650, p<0.01; precipitation wettest month (Bio13): U=134.330, p<0.01; seasonality of precipitation (Bio15): U=175.720, p<0.01 and precipitation of the coldest quarter (Bio19): U=146.550, p<0.01). Environmental variables and road distances, on the coverage of this infrastructure in the area, can contribute to a good characterization of the ANP. The information from collected records is adequate for small scales of analysis (1 and 0.5°).
Una de las premisas fundamentales para los administradores de los recursos naturales y la biodiversidad es determinar el universo de su ámbito de trabajo. De esta manera, una de las tareas más importantes de los biólogos es recolectar, organizar y sistematizar el conocimiento botánico a través de los inventarios biológicos (Stork y Samways, 1995), los cuales se reconocen como la forma más generalizada para documentar la diversidad de especies. La información contenida en las colecciones posee un importante acervo de nuestro patrimonio biológico (Navarro, Peterson, Nakazawa y Liebig-Fossas, 2003), y reviste un gran valor para generar información relevante, principalmente cuando se utiliza en combinación con otras fuentes de datos y con enfoques analíticos emergentes (Martínez-Meyer y Sánchez-Cordero, 2006). Por otra parte, los datos contenidos pueden ayudarnos a describir aspectos sobre la caracterización biogeográfica y ecológica de las especies (Morrone, 2000; Salinas-Rodríguez, Estrada-Castillón y Villarreal-Quintanilla, 2013), así como el análisis de los patrones de distribución y riqueza biológica (Estrada-Castillón et al., 2010; Soberón y Peterson, 2004; Velazco-Macías, Alanís-Flores, Alvarado-Vázquez, Ramírez-Freire y Foroughbakhch-Pournavab, 2011), además de cobrar un especial interés en la planeación sistemática de la conservación y la protección de especies y sus poblaciones (Alanís-Flores, Velazco-Macías, Foroughbakhch, Valdez y Alvarado, 2004; Arzamendia y Giraudo, 2004).
México cuenta con una importante tradición taxonómica, dado que la sistematización de datos relativos a la biodiversidad mexicana, y en particular de la flora, data desde el quinquenio de 1753-1757, period en que destacan las contribuciones de Francisco Javier Clavijero y José Antonio Alzate (Llorente-Bousquets y Ocegueda, 2008). Es interesante observar que el impulso más fuerte de la recolecta de datos florísticos para el noreste de México se ha dado desde los años 1980 al 2000 (Velazco-Macías, 2009). Por otra parte, en las últimas décadas, la pérdida de la biodiversidad se ha considerado como uno de los temas más importantes dentro de las diversas agendas de los distintos niveles de gobierno, como consecuencia del impacto antrópico directo (sobreexplotación) o indirecto (pérdida y alteración del hábitat) (Halffter y Moreno, 2005). Por tal razón, se han dirigido esfuerzos significativos a la generación de programas para su conservación, entre los que destacan el desarrollo y manejo de áreas naturales protegidas en sus distintas modalidades (Cantú, Wright, Scott y Strand, 2004; Castaño-Villa, 2005).
Considerando a las especies y subsecuentemente a los inventarios biológicos como uno de los insumos principales utilizados para la definición y selección de áreas prioritarias (Halffter y Ezcurra, 1992; Kress et al., 1998), resulta importante conocer la representatividad de los esfuerzos de recolecta para determinar los sesgos que potencialmente puedan limitar la definición de patrones geográficos de la biodiversidad (Hijmans et al., 2000; Hortal, Lobo y Jiménez-Valverde, 2007; Soberón, Llorente y Oñate, 2000) y, por ende, limitar su utilidad para el desarrollo de estas estrategias para la gestión del territorio natural (Hortal y Lobo, 2006; Tobler, Honorio, Janovec y Reynel, 2007). En este sentido, se han desarrollado varios enfoques empíricos para evaluar la fortaleza de los inventarios y las bases de datos sobre biodiversidad, y se han llevado a cabo evaluaciones en términos estadísticos con apoyo de estimaciones de carácter paramétrico y no paramétrico (Colwell y Coddington, 1994; Chadzon, Colwell, Denslow y Guariguata, 1998; López-Gómez y Williams-Linera, 2006), y a distintas escalas geográficas (Villaseñor, Maeda, Colín-López y Ortiz, 2005). Sin embargo, un aspecto fundamental en la evaluación de las colecciones y bases de datos curatoriales es la caracterización de estos registros en términos de su representatividad ambiental, dado que la presencia de los organismos con relación a sus variables ambientales proporciona información vital para describir su nicho ecológico fundamental existente y aproximarse a su nicho realizado (Grinell, 1917; Hortal, Jiménez-Valverde, Gómez, Lobo y Basealga, 2008; Soberón, 2007).
Para abordar esta cuestión, se evaluaron las recolectas botánicas del Área de Protección de Recursos Naturales Cuenca Alimentadora del Distrito Nacional de Riego 004 Don Martín (APRN CADDR 004), en lo respectivo a la porción río Sabinas, La Encantada, Santa Rosa y El Burro (Conanp, 2014), por ser una de las reservas más extensas dentro del noreste de México y revestir gran importancia para la conservación por distintos esquemas de regionalización establecidos (Arriaga et al., 2000; Benítez, Arizmendi y Márquez, 1999). Por otra parte, como grupo botánico para este análisis, se seleccionaron 91 especies de plantas arbustivas, por ser un conjunto representativo en términos de estructura y diversidad, dentro de los paisajes regionales del noreste mexicano (Canizales-Velázquez et al., 2009; González-Rodríguez, Ramírez-Lozano, Cantú-Silva, Gómez-Meza y Uvalle-Sauceda, 2010; Mora-Donjuán et al., 2013), y además, porque varias de sus especies constituyen un recurso fitogenético utilizado con frecuencia por las distintas comunidades humanas en la herbolaria tradicional, como elementos dendroenergéticos (obtención de leña y carbón), como forraje para el ganado, para la elaboración de artesanías, para la fabricación de muebles y herramientas, y como especies ornamentales (Estrada-Castillón, Yen-Méndez, Delgado-Salinas y Villarreal-Quintanilla, 2004; Ramos-Silva et al., 2007). Esta situación ha favorecido que este grupo se encuentre bien representado en las colecciones biológicas y los inventarios regionales del noreste de México (Estrada-Castillón et al., 2004; Estrada-Castillón, Villarreal-Quintanilla y Jurado, 2005; Villarreal-Quintanilla, 2001).
Con este trabajo se busca describir la pertinencia del uso de los datos provenientes de recolectas en el área para el análisis de patrones de distribución, y su aplicación para el uso en la toma de decisiones para la conservación, para lo cual consideramos 3 objetivos principales: a) describir los tipos de sesgos de muestreo que estos datos representan (sesgo de distancia a carretera, altitud y tipos de vegetación); b) definir qué tanto se capta de la heterogeneidad ambiental de la reserva en cuestión, en las recolectas previamente desarrolladas, y c) determinar el nivel de resolución espacial que puede soportar la información previamente recolectada (efecto de la escala sobre los datos recolectados). Todo ello con la intención de poder validar esta información y, en su defecto, proponer recomendaciones que permitan optimizar su uso, para describir patrones y procesos en aras de la conservación de la biodiversidad de esta reserva natural.
Materiales y métodosEl APRN CADDR 004 fue decretada en el año de 2002 (Semarnat, 2002). En particular el polígono correspondiente a la porción río Sabinas, La Encantada, Santa Rosa y serranías del Burro. Se localiza en la parte centro noreste del estado de Coahuila y abarca una superficie de 8,024.53 km2, que representa cerca del 10% de la superficie estatal. Pertenece a la provincia fisiográfica “Grandes Llanuras de Norteamérica”, caracterizada por presentar climas secos y semisecos con temperaturas mínimas que suceden a mediados del invierno, llegando a registrarse hasta –16°C. Los valores extremos de temperatura máxima oscilan entre 43 y 55°C, en los meses del verano (junio-septiembre), mientras que la precipitación ocurre en el rango de los 300 a los 600mm anuales. La altitud presente dentro del área estudiada oscila entre los 265 hasta 2,203 m snm. Por su parte, la edafología del área se caracteriza principalmente por presentar tipos de suelos xerosol, regosol y vertisol. Geológicamente se encuentran rocas sedimentarias de origen marino y en segundo plano rocas magmáticas del tipo basáltico (Inegi, 1983). La reserva forma parte de la provincia Tamaulipeca (Morrone, 2005), la cual constituye una amplia zona de ecotonía entre la Planicie Costera del Golfo y el Altiplano Mexicano, que constituye un importante centro de endemismos (Rojas-Mendoza, 1965; Rzedowski, 1978). Su vegetación está formada en su mayor parte por matorrales xerófilos, entre los cuales destacan el matorral espinoso tamaulipeco y el matorral micrófilo; también se presenta vegetación riparia, pastizales, matorral submontano y encinares (ICE-FCF, 2004).
Para el análisis del nivel del inventario de plantas arbustivas, se recopilaron 681 registros, correspondientes a la información de 91 especies, 57 géneros y 27 familias (tabla 1), a partir de recolectas y observaciones de campo de los autores, en literatura especializada, así como de registros provenientes de bases de datos digitales de la Red Mundial de Información sobre Biodiversidad (REMIB, 2014) y el sitio de Global Biodiversity Information Facility (GBIF, 2014). Se utilizó el sistema taxonómico propuesto por The Angiosperm Phylogeny Group (2009) y para la revisión de las autoridades de las especies se utilizaron The Plant List (2010), The International Plant Names Index (2012) y el International Code of Botanical Nomenclature (Vienna Code, 2006). De la base de datos resultante se eliminaron los registros que presentaban inconsistencias geográficas (falta de claridad en la descripción de la localidad); de igual forma, se descartaron los registros que presentaron imprecisiones en la determinación taxonómica, como por ejemplo, la especie del género no determinada. Toda la información se administró para su despliegue dentro de sistemas de información geográfica. Se consideró como marco para la recopilación de los registros de estas especies el área comprendida por el polígono del Área Natural Protegida Río Sabinas, Coahuila (fig. 1).
Listado de plantas arbustivas y número de registros para cada especie que comprende el presente estudio.
| Familia | Especie | Número de registros |
|---|---|---|
| Amaranthaceae | Atriplex canescens (Pursh) Nutt. | 1 |
| Pistacia mexicana Kunth | 1 | |
| Rhus aromatica Aiton | 1 | |
| Rhus glabra L. | 1 | |
| Rhus lanceolata (A.Gray) Britton | 6 | |
| Rhus virens Lindh. ex A. Gray var. virens Lindh. ex A. Gray | 12 | |
| Asteraceae | Baccharis neglecta Britton | 4 |
| Flourensia cernua DC. | 1 | |
| Gochnatia hypoleuca (DC.) A. Gray subsp. hypoleuca | 3 | |
| Gutierrezia sarothrae (Pursh) Britton y Rusby | 1 | |
| Gutierrezia texana (DC.) Torr. y A. Gray var. glutinosa (S. Schauer) M. A. Lane | 2 | |
| Gymnosperma glutinosum (Spreng.) Less. | 3 | |
| Berberidaceae | Berberis trifoliolata Moric. | 1 |
| Bignoniaceae | Chilopsis linearis (Cav.) Sweet subsp. arcuata (Fosberg) Henr. | 1 |
| Cannabaceae | Celtis laevigata Willd. | 2 |
| Celtis ehrenbergiana (Klotzsch) Liebm. | 14 | |
| Celastraceae | Schaefferia cuneifolia A. Gray. | 3 |
| Ebenaceae | Diospyros texana Scheele | 11 |
| Ehretiaceae | Ehretia anacua (Terán y Berland.) I. M. Johnst | 1 |
| Euphorbiaceae | Croton cortesianus Kunth | 1 |
| Croton dioicus Cav. | 4 | |
| Croton fruticulosus Torr. | 7 | |
| Croton incanus Kunth | 5 | |
| Croton leucophyllus Müll.Arg. | 2 | |
| Croton pottsii (Klotzsch) Müll.Arg. | 1 | |
| Croton suaveolens Torr. | 1 | |
| Manihot subspicata D. J. Rogers y Appan | 1 | |
| Fabaceae | Acaciella angustissima (Mill.) Britton y Rose var. texensis (Torr. y A. Gray) L. Rico | 6 |
| Bauhinia lunarioides A. Gray ex S. Watson | 6 | |
| Bauhinia macranthera Benth ex Hemsl. | 5 | |
| Calliandra eriophylla Benth. | 2 | |
| Cercis canadensis L. | 3 | |
| Chamaecrista greggii (A. Gray) A. Heller | 10 | |
| Dermatophyllum secundiflorum (Ort.) Gandhi y Reveal | 17 | |
| Eysenhardtia texana Scheele | 20 | |
| Havardia pallens (Benth.) Britton y Rose | 6 | |
| Mariosousa coulteri (Benth.) Seigler y Ebinger | 7 | |
| Mimosa aculeaticarpa var. biuncifera (Benth.) Barneby | 16 | |
| Parkinsonia aculeata L. | 11 | |
| Parkinsonia texana (A. Gray) S. Watson var. macra (I. M. Johnst.) Isely | 17 | |
| Prosopis glandulosa Torr. var. torreyana (L. D. Benson) M. C. Johnst. Torr. | 23 | |
| Prosopis reptans Benth. | 1 | |
| Senegalia berlandieri (Benth.) Britton y Rose | 31 | |
| Senegalia greggii (A. Gray) Britton y Rose | 54 | |
| Senegalia roemeriana (Scheele) Britton y Rose | 6 | |
| Senna wislizeni (A. Gray) H. S. Irwin y Barneby var. painteri (Britton y Rose) H. S. Irwin y Barneby | 2 | |
| Vachellia farnesiana (L.) Wight y Arn. | 62 | |
| Vachellia glandulifera (S. Watson) Seigler y Ebinger | 3 | |
| Vachellia rigidula (Benth.) Seigler y Ebinger | 72 | |
| Vachellia schaffneri (S. Watson) Seigler y Ebinger | 5 | |
| Vachellia vernicosa (Britton y Rose) Seigler y Ebinger | 7 | |
| Zapoteca media (M. Martens y Galeotti) H. M. Hern. | 2 | |
| Fagaceae | Quercus invaginata Trel. | 1 |
| Garryaceae | Garrya ovata Benth. var. lindehimeri (Torr.) J. M. Coult. y W. H. Evans | 5 |
| Koeberliniaceae | Koeberlinia spinosa Zucc. | 2 |
| Krameriaceae | Krameria bicolor S. Watson | 2 |
| Lamiaceae | Poliomintha glabrescens A. Gray ex Hemsl. | 7 |
| Salvia ballotiflora Benth. | 4 | |
| Oleaceae | Forestiera angustifolia Torr. | 4 |
| Forestiera reticulata Torr. | 1 | |
| Rhamnaceae | Ceanothus caeruleus Lag. | 1 |
| Ceanothus greggii A. Gray | 5 | |
| Ceanothus herbaceus Raf. | 7 | |
| Colubrina greggii S. Watson | 8 | |
| Colubrina stricta Engelm. ex M. C. Johnst. | 9 | |
| Condalia ericoides (Cav.) Sweet | 1 | |
| Condalia hookeri M. C. Johnst. | 1 | |
| Condalia mexicana Schltdl. | 1 | |
| Karwinskia humboldtiana (Schult.) Zucc. | 5 | |
| Ziziphus obtusifolia (Hook. ex Torr. y A. Gray) A. Gray | 9 | |
| Rosaceae | Cercocarpus fothergilloides Kunth | 3 |
| Cercocarpus montanus Raf. | 7 | |
| Crataegus baroussana Eggl. | 1 | |
| Crataegus greggiana Eggl. | 5 | |
| Crataegus johnstonii J. B.Phipps | 1 | |
| Prunus serotina subsp. virens (Woot. y Standl.) McVaugh. | 3 | |
| Rubiaceae | Cephalanthus occidentalis L. | 1 |
| Rutaceae | Zanthoxylum fagara (L.) Sarg. | 13 |
| Sapindaceae | Ungnadia speciosa Endl. | 6 |
| Scrophulariaceae | Leucophyllum frutescens (Berland.) I. M. Johnst. | 15 |
| Simaroubaceae | Castela erecta Liebm. | 9 |
| Solanaceae | Lycium berlandieri Dunal var. berlandieri | 1 |
| Verbenaceae | Aloysia gratissima (Gillies y Hook.) Tronc. | 6 |
| Aloysia macrostachya (Torr.) Moldenke | 2 | |
| Lantana achyrantifolia Desf. | 8 | |
| Lantana camara L. | 4 | |
| Lantana macropoda Torr. | 10 | |
| Lantana urticoides Hayek | 6 | |
| Lippia graveolens Kunth | 2 | |
| Zygophyllaceae | Guaiacum angustifolium Engelm. | 16 |
| Larrea tridentata (Sessé y Moc. ex DC.) Coville | 2 |
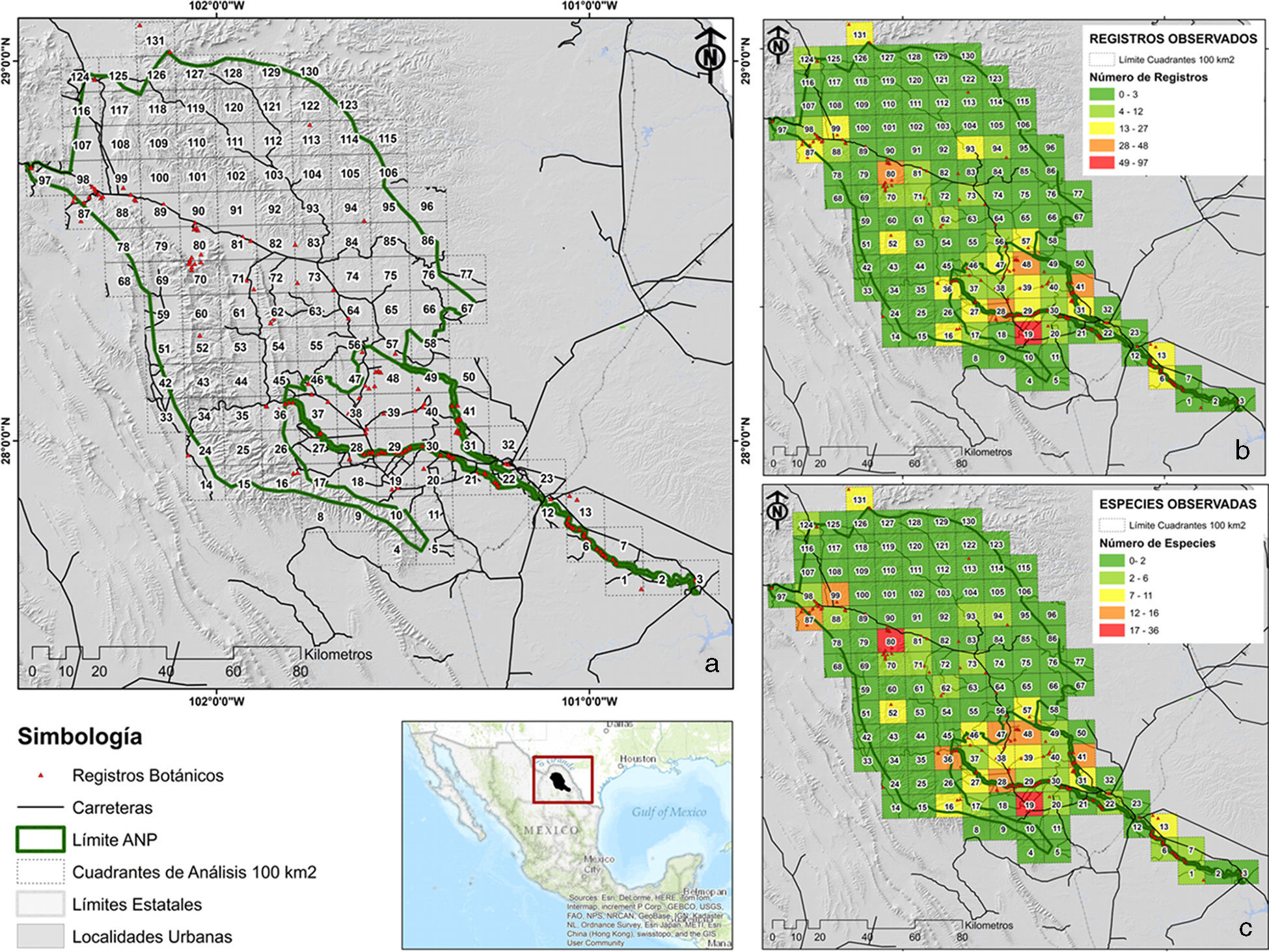
 Localización del área de estudio y cuadrantes de análisis de la información (100 km2); b) número de registros observados para cada uno de los 131 cuadrantes de 100 km2 analizados; c) número de especies observadas para cada uno de los 131 cuadrantes de 100 km2 analizados.")
El área correspondiente al polígono del APRN CADDR 004 y en específico a la porción río Sabinas, La Encantada, Santa Rosa y serranías del Burro (8,024.53 km2), se dividió en cuadrantes de 10×10km (∼100 km2), y se consideró que esta superficie era la adecuada por el nivel de resolución espacial de la información; además, porque la escala de trabajo seleccionada puede aportar información que ayude a tomar decisiones sobre dónde invertir recursos para la conservación e identificar áreas clave útiles para evaluaciones posteriores a escalas más finas (Conabio, Conanp, TNC, Pronatura, FCF, UANL, 2007). Del área dividida resultaron 131 cuadrantes, y a cada uno de ellos se les asignó un identificador numérico individual. Los registros geográficos de las especies se desplegaron sobre el área y, posteriormente, se cuantificó los números de registros y de especies para cada cuadrante (fig. 1).
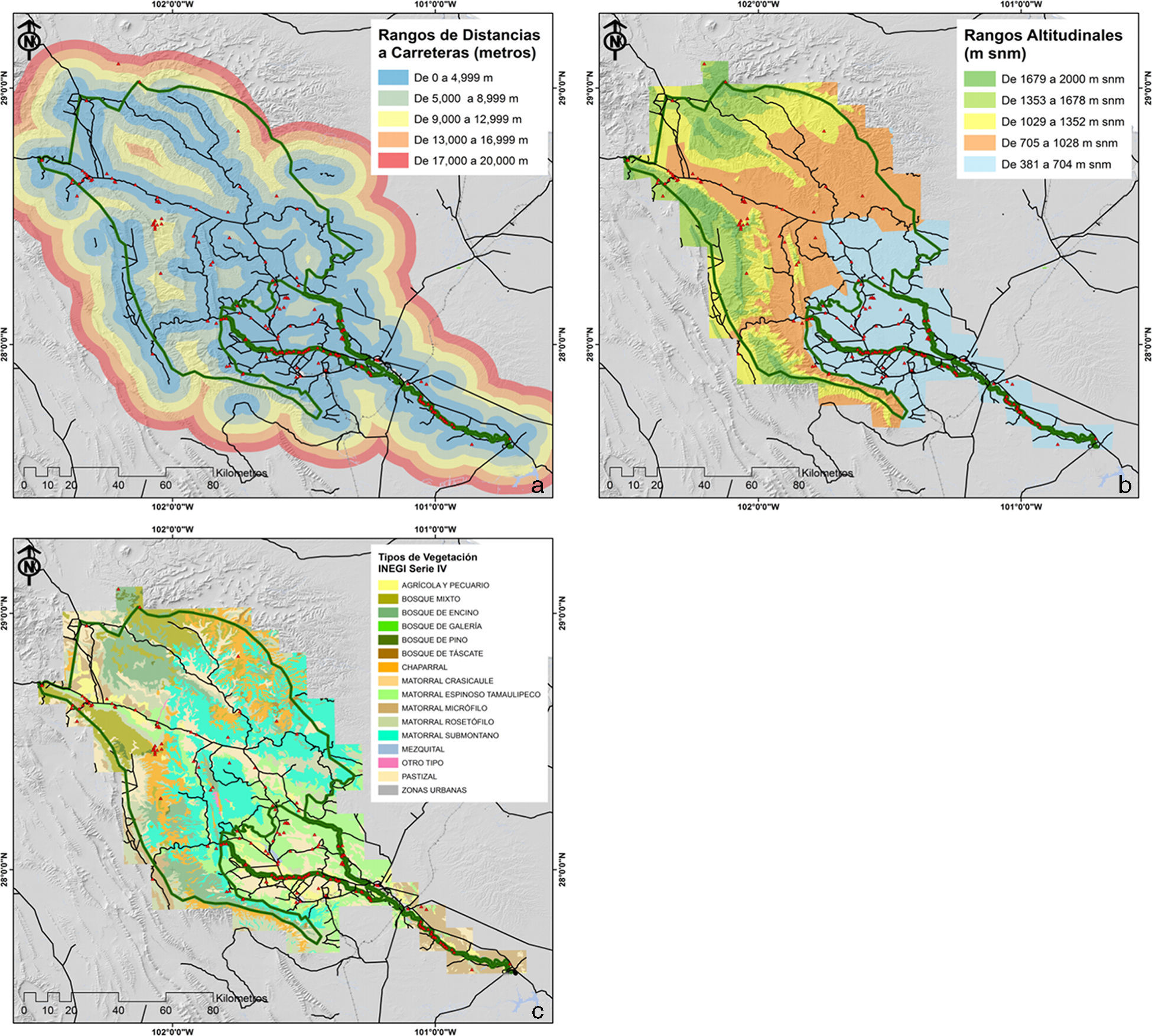
Para el análisis de sesgos de distancia a carreteras, se generó una matriz de distancia a la carretera a partir de la capa de vías de comunicación principales escala 1:250,000 proveniente del Continuo Nacional de Topografía serie II de Inegi (1990). Se desarrollaron 5 clases de distancia (de 0 a 4,999m; de 5,000 a 8,999m; de 9,000 a 12,999m, de 13,000 a 16,999m y de 17,000 a 20,000m), las cuales fueron calculadas a partir de la distancia máxima (16km) documentada para los registros recolectados en el área de estudio. Posteriormente, los registros puntuales (n=681) fueron vertidos sobre la capa generada y se determinó el número de registros y especies presentes para cada rango de distancias a carreteras. Estos datos se compararon contra una matriz de puntos completamente aleatoria (n=681), mediante una prueba de χ2 (fig. 2a).
 distancia a carreteras; b) rangos altitudinales, y c) vegetación dentro del área de estudio.")
De igual forma, a partir de las curvas de nivel escala 1:250,000 (Conabio, 1998), se generó un modelo digital de elevación, del cual se definieron 5 rangos altitudinales (de 381 a 705m snm, de 706 a 1,028m snm, de 1,029 a 1,352m snm, de 1,353 a 1,678m snm y de 1,679 a 2,000m snm), dentro de los cuales se virtieron los registros de plantas arbustivas del área (n=681 registros), para luego describir el número de registros y especies contenidos para cada rango analizado. La matriz aleatoria de puntos, previamente generada (n=681 registros), se comparó contra los registros y rangos altitudinales anteriormente descritos a través de una prueba de χ2 (fig. 2b).
Para comparar los registros previamente recolectados, contra un diseño aleatorizado estratificado, se consideraron los tipos agrupados de vegetación de la serie IV de vegetación (Inegi, 2007), y se determinó con base en su extensión, los estratos y sus proporciones respectivas. Así, se asignó una muestra proporcional de registros para cada uno de los estratos (tipos de vegetación) y se compararon estadísticamente mediante una prueba de χ2 contra los registros previamente recolectados en el área (fig. 2c).
Para analizar la representatividad ambiental del inventario del área protegida, se utilizó un total de 5 de las 19 variables climáticas de la base de datos de WorldClim (Hijmans, Cameron, Parra, Jones y Jarvis, 2005). Para evitar la redundancia en la información, se discriminaron aquellas variables que presentaron valores iguales o mayores al 0.70 de correlación de rangos de Spearman, con una p<0.01. Este umbral elegido describe relaciones fuertes entre los parámetros evaluados, lo que reduce adecuadamente el número de variables que utilizar. De esa manera, se trabajó con las capas de temperatura media anual (Bio1); isotermalidad (Bio3); precipitación del mes más húmedo (Bio13); temperatura máxima del mes más cálido (Bio15), y precipitación del trimestre más frío (Bio19). Respecto a la información botánica, se consideró la totalidad de las especies y registros documentados para el área (n=91 y 681 registros, respectivamente), además, se utilizó el conjunto aleatorizado con igual número de réplicas previamente elaborado (n=681). Para cada registro geográfico de ambos conjuntos, se asignó el valor de cada una de las 5 variables seleccionadas, a través de procesos de análisis espacial (SDMtoolbox v.1 para ArcGis 10.x; http://sdmtoolbox.org/). Para la comparación de estos datos se utilizó el test no paramétrico de Mann-Whitney incluido dentro del programa PAST 3.01 (Hammer, Harper y Ryan, 2001), dado que las variables climáticas utilizadas no exhibieron normalidad en los datos.
Asimismo, con el propósito de analizar el efecto del sesgo de distancia a carretera, sobre la descripción de las condiciones ambientales de las especies en el área, se agruparon los registros observados y aleatorizados, considerando los 5 rangos de distancia definidos (de 0 a 4,999m; de 5,000 a 8,999m, de 9,000 a 12,999m, de 13,000 a 16,999m y de 17,000 a 20,000m). Se buscaron diferencias estadísticas para cada rango de distancia y para cada variable a través de la prueba de Mann-Whitney.
Para analizar el efecto de la escala sobre el grado de conocimiento del inventario de plantas arbustivas, se definieron 4 particiones geográficas de 1×1° (4 cuadrantes con una superficie de ∼10,908.30 km2); 0.5×0.5° (11 cuadrantes con superficies aproximadas de 2,734.05km2); 0.25×0.25° (30 cuadrantes con superficies de ∼ 683 km2) y 0.10×0.10° (131 cuadrantes de ∼100 km2) de resolución espacial. Para cada una de las 4 escalas de análisis definidas (1, 0.5, 0.25 y 0.1°) se estimó la riqueza potencial mediante el estimador no paramétrico Chao 2, el cual se ajusta a la naturaleza de los datos (incidencia) y se encuentra disponible dentro del programa EstimateS 9.1.0. (Colwell, 2013):
donde SOBS es el número total de especies observadas en toda las muestras llevadas al pool, y Q1, el número de especies que se presentan en exactamente una muestra (Q1 es la frecuencia de especies únicas, Q2 la frecuencia de especies duplicadas). Por último, se determinó el nivel de completitud (Moreno y Halffter, 2000) para cada una de las 4 escalas geográficas, y se comparó de forma cuantitativa cada valor obtenido:donde ICi es el índice de completitud, S.OBS, las especies observadas para el área de análisis y S.EST, las especies estimadas para el área de análisis por el modelo de Chao 2.Resultados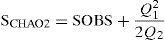
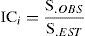
De las 91 especies documentadas, el número de registros osciló entre 1 y 72, con un promedio de 7 registros por especie. El taxón con mayor número de registros fue Vachellia rigidula (n=72), seguida de Vachellia farnesiana (n=62) y Senegalia greggii (n=54). En contraparte, 24 especies solamente se documentan con 1 registro dentro del área de estudio, mientras que 10 especies presentaron solamente 2 registros (Celtis laevigata, Croton leucophyllus, Calliandra eriophylla, Senna wislizeni var. painteri, Zapoteca media, Koeberlinia spinosa, Krameria bicolor, Aloysia macrostachya, Lippia graveolens y Larrea tridentata). En cuanto a los cuadrantes, se observó un total de 52 cuadrantes con al menos 1 registro, con un promedio de 13.09 y un rango entre 1 y 97 registros por cuadrante; por otro lado, 79 cuadrantes no cuentan con información botánica (fig. 1b). Respecto al número de especies por cuadrante, se observó un máximo de 36 (cuadrante 19), con un mínimo de 1 especie y un promedio de 6.84 especies por cuadrante. Geográficamente, la mayor proporción de registros y especies se presentó para la parte centro y sureste del área protegida (fig. 1c).
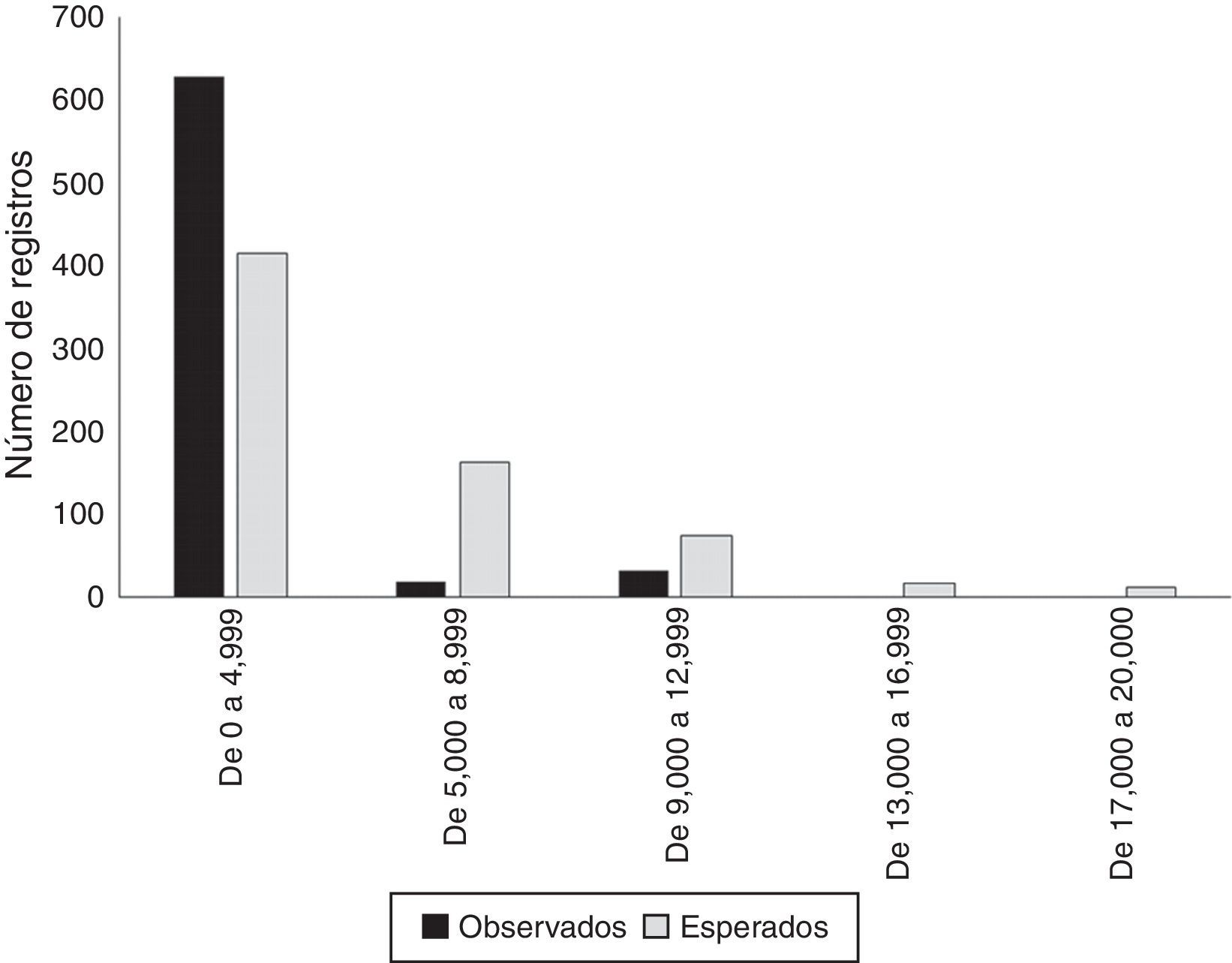
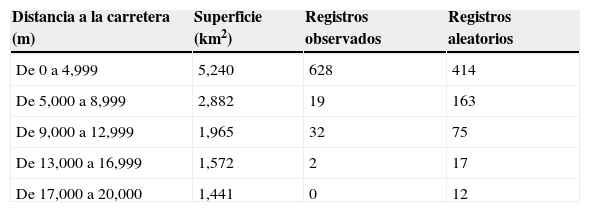
La mayor proporción (92.21%) de las recolectas y observaciones sobre plantas arbustivas en el área se presentó en los primeros 4,000m a partir de la infraestructura de caminos existente (tabla 2; fig. 2a; fig. 3). Por su parte, las clases posteriores a este rango de distancia, se encuentran subrepresentadas, en términos de registros esperados, con una diferencia significativa respecto a la muestra observada en los rangos de distancia a vías de comunicación (χ2=199.01; p<0.01).(figs. 4 y 5)
Rangos de distancia a carreteras y número registros observados y esperados del inventario de plantas arbustivas.
| Distancia a la carretera (m) | Superficie (km2) | Registros observados | Registros aleatorios |
|---|---|---|---|
| De 0 a 4,999 | 5,240 | 628 | 414 |
| De 5,000 a 8,999 | 2,882 | 19 | 163 |
| De 9,000 a 12,999 | 1,965 | 32 | 75 |
| De 13,000 a 16,999 | 1,572 | 2 | 17 |
| De 17,000 a 20,000 | 1,441 | 0 | 12 |
 y número de registros completamente al azar (barra gris), considerando los distintos rangos de distancia a vías de comunicación dentro del área de estudio.")
 y número de registros completamente al azar (barra gris), considerando los distintos rangos altitudinales presentes dentro del área de estudio.")
 y esperados (barra gris) para los distintos tipos de vegetación presentes en el área de estudio.")
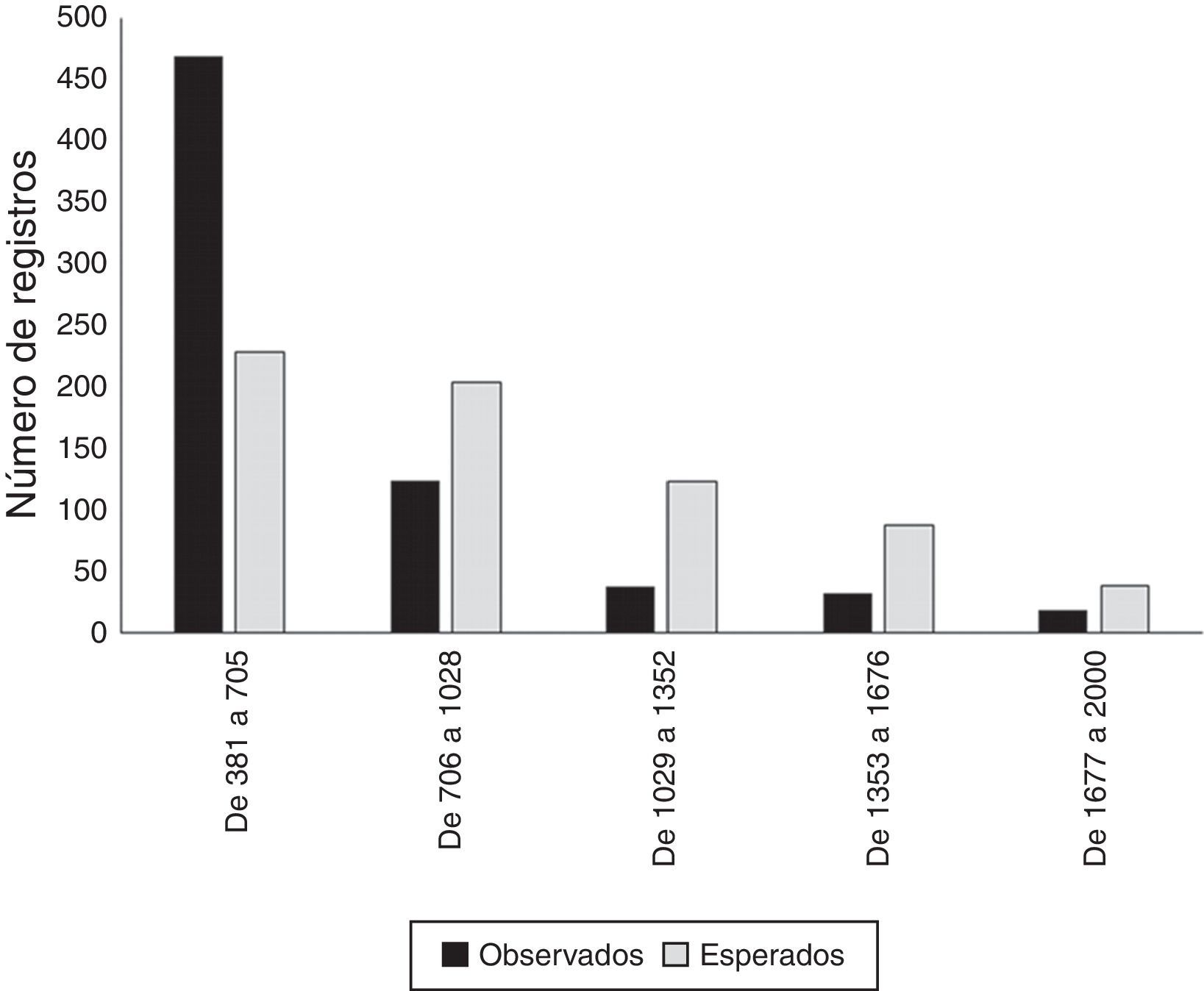
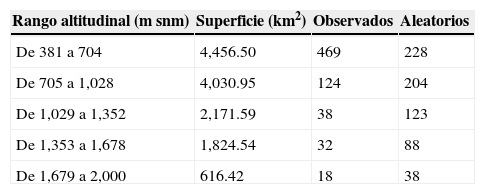
Se presentó una diferencia significativa entre los registros observados y los esperados para cada rango de altitud, dentro del área de estudio (χ2=391.02; p<0.01). Se observa que el 68% de las recolectas se presentaron dentro del rango altitudinal entre los 381 a 705m snm. Considerando a la muestra aleatoria para comparación, las recolectas y observaciones desarrolladas superan en un 205% la cantidad esperada de registros dentro de este rango. Por su parte, los subsecuentes rangos altitudinales se encuentran con un déficit de muestras que oscilan entre el 30.89% (rango altitudinal de 1,029 a 1,352m snm) y el 60.78% (rango altitudinal de 705 a 1,028m snm) (tabla 3, figs. 2c y 4).
Distribución de los registros observados y esperados de plantas arbustivas para los distintos rangos altitudinales presentes en el área de estudio.
| Rango altitudinal (m snm) | Superficie (km2) | Observados | Aleatorios |
|---|---|---|---|
| De 381 a 704 | 4,456.50 | 469 | 228 |
| De 705 a 1,028 | 4,030.95 | 124 | 204 |
| De 1,029 a 1,352 | 2,171.59 | 38 | 123 |
| De 1,353 a 1,678 | 1,824.54 | 32 | 88 |
| De 1,679 a 2,000 | 616.42 | 18 | 38 |
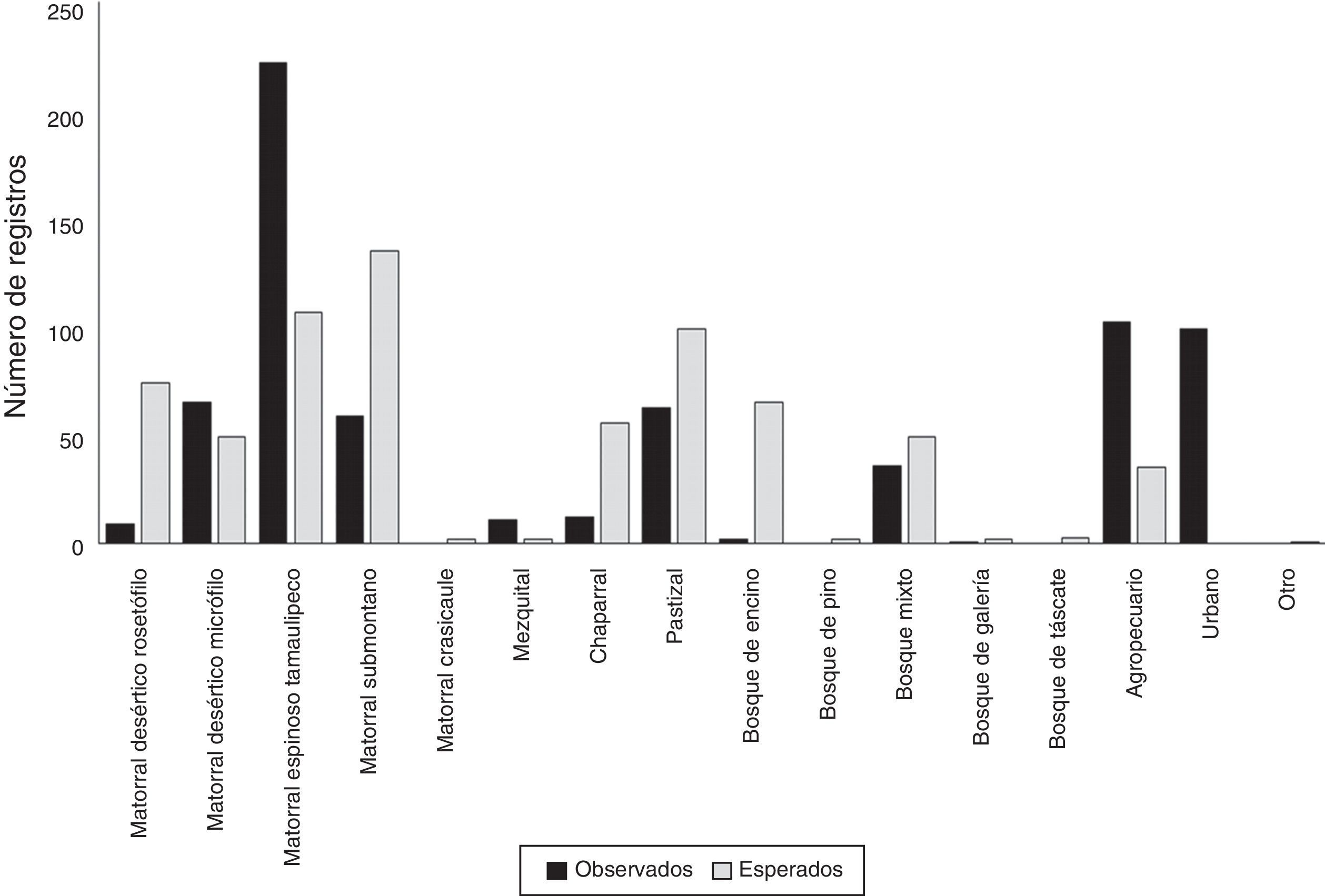
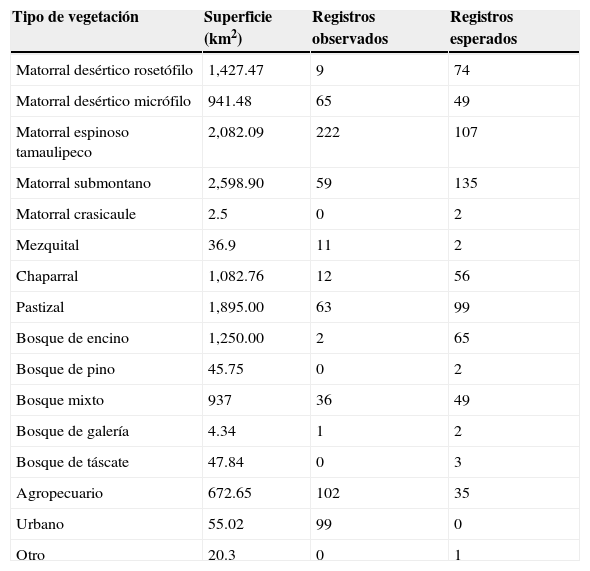
El mayor número de registros observados se presentó para el matorral espinoso tamaulipeco, el cual se encuentra sobrerrepresentado en un 207%, comparativamente con un diseño aleatorizado estratificado. De igual manera, las categorías de uso de suelo y vegetación del tipo agropecuario, matorral desértico micrófilo y mezquital fueron muestreadas con mayor intensidad a lo esperado (291.43; 132.65; 550.00%, respectivamente). Por otra parte, las restantes 12 categorías de vegetación resultaron subrepresentadas acorde con la cantidad de registros esperados mediante un muestreo aleatorizado estratificado (tabla 4; figs. 2b y 5). De esa forma, se observó una diferencia significativa entre los registros observados contra los esperados (χ2=518.13; p<0.01).
Distribución de los registros observados y esperados de plantas arbustivas para los distintos tipos de vegetación presentes en el área de estudio.
| Tipo de vegetación | Superficie (km2) | Registros observados | Registros esperados |
|---|---|---|---|
| Matorral desértico rosetófilo | 1,427.47 | 9 | 74 |
| Matorral desértico micrófilo | 941.48 | 65 | 49 |
| Matorral espinoso tamaulipeco | 2,082.09 | 222 | 107 |
| Matorral submontano | 2,598.90 | 59 | 135 |
| Matorral crasicaule | 2.5 | 0 | 2 |
| Mezquital | 36.9 | 11 | 2 |
| Chaparral | 1,082.76 | 12 | 56 |
| Pastizal | 1,895.00 | 63 | 99 |
| Bosque de encino | 1,250.00 | 2 | 65 |
| Bosque de pino | 45.75 | 0 | 2 |
| Bosque mixto | 937 | 36 | 49 |
| Bosque de galería | 4.34 | 1 | 2 |
| Bosque de táscate | 47.84 | 0 | 3 |
| Agropecuario | 672.65 | 102 | 35 |
| Urbano | 55.02 | 99 | 0 |
| Otro | 20.3 | 0 | 1 |
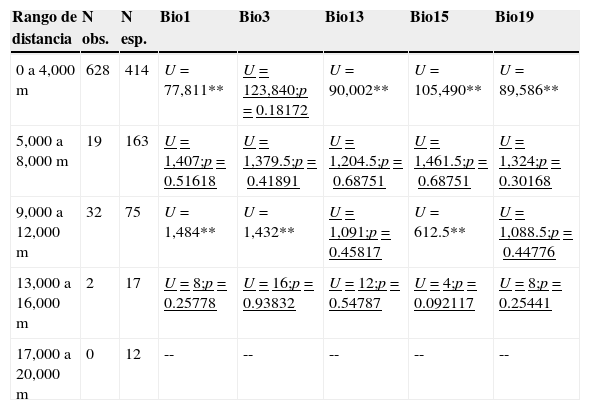
Las recolectas desarrolladas en el área presentaron diferencias significativas para todas las variables exploradas (temperatura media anual [Bio1]: U=117,900; p<0.01; isotermalidad [Bio3]: U=189,650; p<0.01; precipitación del mes más húmedo [Bio13]: U=134,330; p<0.01; estacionalidad de la precipitación [Bio15]: U=175,720; p<0.01; y la precipitación del trimestre más frío [Bio19]: U=146,550; p<0.01). Por otra parte, pese a que en el primer rango de distancia (de 0 a 4,999m) se observó el mayor número de registros (n=628), se presenta una diferencia significativa para todas las variables analizadas (tabla 5); es importante notar que para este rango el número de registros observados supera en 151% a los registros esperados aleatoriamente para este rango de distancia. Una situación similar se observa en el rango de 9,000 a 12,999m, en que, pese a contar con 32 registros botánicos, solamente la variable de isotermalidad no fue diferente entre ambos conjuntos analizados (U=1,091; p<0.4581) (tabla 5). Para los rangos comprendidos por las distancias entre 5,000 y 8,999 y el de 13,000 a 16,999m, la heterogeneidad ambiental es capturada por los registros previamente desarrollados, puesto que para todas las variables estudiadas no se presentaron diferencias significativas (tabla 5). Sin embargo, es importante señalar que los registros observados para estos 2 rangos apenas representan el 11% del esperado para estas distancias a carreteras. La clase de 17,000 a 20,000m no fue comparada, dado que no hubo registros observados recolectados para esta zona.
Valores de la prueba de Mann-Whitney para los distintos rangos de distancia a las carreteras y las variables bioclimáticas.
| Rango de distancia | N obs. | N esp. | Bio1 | Bio3 | Bio13 | Bio15 | Bio19 |
|---|---|---|---|---|---|---|---|
| 0 a 4,000 m | 628 | 414 | U=77,811** | U=123,840;p=0.18172 | U=90,002** | U=105,490** | U=89,586** |
| 5,000 a 8,000 m | 19 | 163 | U=1,407;p=0.51618 | U=1,379.5;p=0.41891 | U=1,204.5;p=0.68751 | U=1,461.5;p=0.68751 | U=1,324;p=0.30168 |
| 9,000 a 12,000 m | 32 | 75 | U=1,484** | U=1,432** | U=1,091;p=0.45817 | U=612.5** | U=1,088.5;p=0.44776 |
| 13,000 a 16,000 m | 2 | 17 | U=8;p=0.25778 | U=16;p=0.93832 | U=12;p=0.54787 | U=4;p=0.092117 | U=8;p=0.25441 |
| 17,000 a 20,000 m | 0 | 12 | -- | -- | -- | -- | -- |
** p < 0.01; valores subrayados p > 0.05.
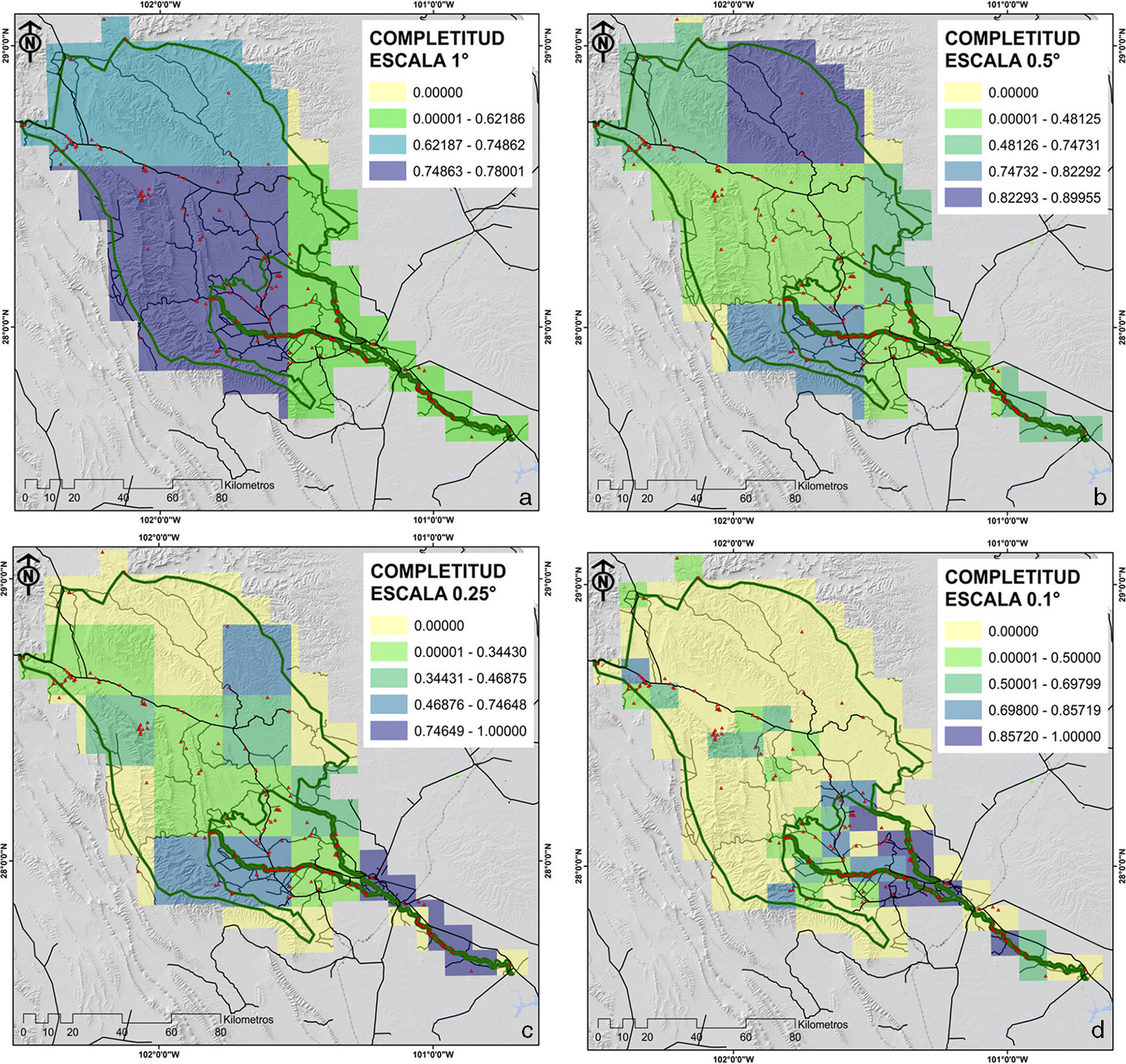
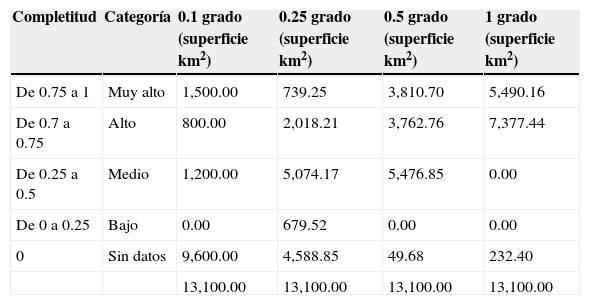
Para el caso de las resoluciones espaciales más bajas (1y 0.5°) la mayor proporción de la superficie de la reserva (98 y 99% del ANP, respectivamente) presentan valores de completitud que van de medios a muy altos (rangos de completitud de 0.25 a 1) (tabla 6; figs. 6a y 6b). En contraparte, para el caso de las resoluciones más finas de un cuarto de grado (0.25°) y de 0.10° (100 km2) se observó una disminución de la superficies para estos rangos altos y muy altos de completitud del orden del 65 y 26% del ANP, respectivamente (tabla 6; figs. 6c y 6d).
Rangos de completitud y superficie estimada (km2) para las distintas escalas de análisis.
| Completitud | Categoría | 0.1 grado (superficie km2) | 0.25 grado (superficie km2) | 0.5 grado (superficie km2) | 1 grado (superficie km2) |
|---|---|---|---|---|---|
| De 0.75 a 1 | Muy alto | 1,500.00 | 739.25 | 3,810.70 | 5,490.16 |
| De 0.7 a 0.75 | Alto | 800.00 | 2,018.21 | 3,762.76 | 7,377.44 |
| De 0.25 a 0.5 | Medio | 1,200.00 | 5,074.17 | 5,476.85 | 0.00 |
| De 0 a 0.25 | Bajo | 0.00 | 679.52 | 0.00 | 0.00 |
| 0 | Sin datos | 9,600.00 | 4,588.85 | 49.68 | 232.40 |
| 13,100.00 | 13,100.00 | 13,100.00 | 13,100.00 |
 1 grado; b) 0.5 grados; c) 0.25 grados, y d) 0.1 grados.")
Los taxa mejor representados en el inventario de plantas arbustivas fueron Vachellia rigidula, Vachellia farnesiana y Senegalia greggii, especies que son comunes en los matorrales del noreste de México y sur de Estados Unidos y, que en diversas localidades de la región ostentan altos valores de importancia en términos de la composición y estructura de estos hábitat (Canizales-Velázquez et al., 2009; Estrada-Castillón et al., 2005; García-Hernández y Jurado, 2008; Mora-Donjuán et al., 2013). Por otra parte, es importante señalar que acorde con la distribución geográfica de los registros, poco más del 60% de los cuadrantes de análisis que conformaron el área de estudio carece de datos botánicos (n=79 cuadrantes), y de los 52 cuadrantes que presentan información solamente 23 cuentan con un número mayor de 10 registros (fig. 1b). De igual forma, en el caso de la diversidad observada, el 90% de los cuadrantes contiene menos del 10% de la riqueza total registrada para el área (fig. 1c). Esto evidencia que estas zonas del área protegida no han sido sujetas a exploraciones botánicas, o bien la información no se encuentra disponible para su integración y análisis. Una planeación del esfuerzo de recolectas hacia estas áreas poco estudiadas derivaría, por un lado, en el registro de un mayor número de especies por unidad de estudio (cuadrantes de 100km2). De acuerdo con ese contexto, los sitios con un buen nivel del inventario podrían correlacionarse con variables bióticas y abióticas y facilitar la inferencia de patrones de riqueza a escala regional (Hernández-Pérez, González-Espinosa, Trejo y Bonfil, 2011); además, la información generada podría soportar el desarrollo de modelos predictivos que maximicen la información disponible sobre la diversidad botánica del área (Araujo y Guisan, 2006; Sánchez-Fernández, Lobo, Abellán y Millán, 2011).
El síndrome de los recolectores o de las carreteras (Llorente-Bousquets, Luna, Soberón y Bojórquez, 1994; Soberón et al., 2000) es uno de los sesgos descritos con más frecuencia en el desarrollo de las recolectas biológicas. En particular, este sesgo puede comprometer la representatividad de los muestreos y, por ende, no representar de forma adecuada la caracterización del nicho ecológico de las especies (Hortal et al., 2007; Hortal et al., 2008). Sin embargo, las causas de este fenómeno se deben en parte a la escasa promoción y apoyo para el desarrollo de muestreos planificados, lo cual ha propiciado que los investigadores aprovechen estas vías de acceso para documentar la biodiversidad de una región. De esta forma, y a pesar de este sesgo, se ha logrado integrar los inventarios y tener la base para estimaciones más precisas del número de especies de una región. Por otra parte, estos datos provenientes de colecciones botánicas desarrolladas para el área pueden soportar el análisis de patrones y procesos, considerando escalas de baja resolución espacial (Hijmans et al., 2000). En particular, en el caso del área de estudio, este sesgo es significativo (χ2=199.01; p<0.01), producto del 92% de los muestreos que se encuentran sobrerrepresentados a no más de 4,000m de las vías de acceso del área. En contraparte, las categorías de distancia mayores a 5,000m no cuentan con suficientes registros considerando un muestreo totalmente aleatorizado (tabla 2; fig. 2a). Esta condición puede evidenciar inventarios, en cuya composición taxonómica cobren mayor importancia las especies más tolerantes a condiciones ecológicas provenientes de disturbio, o describir aspectos marginales del propio nicho de las especies ubicadas dentro de estos rangos de distancias (Hijmans et al., 2000).
Otra tendencia observada en los muestreos y registros de plantas arbustivas documentadas para la zona ha sido el marcado esfuerzo hacia una limitada porción del rango altitudinal presente para la zona de estudio; de esa manera cerca del 90% del total de registros documentados se ubican en los rangos de 381 a 1,028 de altitud (m snm) (tabla 3; fig. 2c). Considerando que el gradiente altitudinal es un promotor de la variabilidad ecológica para diversos grupos botánicos (Encina-Domínguez y Villarreal-Quintanilla, 2002), entre ellos el conglomerado de plantas arbustivas, se puede señalar que, con base en este esfuerzo desarrollado, se corre el riesgo de representar parte del hábitat para ciertas especies con condiciones ecológicas particulares. Por último, respecto a los tipos de vegetación presentes para el área, se observó una desproporcionalidad de muestreo (χ2=518.13; p<0.01), enfatizándose hacia algunas comunidades vegetales como el matorral espinoso tamaulipeco, el cual comparativamente con un muestreo aleatorizado estratificado, se encuentra sobrerrepresentado en un 207% (tabla 4; fig. 2b). De igual forma, se observó una diferencia en el esfuerzo de muestreo efectuado hacia áreas de carácter no natural como zonas agrícolas y pecuarias, además de zonas urbanas, categorías que representan casi el 30% de los registros para el área. Para este caso, podemos señalar que la categorización del sitio de muestreo dentro de estas clases puede deberse a 3 condiciones probables: la primera, referente a la época de la recolecta cuando la localidad muestreada pudo pertenecer a otro tipo de vegetación (cambio de uso del suelo); la segunda, derivada de procesos de revegetación de uso de suelo (matorralización), proceso observado para otras regiones del noreste de México (Moreno-Talamantes y García-Aranda, 2012), y la tercera, posiblemente debida a la generalización relativa a la escala de la capa temática. Por otra parte, otros tipos de vegetación con una importante extensión dentro del área se encuentran subrepresentados, tal es el caso del matorral desértico rosetófilo, el chaparral, el bosque de encino y el matorral submontano (fig. 2b). En particular, esta última comunidad vegetal debe estudiarse de forma más enfática para cumplir, por lo menos, con la muestra requerida considerando un muestreo aleatorizado estratificado (n=135 registros), dado que es una comunidad con una alta diversidad específica, además de considerarse como una zona de transición entre áreas de carácter xerófilo y bosques de distintas condiciones (Canizales-Velázquez et al., 2009; Estrada-Castillón et al., 2004; Estrada-Castillón et al., 2005; Salinas-Rodríguez et al., 2013).
Cuando se considera la muestra total de registros, la heterogeneidad ambiental del área protegida se ve pobremente representada dentro del inventario botánico desarrollado, exhibiendo diferencias significativas para todas las variables entre los 2 conjuntos de datos analizados (observados contra aleatorios). Por otra parte, al analizar estas mismas variables considerando los distintos rangos de distancia a carreteras, se observa que para la primera comparación (rango de 0 a 4,999m), solamente la isotermalidad (Bio13) fue representada de forma adecuada (U=123,840; p<0.1817). En contraparte, las variables restantes fueron significativamente diferentes (tabla 5). Es importante señalar que para este rango de distancia se determinó la mayor proporción de registros observados; sin embargo, el área proporcional comprendida por este rango ocupa el 40% de la superficie del área natural protegida (tabla 2), lo que le confiere la posibilidad de una mayor variabilidad ambiental para esta zona específica. Aunado a ello, y con un análisis más detallado de la distribución de los registros, puede señalarse que estos se encuentran agrupados hacia distancias entre los 1,000 y 2,000m de las carreteras.
Para los subsecuentes rangos de distancia analizados, se logra representar entre 2 (rango de distancia a carreteras de 9,000 a 12,999m) y 5 variables (rangos de 5,000 a 8,999 y 13,000 a 16,999m) (tabla 5). Para el caso del rango de 9,000 a 12,999m, se determinaron similitudes entre ambas comparaciones para las variables de precipitación (Bio13: precipitación del mes más húmedo, y Bio19: precipitación del trimestre más frío), mientras que para los restantes rangos no hubo diferencias para la totalidad de las variables. Sin embargo, un proceso que pudo haber influido en la caracterización de esta heterogeneidad ambiental para estos rangos de distancias a carreteras es la reducción de la superficie dentro de estas categorías de distancia dentro de la reserva, situación que disminuye la variación ambiental y, por ende, facilita la caracterización de las variables analizadas, aunque se cuente con una muestra de registros menor a lo esperado (tabla 5). En este contexto, es importante enfatizar que el cubrimiento de la infraestructura de caminos dentro del área estudiada puede contribuir para una buena caracterización de la misma, si al menos se considera un diseño de muestreo para representar los aspectos geográficos y ambientales de la región en cuestión, con lo que se minimizan las limitaciones prácticas como la accesibilidad a los sitios, la falta de tiempo, la disponibilidad de fondos, así como a los objetivos de la propia investigación (Araujo y Guisan, 2006; Hijmans et al., 2000; Hortal y Lobo, 2006; Mateo, Felicísimo y Muñoz, 2011; Villaseñor et al., 2005).
En general podemos inferir que el conocimiento del grupo botánico analizado es adecuado para las escalas pequeñas (1 y 0.5°), dado que estas resoluciones espaciales nos describen valores altos de completitud para la mayor proporción del área de estudio (más del 95%) (tabla 6; figs. 6b y 6b). A niveles geográficos más finos o escalas más grandes la completitud disminuye, debido a una mayor proporción de vacíos de información (áreas sin datos de recolectas) (tabla 6; figs. 6c y 6d), situación que infiere la necesidad de realizar esfuerzos adicionales que permitan documentar mejor la riqueza florística de este grupo (Villaseñor et al., 2005). Por otra parte, es importante señalar que para las resoluciones más finas, la información sobre los registros de las especies termina fragmentándose, y en algunos casos reduciéndose a uno o pocos registros y especies, lo que limita el funcionamiento del estimador en términos de su capacidad predictiva, dado que la muestra no reúne los supuestos para su estimación, como lo ocurrido en nuestro caso al no contar con la cantidad mínima de réplicas para su análisis (Jiménez-Valverde y Hortal, 2001; Villaseñor et al., 2005). Esta situación se evidencia para 9 cuadrantes (12, 24, 52, 64, 83, 89, 94, 97, 99) en el nivel de partición más grande (0.1×0.1°). De forma paradójica, el cuadrante 19 que presenta la mayor intensidad de muestreo y, por ende, el mayor número de especies (36 especies y 97 registros), se describe con un valor bajo de completitud (0.312), lo que evidencia, en primer lugar, la escasa proporción del área muestreada dentro del propio cuadrante de 100km2 y la carencia de un diseño para tratar de representar la heterogeneidad de la zona de estudio. Esta “fotografía” puede deberse en parte, a la proporción de especies raras (aquellas que se presenta 1 o 2 veces en el cuadrante) dentro del propio inventario (Jiménez-Valverde y Hortal, 2001; López-Gómez y Williams-Linera, 2006; Moreno, 2001).
El presente estudio se realizó con un grupo vegetal conspicuo, bien conocido y de distribución generalizada en el noreste de México, y a través de estos datos se describieron ciertos procesos relativos a la sistematización de este conocimiento, lo que induce a pensar sobre lo que sucede con otros grupos menos conocidos. Si bien es cierto que los datos de recolectas en el APRN CADDR 004 contribuyen a describir la importancia de esta zona para la biodiversidad, esta información no está exenta de presentar sesgos de distancia a carreteras, por rangos altitudinales y por tipos de vegetación, los cuales pueden a llegar a comprometer la utilidad de la información, puesto que evidencia la falta de un cubrimiento geográfico y ambiental del área en estudio. Aun así, puede señalarse que los datos recolectados para el área pueden ser útiles cuando se busquen respuestas a escalas pequeñas.
Dado que el área en cuestión corresponde a un área natural protegida de carácter federal, es conveniente dirigir inversiones hacia la sistematización del conocimiento taxonómico de la zona, basadas en diseños de muestreo sistemático, para que los datos derivados de dichos trabajos puedan utilizarse en la descripción de patrones, aspectos ecológicos y biogeográficos del área. Por otra parte, un esfuerzo planificado en la toma de datos sobre las especies de plantas arbustivas podría ser útil en combinación con herramientas y nuevos enfoques informáticos como los modelos de nicho ecológico y el desarrollo de análisis espaciales a través de sistemas de información geográfica. Ello podría traducirse en una más y mejor información que pueda soportar la toma de decisiones para la conservación de esta región. Por último, es crucial enfatizar la importancia del trabajo taxonómico y el desarrollo de recolectas biológicas como el pilar del conocimiento sobre nuestra biodiversidad y la base para el planteamiento de estrategias de conservación y aprovechamiento sustentable de las áreas protegidas y el APRN CADDR 004.
El primer autor agradece al Conacyt (6153) por la beca para estudio de Posgrado otorgada.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.