







El objetivo de este trabajo fue conocer y describir el ciclo reproductivo y el dimorfismo sexual morfométrico de Homonota fasciata en el monte de San Juan. Para analizar la reproducción, se midió en hembras la longitud y ancho de folículos yemados y/o huevos en oviducto; en machos se midió el ancho y largo de testículos, con el fin de estimar sus volúmenes. El dimorfismo sexual se examinó midiendo 10 variables morfométricas en 49 individuos (26 hembras y 23 machos). Homonota fasciata inició su actividad reproductiva en primavera para los 2 años de registro. El tamaño de camada fue de un huevo. En hembras, se evidenció al menos una triple postura y el ciclo gonadal varió con las estaciones. En machos, el desarrollo testicular no exhibió variaciones, demostrando una condición reproductiva continua a través de la estación de actividad. Se halló dimorfismo sexual en 2 variables: longitud radio-cúbito, siendo mayor en hembras que en machos; y longitud tibia-fíbula, siendo esta variable mayor en machos y atribuyendo el dimorfismo encontrado a la marcada territorialidad de la especie. Este estudio es un valioso aporte al conocimiento de la biología de la especie, en un contexto tanto ecológico como de conservación.
The aim of this study was to know and describe the reproductive cycle and sexual dimorphism of Homonota fasciata form the Mount of San Juan. To analyze reproduction, females were measured in the length and breadth of yemados follicles and/or eggs in the oviduct, in males testes length and width were measured, in order to estimate their volumes. Sexual dimorphism was examined measuring 10 morphometric variables in 49 individuals (26 females and 23 males). Homonota fasciata started their reproductive activity in spring for 2 years recorded. The smallest size was 1 egg. Females, showed at least a triple posture and the gonadal cycle varied with seasons. In males, testicular development did not exhibit variations, showing a continuous reproductive condition through the activity season. We found sexual dimorphism in 2 variables: radio-ulna length, being higher in females than in males, and tibia-fibula length, being this variable major in males and attributing dimorphism found to the marked the territoriality of the species. This study is a valuable contribution to the knowledge of this species in a context of ecology and conservation.
El conocimiento de parámetros poblacionales tales como el ciclo reproductivo, los hábitos alimenticios y las estrategias en el uso del hábitat permiten evaluar objetivamente la vulnerabilidad de las especies y su estado de conservación. Resulta imprescindible tener en cuenta dicha información al momento de tomar decisiones de manejo de las especies y sus hábitats, frente al nuevo escenario de cambios ambientales que sufre el planeta (Christie, 1984; Aun y Martori, 1998; Martori et al., 2002).
Las lagartijas, por su amplia distribución en el mundo y la gran diversidad de ambientes que habitan, presentan diferentes patrones reproductores, estableciéndose 3 tipos generales: uno de tipo continuo, que es observado comúnmente en especies que habitan zonas tropicales; otro de tipo anual, presente en lagartos tanto de zonas tropicales como templadas, y por último, el tipo bianual o trianual de zonas desérticas de alta montaña (Hernández-Gallegos et al., 2010).
Hay pocos antecedentes de trabajos que abordan esta temática en el género Homonota, como los de Aun y Martori (1994), Cruz (1994), Blanco et al. (2001), Ibargüengoytía y Casalins (2007) y Blanco et al. (2011).
Muchos reptiles son sexualmente dimórficos en el tamaño del cuerpo, coloración o en su comportamiento como resultado de presiones selectivas que operan diferencialmente entre machos y hembras (Villavicencio et al., 2003), pudiendo relacionar las causas del dimorfismo con funciones ecológicas, estrategias de alimentación o selección sexual, debido a que diferentes tamaños o aspectos corporales son más efectivos que otros (Pianka, 1986; Smith et al., 1997; Herrel et al., 2004; Laspiur et al., 2006).
El género Homonota perteneciente a la familia Phyllodactylidae en Argentina se distribuye desde la frontera con Bolivia (25° S) hasta la provincia de Santa Cruz (52° S), constituyendo los geckos de distribución más austral del mundo.
Homonota fasciata es un lagarto pequeño, que mide hasta 60mm de longitud hocico cloaca, con cola poco más larga que el cuerpo y muy autotómica. Posee ojos grandes sin párpado móvil, con pupila vertical. Las escamas que cubren la cabeza son granulares y yuxtapuestas. Las escamas dorsales son grandes, con carenas elevadas que le dan una textura granulosa al cuerpo (Cabrera, 2009). Homonota fasciata posee hábitos crepusculares, es insectívoro y es común en roquedales serranos y grietas de construcciones. Se distribuye en ambientes de monte y Chaco hasta los 2 500m snm (Cei, 1993).
Homonota fasciata posee 2 especies sinónimas, H. horrida en Argentina y H. pasteuri en Brasil. En este trabajo se siguió este criterio de sinonimia propuesto por Abdala y Lavilla (1994), considerando a las 3 como una misma especie.
El hábitat de la especie se encuentra sometido a diversas presiones, por un lado antrópicas, debido al avance de las fronteras agrarias y por otro lado, a presiones ambientales, dadas las características de extrema aridez del sitio. Por esta razón, el objetivo de este trabajo fue conocer y describir el ciclo reproductivo y el dimorfismo sexual morfométrico de H. fasciata en un sector del monte de San Juan, con el propósito de aportar datos relevantes de su biología, que permitan fijar pautas de manejo para la misma, así como para los ambientes que habita.
Materiales y métodosÁrea de estudio. Los médanos Grandes pertenecen a la provincia fitogeográfica del monte y comprenden una amplia región al sudeste de la provincia de San Juan, cubriendo un área de 2 000km2 en el pie de monte oriental de la sierra de Pie de Palo, a 576m snm (31°44' S, 68°10' O). La zona es una llanura eólica-aluvial cuaternaria, surcada por desagües temporarios y cadenas de médanos. El clima de la zona es seco y cálido, con una temperatura media anual de 18° C y precipitaciones estivales medias de 103mm (De Fina, 1992). Es una zona de médanos vivos donde predomina vegetación xerófila formando parches de densa vegetación dentro de una matriz de suelo desnudo denominada interparche (Aguiar y Sala, 1999).
Trabajo de campo. La recolección de ejemplares se llevó a cabo mediante 100 trampas de caída tipo Barber (Tellería, 1986), durante 2 años consecutivos (1999-2000), las trampas permanecieron abiertas en los periodos de actividad de la especie y se revisaron 1 vez por semana. Se recolectaron muestras mínimas representativas luego de analizar las varianzas en los parámetros biológicos en premuestreos. Los ejemplares capturados fueron sacrificados mediante frío, fijados por inyección de formaldehído al 10% y conservados en alcohol al 70%. Las muestras se depositaron en la Colección Científica Herpetológica de la Universidad Nacional de San Juan.
Reproducción. Se realizaron observaciones en individuos adultos, a los que se les midió la longitud hocico-cloaca (LHC) y se disecaron para la extracción de gónadas y cuerpos grasos abdominales. En hembras se registró el número de folículos yemados y el número de huevos en oviducto; midiendo el largo y el ancho de los mismos con un calibre tipo Vernier de precisión 0.01mm. Para el cálculo de sus volúmenes, se aplicó la fórmula propuesta por Dunham (1983) que es utilizada para el cálculo del volumen de una esfera elipsoide. También se evaluó la condición del oviducto, ensanchado o no.
En cuanto a los machos, se observó el estado convoluto de los epidídimos como indicador de madurez sexual y se midió el ancho y largo de testículos, para luego calcular sus volúmenes utilizando la fórmula de Dunham (1983).
Los cuerpos grasos extraídos se pesaron en una balanza analítica, con una precisión de 0.0001g.
Para realizar comparaciones entre muestras y entre sexos se utilizaron análisis de varianza y covarianza (Andeva y Ancova), respectivamente. Las variables reproductivas fueron transformadas a su logaritmo natural para lograr una tendencia lineal. Se analizaron los efectos de las estaciones: primavera, verano, otoño, sobre los ciclos de cuerpos grasos mediante Ancova y sobre los ciclos gonadales a través de Andeva.
Para analizar las variaciones estacionales de los parámetros reproductivos, se tuvieron en cuenta las estaciones térmicas propuestas por Poblete y Ruiz (2006), las cuales contemplan las siguientes fechas de inicio y finalización de las estaciones: verano, 14 de noviembre al 10 de marzo; otoño, 11 de marzo al 25 de mayo; invierno, 26 de mayo al 31 de agosto; primavera, 01 de septiembre al 13 de noviembre.
Dimorfismo sexual. Se midieron 10 variables morfométricas en hembras y machos adultos, utilizando un calibre tipo Vernier de precisión 0.01mm: longitud hocico-cloaca (LHC); longitud de cabeza (LCA); ancho de cabeza, entre las comisuras de la boca (ANCA); altura máxima de la cabeza (ALCA); distancia de separación entre los miembros anteriores y posteriores (DEM); longitud de húmero (LH); longitud radio-cúbito (LRC); longitud de fémur, desde la ingle hasta la rodilla (LF); longitud tibia-fíbula (LTF) y largo de la cola (LC).
Con el propósito de probar las diferencias morfológicas sexuales en cuanto a la longitud hocico-cloaca (LCH) se realizó un Andeva. Para examinar la relación entre cada variable y la longitud hocico-cloaca (LHC), se llevaron a cabo análisis de regresión lineal. En los casos en que las regresiones fueron significativas, se analizó el dimorfismo con un Ancova, tomando como covariable la longitud hocico-cloaca (LHC), a fin de remover el efecto del tamaño corporal sobre el resto de las variables morfométricas.
ResultadosSe analizó un total de 49 ejemplares, 26 hembras y 23 machos, recolectados en primavera, verano y otoño de ambos años, de acuerdo con el periodo de actividad de la especie. La proporción de sexos fue variable en las diferentes estaciones, hallando en las trampas todos individuos del mismo sexo en algunas estaciones.
Reproducción. Homonota fasciata presenta un ciclo reproductivo anual, iniciando su actividad reproductiva en primavera temprana para los 2 años de registro. El tamaño mínimo reproductivo fue de 40mm. Se observó que el tamaño de camada fue siempre de un solo huevo. Los menores tamaños de ejemplares juveniles se registraron para los meses de noviembre, diciembre y enero, es decir, para los meses correspondientes a las estaciones primavera y verano.
En hembras, se observó la presencia simultánea de huevo en oviducto, folículos yemados y oviducto ensanchado (Fig. 1).

En cuanto a la actividad gonadal de hembras, se encontró una variación significativa a través del tiempo (Andeva, F(3; 245)=11.33; p<0.001); N=26) (Fig. 2). Mostrando la máxima actividad en primavera, disminuyendo gradualmente hasta el otoño, estación en la cual todas las hembras se encontraron en estado no reproductivo. Para los machos, el desarrollo testicular fue sincrónico con el ciclo gonadal de hembras, pero no se observaron variaciones significativas entre las estaciones (Andeva, F(3; 3.09)=1.68; p=0,20); N=23) (Fig. 3).


El ciclo de cuerpos grasos de machos no mostró diferencias significativas con las estaciones (Ancova, F(3; 6.56)= 1.59; p=0.22, covariable: LHC) (Fig. 4); en cambio en hembras, la reserva de lípidos varió significativamente (Ancova, F(3; 13.97)=4.66; p=0.01, covariable: LHC) (Fig. 5). Esta variación acompaña la de los ciclos gonadales, es decir, se hallan los mayores pesos de cuerpos grasos en primavera, luego disminuyen hacia el verano y, finalmente, inicia un aumento en su masa cuando comienza el otoño. Dimorfismo sexual. No se encontraron diferencias significativas en las longitudes hocico-cloaca entre machos y hembras (Andeva; F(1; 1)=0.02; p=0.87).


Con respecto a las demás variables morfométricas, a excepción del largo de cola (LC), todas las variables correlacionaron positivamente con el largo hocico-cloaca (LHC).
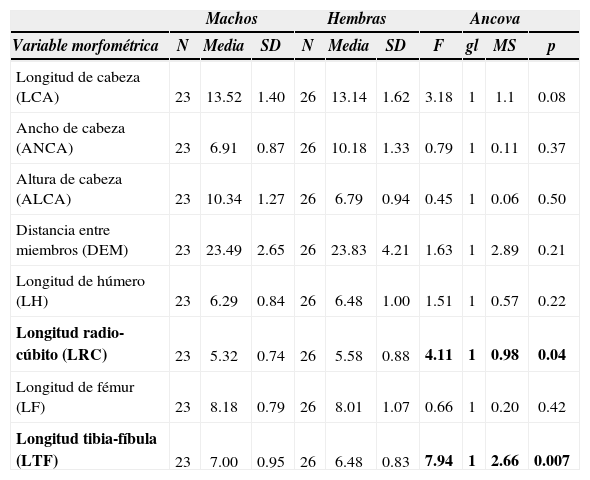
Homonota fasciata presentó dimorfismo sexual sólo en 2 variables. Por un lado la longitud radio-cúbito fue sexualmente dimórfica (Cuadro 1), siendo esta variable mayor en hembras que en machos; y por otro lado, se encontró dimorfismo en la longitud tibia-fíbula (Cuadro 1), hallándose mayores longitudes en ejemplares machos. No se encontraron diferencias significativas entre machos y hembras para el resto de las variables morfométricas.
Estadística descriptiva y análisis de covarianza para las variables morfométricas en comparación. En negritas, las diferencias significativas entre sexos; n=49
| Machos | Hembras | Ancova | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variable morfométrica | N | Media | SD | N | Media | SD | F | gl | MS | p |
| Longitud de cabeza (LCA) | 23 | 13.52 | 1.40 | 26 | 13.14 | 1.62 | 3.18 | 1 | 1.1 | 0.08 |
| Ancho de cabeza (ANCA) | 23 | 6.91 | 0.87 | 26 | 10.18 | 1.33 | 0.79 | 1 | 0.11 | 0.37 |
| Altura de cabeza (ALCA) | 23 | 10.34 | 1.27 | 26 | 6.79 | 0.94 | 0.45 | 1 | 0.06 | 0.50 |
| Distancia entre miembros (DEM) | 23 | 23.49 | 2.65 | 26 | 23.83 | 4.21 | 1.63 | 1 | 2.89 | 0.21 |
| Longitud de húmero (LH) | 23 | 6.29 | 0.84 | 26 | 6.48 | 1.00 | 1.51 | 1 | 0.57 | 0.22 |
| Longitud radio-cúbito (LRC) | 23 | 5.32 | 0.74 | 26 | 5.58 | 0.88 | 4.11 | 1 | 0.98 | 0.04 |
| Longitud de fémur (LF) | 23 | 8.18 | 0.79 | 26 | 8.01 | 1.07 | 0.66 | 1 | 0.20 | 0.42 |
| Longitud tibia-fíbula (LTF) | 23 | 7.00 | 0.95 | 26 | 6.48 | 0.83 | 7.94 | 1 | 2.66 | 0.007 |
Reproducción. Los patrones observados en la actividad reproductiva de H. fasciata, en cuanto al inicio del ciclo en primavera temprana y al tamaño mínimo reproductivo de machos y hembras, coinciden con los resultados hallados para esta misma especie en la provincia de Salta porCruz (1994) y en Córdoba por Aun y Martori (1994), en Catamarca para H. andicola (Blanco et al., 2009) y en San Juan para H. underwoodi (Blanco et al., 2011). Por el contrario, los tiempos de actividad reproductiva que presentó esta especie difieren con los observados por Ibargüengoytia y Casalins (2007) para H. darwini en la Patagonia, cuya distribución está restringida a zonas frías con reducida disponibilidad temporal diaria y estacional, lo que podría explicar las diferencias.
El tamaño de puesta registrado es fijo, de un huevo para todas las hembras, lo que concuerda con otros estudios como los de Gómez y Acosta (1998), Martori et al. (2002) y Blanco et al. (2009) para otras especies congéneres, indicando que ésta es una característica que se conserva filogenéticamente.
El hecho de haber encontrado hembras de H. fasciata en proceso de vitelogénesis, conteniendo un huevo oviductal y con el otro oviducto ensanchado simultáneamente, sugiere que esta especie realiza al menos 3 posturas por ciclo. Evidencias de múltiples posturas han sido halladas en otros trabajos afines (Vitt, 1986, 2000; Kretzschmar y Abdala, 2001; Cacciali et al., 2007; Blanco et al., 2009) y pueden estar relacionadas a estrategias para garantizar la supervivencia de al menos un huevo, en ambientes con severos riesgos de deshidratación (Cruz, 1994). Los menores tamaños de juveniles registrados a fines de primavera y luego en verano, indican que los nacimientos se producen en estas épocas, ratificando que hay más de una camada por ciclo reproductivo.
El ciclo de cuerpos grasos en hembras presentó una variación significativa a través de las estaciones, acompañando inversamente al ciclo reproductivo. Esto está asociado a la utilización de energía almacenada, por parte de hembras, para emerger de la hibernación y dar inicio a la vitelogénesis en la época en la que aún no hay suficiente cantidad de alimento disponible, concordando con Blanco et al. (2011).
El hecho de que en machos la reserva de lípidos no varíe a través de las estaciones y que no se observen diferencias significativas en el desarrollo testicular entre las estaciones, puede ser indicio de actividad reproductiva continua (Cruz, 1994), pero no sincrónica entre todos los machos, coexistiendo en la población individuos reproductivos y no reproductivos. Resultados que coinciden con lo hallado por Ibargüengoytia y Casalins (2007) para H. darwini, pero difieren con lo observado para H. whitti por Martori et al. (2002) y para H. underwoodi por Blanco et al. (2011). Además, el ciclo de los machos corresponde con la actividad reproductiva de las hembras de la especie y explica a su vez los indicios de varias posturas por ciclo.
La filogenia y la historia biogeográfica juegan un rol importante en la determinación de los patrones reproductivos en los Squamata (Dunham y Miles, 1985), por lo tanto, podría inferirse una componente filogenética en la determinación del ciclo reproductivo, como también cierta plasticidad de los eventos en función de las condiciones ambientales. En ambientes tropicales la reproducción de los geckónidos es continua (Vitt, 1986) y si estos ambientes muestran cierta estacionalidad, el esfuerzo reproductivo también lo refleja (Aun y Martori, 1994).
Dimorfismo sexual. De acuerdo con los resultados obtenidos, el dimorfismo sexual hallado en H. fasciata en los miembros anteriores y posteriores no habría surgido por selección sexual, sino que podría deberse a patrones diferenciales en las trayectorias de crecimiento después de la madurez sexual de machos y hembras (Cox y John-Alder, 2007).
En general, los miembros más largos en los machos se asocian con encuentros agonísticos intra e interespecíficos (Hibbitts et al., 2005; Laspiur y Acosta, 2007) y, además, se conoce que esta especie es marcadamente territorial (Abdala, 1986).
Abdala (1986) observó que el macho de H. fasciata adopta una posición denominada “dressée”, en la cual el individuo estira completamente sus miembros anteriores y posteriores, curvando su dorso, como postura intimidatoria ante la presencia de otro individuo de la misma especie u otra; comportamiento que resalta la territorialidad de la especie y que se podría asociar al dimorfismo encontrado.
Nuestros resultados coinciden con los de Cruz (1994), ya que para la misma especie no se observaron diferencias en el LHC entre los sexos. Algunos geckos de la familia, en Nueva Zelanda (Hare y Cree, 2005) y en Sudáfrica (Hibbitts et al., 2005), tampoco presentan dimorfismo sexual en el LHC.
Por otro lado, estos resultados difieren con los de Ibargüengoytia y Casalins (2007) para H. darwini de Patagonia y con los hallados por Vitt (1986) puesto que en geckónidos de Brasil se encontraron hembras de mayor tamaño, con múltiples posturas por estación reproductiva, pudiendo haber obrado en ese sentido la selección sexual.
El dimorfismo hallado por Cruz (1994) en el LCA, ALCA y ANCA entre sexos de H. fasciata, no coincide con nuestros resultados a pesar de tratarse de la misma especie en estudio, aunque de ambientes muy distintos y diferente latitud. Esto sugeriría que el dimorfismo sexual tal vez no esté fijado fuertemente por la filogenia, sino que dependería evolutivamente de la historia reciente de cada especie y su distribución actual.
Sería necesario conocer más aspectos sobre la biología de esta especie y realizar estudios de ontogenia para poder interpretar el dimorfismo hallado y el existente en otras especies del género en ambientes de monte, para poder realizar comparaciones e interpretaciones en un contexto evolutivo.
A la Dirección de Conservación, Subsecretaría de Conservación y Áreas Protegidas dependiente de la Secretearía de Ambiente y Desarrollo Sustentable de la provincia de San Juan por los permisos de colecta otorgados (No 58-2000-SA). A Pablo Gómez, Juan Manuel Acosta, Rodrigo Acosta y Sol Acosta por la ayuda en el campo; a Ricardo Martori por el apoyo brindado con los proyectos relacionados con este estudio. Este trabajo fue financiado parcialmente por el proyecto CICITCA21-E 333 (UNSJ) (Director Juan Carlos Acosta). Finalmente, agradecemos los comentarios de 2 revisores anónimos, que ayudaron a mejorar el contenido del manuscrito.