





El conocimiento de los parámetros tróficos ayuda a comprender la historia natural de la especie, sus requerimientos ecológicos y estrategias involucradas en la búsqueda de alimento. Por ello, describimos y analizamos la dieta de Phymaturus cf. palluma y su variación en términos estacionales e intrapoblacionales. Los ítems fueron identificados en términos cuali-cuantitativos. Se estimó el IRI según el volumen, numerosidad y frecuencia de ocurrencia de los ítems. Se calcularon porcentajes de material vegetal y parásitos nemátodos en tracto digestivo. Phymaturus cf. palluma presenta una alimentación herbívora basada en estructuras vegetales reproductivas y vegetativas. El ítem “flores” fue fundamental, tanto en adultos como juveniles. En adultos macho y hembra, las flores de Adesmia pinnifolia fueron ítems fundamentales, sin embargo, la similitud cualitativa entre machos y hembras fue baja. Los machos presentaron mayor amplitud trófica respecto a hembras. Las estaciones de verano fueron las de mayor diversidad trófica. En cuanto al material vegetativo, hubo diferencias significativas sólo en el consumo de hojas entre estaciones. La competencia estacional por el recurso alimentario sería muy leve debido al amplio espectro de ítems encontrados en las distintas fechas. Phymaturus cf. palluma es un lagarto generalista que forrajea material vegetal en forma activa.
Knowledge of the trophic parameters helps to understand the natural history of the species, their ecological requirements and strategies involved in the food acquisition. Therefore, we describe and analyze the diet of Phymaturus cf. palluma regarding to seasonal and intra populational variation. Items were identified according qualitative and quantitative terms. IRI index was estimated as function of volume, numerosity and frequency of items. Percentages of vegetal material and parasitic nematodes in the digestive tract were calculated. Phymaturus cf. palluma present an herbivorous diet based upon vegetative and reproductive plant structures. In adults and juvenile individuals “flowers” were fundamental item. Overall, in male and female adults, flowers of Adesmia pinnifolia were fundamental; however the qualitative similarity among them was low. Trophic amplitude was higher in males than in females. Summers were the more trophic diversity seasons. Significant differences in the “leaves” consumption between seasons were founded. Seasonal competence for food should be low due to the wide spectrum of items in different periods. Phymaturus cf. palluma is a generalist lizard that feeds vegetal material actively.
El estudio del nicho trófico permite conocer aspectos relacionados con el espectro nutricional en el contexto de la población, incluyendo la comparación de las preferencias de los subgrupos sexuales y etarios de la especie, como también las variaciones estacionales. Además, permite conocer características comportamentales en la obtención de los recursos (Aun et al., 1999; Teixeira-Fialho et al., 2003). La descripción y cuantificación de la dieta, son uno de los principales aspectos en los estudios de ecología de las especies (Pianka, 1973; Aun et al., 1999; Sih y Christensen, 2001; Vitt y Pianka, 2007). La dieta está estrechamente relacionada con la abundancia del recurso alimenticio, la competencia intraespecífica, la capacidad de captura por parte de los individuos y las condiciones del hábitat (Aun et al., 1999).
La mayoría de los lagartos son exclusiva o principalmente carnívoros (Cooper y Vitt, 2002), insectívoros y a menudo oportunistas (Pianka, 1973). Sin embargo, se ha observado en algunas especies cierta tendencia a la herbivoría en lagartos pequeños y de climas fríos (Espinoza et al., 2004). A pesar de estas diferencias, herbívoros y carnívoros enfrentan problemas dispares con respecto a la dieta y digestión. La materia animal es fácilmente digerida y asimilada, mientras que la mayoría de la energía en el material vegetal está contenida en compuestos fibrosos, como la celulosa, para la cual los vertebrados carecen de enzimas para su digestión. Por ello, los herbívoros poseen especializaciones intestinales —ausentes en carnívoros— como un ciego, valvas intestinales y presencia de nemátodos en el intestino posterior, que facilitan la fermentación de componentes fibrosos y alargan el tiempo de retención de digestión, aumentando así, la eficiencia digestiva (Stevens, 1980; Iverson, 1982; Stevens y Hume, 1998).
Phymaturus es un género de lagartos herbívoros y vivíparos que se distribuye a partir del noroeste de Argentina, desde la provincia de Catamarca y a lo largo de los cordones montañosos de los Andes hasta la estepa patagónica y en áreas adyacentes a la cordillera de los Andes en Chile (Díaz-Gómez, 2008, Lobo et al., 2012). Phymaturus cf. palluma es un lagarto de aspecto robusto y deprimido dorso-ventralmente, de tamaño hasta 110mm; de hábitos saxícolas, ocupa principalmente afloramientos rocosos y se refugia en las grietas o diaclasas de los roquedales. Dadas las particularidades del género, las especies de Phymaturus recientemente han sido categorizadas como “vulnerables” (Abdala et al., 2012).
Por otra parte, son escasos los trabajos publicados sobre las características tróficas de especies del género Phymaturus en el oeste árido de Argentina (Videla, 1983; Acosta et al., 2008). Debido a que no existen estudios previos sobre ecología trófica en P. cf. palluma en San Juan, esta investigación resulta importante por describir y analizar la dieta en términos intrapoblacionales, sexos y grupos de edad, y estacionales. El propósito de este estudio es aportar información sustancial de la especie que permita conocer su rol en ecosistemas andinos, así como también, proporcionar conocimientos teóricos frente a la toma de decisiones de manejo y conservación.
Materiales y métodosÁrea de estudio. Los muestreos se realizaron durante 2 periodos en las siguientes estaciones: primavera-verano 2009 y otoño 2010 (periodo 1); primavera-verano 2010 y otoño 2011 (periodo 2), en la Quebrada de la Puerta (32°31.31' S; 69°42.02' O; 2 878m snm), departamento Calingasta, provincia de San Juan, Argentina. Fitogeográficamente corresponde a la provincia Altoandina (Cabrera y Willink, 1973). Este sector está incluido en la cordillera de los Andes, en la margen oeste del valle de Iglesia-Calingasta-Uspallata, con una orientación norte-sur.
Captura de los ejemplares y análisis de los contenidos gastrointestinales. Las muestras fueron realizadas en el marco de un proyecto mayor destinado a conocer la reproducción, hormonas, esqueletrocronología y estudios sistemáticos. Las capturas se realizaron mediante el método lazo corredizo y a mano. Posteriormente, los ejemplares fueron sacrificados mediante una inyección intraperitoneal de tiopental sódico y se fijaron en formaldehído al 10%. Las muestras se depositaron en la Colección Científica Herpetológica de la Universidad Nacional de San Juan (UNSJ).
En el laboratorio se procedió a la disección de los tractos gastrointestinales. Para la identificación de los ítems alimentarios se utilizó una lupa binocular estereoscópica (10–40 x). Los ítems vegetales se identificaron mediante diagnóstico con herbario, previamente recolectado en el área de estudio y determinación taxonómica propuesta por Kiesling (1994; 2003). Se registró la numerosidad de flores, frutos y semillas —estructuras vegetales reproductivas— y se estimó el volumen de cada uno de ellos utilizando la fórmula de una esfera elipsoide de Dunham (1983): V= 4/3 (½L) (½W)2, donde L= largo de la presa y A= ancho máximo de la presa. Para las estructuras no reproductivas (hojas y tallos), como también para parásitos nemátodos, se estimó el porcentaje de presencia en relación con el contenido total del tracto digestivo de cada individuo.
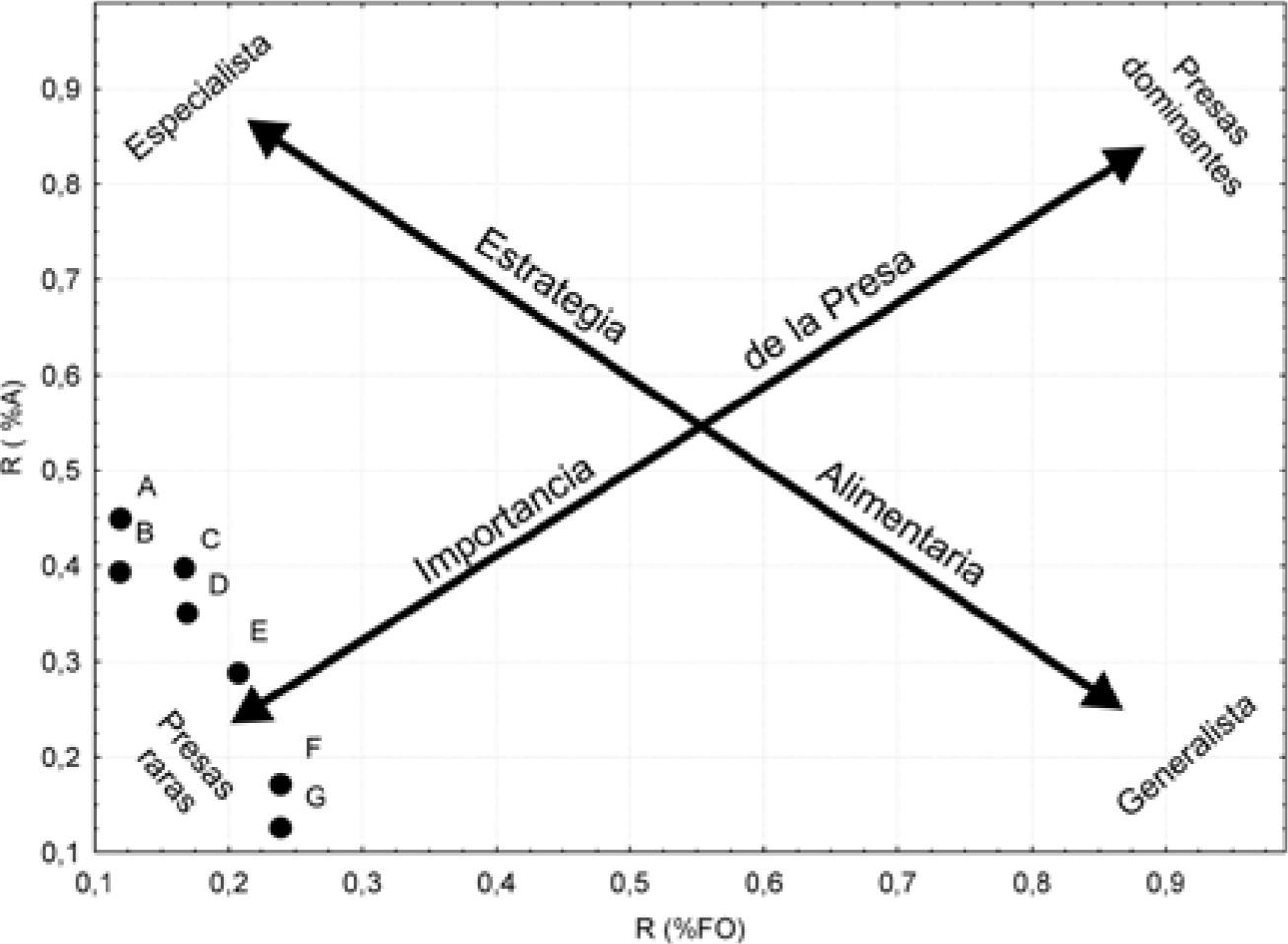
Análisis de datos. Para cada categoría alimentaria: flores, frutos y semillas, se calculó el índice de importancia relativa (IRI) (Pinkas et al., 1971) a fin de determinar la contribución de cada ítem en la dieta. IRI= %FO (%N+%V); donde %N expresa la importancia numérica, %V la importancia volumétrica y %FO expresa la proporción de estómagos conteniendo una categoría específica. Para estimar la variación intrapoblacional y estacional del material vegetal reproductivo: flores, frutos y semillas, se calculó el IRI entre sexos, grupos de edad y estaciones. Para las comparaciones en términos temporales del material vegetativo no reproductivo, hojas y tallos, los valores de porcentaje fueron transformados en categorías ordinales según sus valores representados en las muestras (de 1 a 4) y estas categorías fueron analizadas con la pruebas no paramétricas de Kruskal-Wallis. Para los análisis estadísticos se utilizaron los programas Statistica versión 7.0 (StatSoft, 2004) y Multivariate statistical package (Mvsp) (Kovach, 2001). Para determinaciones de amplitud de nicho trófico y dominancia de ítems alimentarios en particular, se utilizaron los índices de Shannon-Wiener e índice de Simpson, respectivamente. Con el propósito de detectar diferencias cualitativas de la alimentación entre sexos y estaciones, se calculó el índice de similitud de Jaccard. Además, se utilizó el índice de similitud de Morisita-Horn para las comparaciones cuantitativas entre las variables %N, %FO, %Vol. e IRI (Magurran, 1988). Por último, para evaluar la estrategia alimentaria de la especie, se empleó un análisis gráfico propuesto por Costello (1990) basado en la posición relativa que ocupan los diferentes ítems en torno a un sistema cartesiano, en donde las abscisas corresponden al porcentaje de frecuencia de ocurrencia y las ordenadas al porcentaje de abundancia representada por la numerosidad. Para la representación gráfica, los porcentajes fueron transformados a raíz cuadrada según Zar (2010). Las 2 diagonales del gráfico representan la importancia de la presa (dominantes-raras) y la estrategia alimentaria del depredador (especialista-generalista).
ResultadosAnálisis de dieta. En un total de 70 tractos gastrointestinales, el 100% de los individuos consumieron estructuras vegetales no reproductivas, mientras que el 82.86% presentaron estructuras vegetales reproductivas. El cuadro 1 muestra las especies vegetales consumidas. Se contabilizaron un total de 1 710 estructuras reproductivas; incluyendo flores, frutos y semillas. En el 97% de los intestinos se hallaron nemátodos parásitos, no existiendo diferencia significativa entre estaciones respecto a su abundancia (prueba de Kruskal-Wallis, H (5)=9.77; p>0.08). El 28.57% de los tractos presentaron fragmentos de rocas en cantidad variable.
Especies vegetales encontradas en los tractos gastrointestinales de Phymaturus cf. palluma. FL: flores, FR: frutos, EST: estróbilos, SEM: semillas, H: hojas y T: tallos
| Especies vegetales | Machos | Hembras | Juveniles |
|---|---|---|---|
| Adesmia pinnifolia | FL-H | FL-H | FL |
| Artemisia mendozana | H-T | H-T | H |
| Erodium cicutarium | FL-FR-H-T | FL-H-T | FL-FR-H-T |
| Ephedra multiflora | EST-FR-T | EST-FR-H | |
| Gramineas | H | H | H |
| Guindilia aff. dissecta | H | H | |
| Lycium sp. | H | SEM-H | SEM |
| Phacelia secunda | H | H | |
| Oenothera piscensis | H | ||
| Schinus fasciculata | FR | ||
| Senecio aff. riojanus | H-FL | H-FL | H |
| Skhuria pinnata | FL | ||
| Tetraglochin alatum | FR | FR-H |
Se encontraron diferencias significativas en el consumo de hojas entre estaciones (prueba de Kruskal-Wallis, H (5)=29.24; p<0.0001), siendo diferentes las estaciones de otoño y verano 2010 (prueba de comparaciones múltiples K-W (35)=2.99, p<0.04). Mientras que no se observaron diferencias en el consumo de tallos entre estaciones (prueba de Kruskal-Wallis, H (5)=9.22, p>0.10).
Según el IRI, el ítem fundamental en la dieta de la especie fue: “flores” (%IRI= 100%), ubicándose “frutos y semillas” como accidentales (%IRI< 11%). La diversidad trófica calculada para la población fue de H'total=2.67. Mientras que el valor del índice de dominancia de la dieta fue de Λtotal=0.12.
En la figura 1 se observa la posición que ocupan los ítems alimentarios de la dieta de P. cf. palluma en el sistema cartesiano. Siguiendo el criterio de Costello (1990), los ítems alimentarios corresponden a presas raras debido a la baja frecuencia de ocurrencia y baja abundancia. Esta disposición de las presas, sitúan a la población hacia una fase generalista.

Estrategia alimentaria estudiada en P. cf. palluma, donde se muestran los ítems alimentarios de la dieta y se indican aquellos de mayor abundancia y %FO. A, Adesmia pinnifolia FR; B, flores pequeñas 1; C, flores pequeñas 2; D, Ephedra multiflora EST; E, Tetraglochum alatum FR; F, Eurodium cicutarum FL y G, Ephedra multiflora FR.
Análisis intrapoblacional de la dieta. Se encontraron un total de 722 estructuras vegetales reproductivas en hembras (n=32) y 927 en machos (n=29). El ítem “flores” fue el predominante en términos de %N y %FO en ambos sexos, seguido por “frutos” en menor proporción (Cuadro 2).
Se indican los ítems consumidos por machos, hembras y juveniles de P. cf. palluma, %N= porcentaje de numerosidad en el total de las presas, %FO= porcentaje de frecuencia de ocurrencia en el total de estómagos, %Vol.= porcentaje volumétrico de las presas, IRI= índice de importancia relativa
| Machos | Hembras | Juveniles | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ítems | %N | %FO | %Vol. | IRI | %N | %FO | %Vol. | IRI | %N | %FO | %Vol. | IRI |
| flores | 86.8 | 86.2 | 89.63 | 15 208.6 | 78.37 | 68.75 | 48.97 | 8 455.11 | 49.28 | 66.66 | 87.24 | 9 102.36 |
| frutos | 12.12 | 41.37 | 10.02 | 916.34 | 12.95 | 31.25 | 50.59 | 1 985.41 | 4.34 | 22.22 | 2.16 | 144.51 |
| semillas | 0.53 | 10.34 | 0.3 | 8.64 | 8.13 | 12.5 | 0.2 | 104.09 | 34.78 | 22.22 | 0.69 | 788.23 |
Con respecto al %Vol., “frutos” fue el ítem de mayor contribución en hembras, mientras que el ítem “flores” lo fue en los machos. Así, el ítem fundamental, tanto en hembras como en machos, fue “flores” (IRI= 100%), mientras que el resto de los ítems fueron accidentales en ambos sexos. El valor de diversidad trófica fue mayor en machos (H'Machos=2.37) respecto a hembras (H'Hembras= 2.03). Complementariamente, el índice de dominancia resultó menor en machos respecto a las hembras ΛMachos=0.16; ΛHembras=0.23). Teniendo en cuenta las estructuras vegetales reproductivas identificadas, resultaron fundamentales en machos “flores” de Senecio aff. riojanus y de Adesmia pinnifolia, y “frutos” de Tetraglochin alatum. Destacándose luego, como ítem secundario “estróbilos” de Ephedra multiflora y sus “frutos” como ítems accesorios. Particularmente, las hembras presentaron un único ítem fundamental “flores” de A. pinnifolia, resultando el resto de los ítems accidentales (Cuadro 1).
La similitud entre estaciones según el índice de Morisita-Horn, utilizando datos cuantitativos de IRI, resultó alta tanto en machos (M-H> 0.72) como en hembras (M-H> 0.84). Sin embargo, según el coeficiente de similitud de Jaccard (Ij= 0.14), la similitud cualitativa en la dieta de machos y hembras fue baja. Las estaciones de verano (2009 y 2010) fueron las de mayor similitud en machos (índice= 0.99). En cambio en las hembras, las estaciones de mayor similitud se agruparon en 2 situaciones independientes: primavera 2009-otoño 2010 (M-H=0.99) y verano 2010-otoño 2011 (M-H=0.99).
Respecto a los juveniles analizados (n=9), el 100% de los contenidos gastrointestinales presentaron estructuras vegetales no reproductivas, mientras que el 77.7% presentó estructuras vegetales reproductivas. Cerca del 90% de los intestinos presentaron nemátodos parásitos. Se encontraron fragmentos de rocas en mayor proporción respecto a adultos (Rocasjuveniles=44% vs. Rocasadultos=23%), sin alcanzar el 15% del total del contenido analizado. De las 61 estructuras vegetales reproductivas contabilizadas, el ítem alimentario “flores” resultó de mayor contribución según %N, %FO y %Vol., considerándose como ítem fundamental. A diferencia de los análisis en individuos adultos, el ítem “semillas” obtuvo mayor contribución que “frutos”; sin embargo, ambos resultaron ítems accidentales. La diversidad trófica en la dieta de los juveniles según el índice de Shannon-Wiener fue de H'Juveniles=2.03, mientras que la dominancia fue de ΛJuveniles=0.17.
Variación estacional de la dieta. Durante el primer periodo, en otoño 2010, el ítem “semillas” obtuvo mayor %N y %FO respecto a verano y primavera 2009, sin embargo, “flores” fue fundamental en todas las estaciones del periodo 1 como del periodo 2 (Cuadro 3). Verano 2009 presentó mayor diversidad trófica: (H'Primavera=1.85; H'verano=2.61; H'Otoño=1.89). Mientras que la estación más equitativa fue otoño 2010: ΛOtoño=0.22; ΛPrimavera=0.17; ΛVerano==0.11). Según los resultados obtenidos con el índice de similitud de Morisita-Horn con base en el %N y %IRI, indican que primavera y verano 2009 fueron las estaciones más similares (índice%N=0.92; índice %iri=0.99). En el segundo periodo, “flores” fue el único ítem presente en primavera 2010, resultando así la estación menos diversa: (H'Primavera=1.33; H'verano=2.02; H'otoño=1.59). Otoño 2011 resulto ser la estación de mayor dominancia de ítems: Λotoño=0.45; ΛPrimavera=0.22; Λverano=0.19. A diferencia del primer periodo, las estaciones más parecidas en %N y %IRI, fueron verano 2010 y otoño 2011(índice %N=1; índice%IRI=1). Finalmente, el coeficiente de similitud de Jaccard, indicó que las estaciones de verano de ambos periodos fueron las únicas que presentaron similitud en cuanto a sus ítems, sin embargo, el valor del índice fue mínimo (Ij=0.11).
Se indican los ítems consumidos por P. cf. palluma en las distintas estaciones pertenecientes a los 2 periodos de muestreo. %N= porcentaje de numerosidad en el total de las presas; %FO= porcentaje de frecuencia de presencia en el total de estómagos; %Vol.= porcentaje volumétrico de las presas; IRI= índice de importancia relativa
| 1° periodo | primavera 2009 | verano 2009 | otoño 2010 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ítems | %N | %FO | %Vol. | IRI | %N | %FO | %Vol. | IRI | %N | %FO | %Vol. | IRI |
| flores | 61.5 | 50 | 87.3 | 7 440.7 | 85.8 | 90 | 46.8 | 11 936 | 37.1 | 60 | 76.6 | 6 822.7 |
| frutos | 15.4 | 16.7 | 3.0 | 306.9 | 2.9 | 30 | 5.0 | 238.3 | 20.6 | 40 | 17.5 | 1 526.4 |
| semillas | 23.1 | 33.3 | 9.7 | 1 091.8 | 11.3 | 20 | 48.2 | 1 188.6 | 42.3 | 60 | 5.9 | 2 887.6 |
| 2°periodo | primavera 2010 | verano 2010 | otoño 2011 | |||||||||
| flores | 100 | 41.6 | 100 | 8 332 | 86.7 | 100 | 91.1 | 17 784.4 | 88.9 | 56.2 | 87.6 | 9 930.5 |
| frutos | — | — | — | — | 11.5 | 42.8 | 8.9 | 871.7 | 9.81 | 31.2 | 12.32 | 691.6 |
| semillas | — | — | — | — | 1.8 | 4.8 | 0.004 | 8.6 | 1.26 | 12.5 | 0.06 | 16.58 |
Phymaturus cf. palluma es una especie herbívora con tendencia al generalismo, que basa su dieta en estructuras vegetales reproductivas y no reproductivas. Exhibe una táctica activa de forrajeo, alimentándose de una gran variedad de especies vegetales en su dieta expresados por los altos valores de diversidad trófica encontrados. Según el análisis gráfico de la dieta (Costello, 1990), los ítems vegetales son poco frecuentes y proporcionados, sin embargo, los valores de %FO reflejan la tendencia de esta especie al consumo de flores. Esto coincide con Dutra et al. (2011) quienes establecen, para lagartos tropicales herbívoros, que las flores y frutos son las principales partes de la planta que se consumen, probablemente porque contienen menos material fibroso y celulosa que las estructuras verdes y por lo tanto, son más fácilmente digeridos y asimilados. Así, los lagartos pueden seleccionar plantas que les permitan obtener una dieta balanceada (Rocha, 2000) y a su vez contribuir y ser esenciales en la reproducción de algunas plantas y participar en los procesos de polinización y dispersión de semillas de manera exitosa (Olesen y Valido, 2003; Godínez-Álvarez, 2004; Galindo-Uribe y Hoyos-Hoyos, 2007).
Los géneros de plantas Artemisia y Ephedra resultaron las plantas más consumidas en una población de P. palluma en Mendoza (Videla, 1983). Mientras que para una población de P. antofagastensis en la cordillera de Catamarca, frutos de Lycium chanar y estróbilos de E. multiflora fueron los ítems principales de la dieta (Acosta et al., 2008). Por otra parte, en Chile en el Parque Nacional Laguna del Laja, P. aff. palluma consume los frutos de Berberis empetrifolia, Rumex acetosella y Calandrina sp., detectados por la presencia de semillas en las fecas (Celedón-Neghme et al., 2005).
Según Pough (1973) y Vitt (2004) se postula que la herbivoría ha surgido en especies de reptiles de gran tamaño y distribuidas en regiones de clima cálido. Sin embargo, la herbivoría en lagartos pequeños ha sido documentada en Argentina en lagartos, principalmente que habitan ambientes fríos (Quatrini et al., 2001; Valdecantos et al., 2012). Principios biofísicos sugieren que el pequeño tamaño en lagartos es una ventaja en ambientes de altitud y en latitudes elevadas, debido a que logran obtener la temperatura óptima más rápidamente, facilitando así un calentamiento más rápido en ambientes térmicos impredecibles. Por otra parte, la herbivoría probablemente es favorecida en climas fríos debido a los efímeros y menos abundantes que pueden ser los insectos como recurso alimenticio, en comparación con la posibilidad de fácil obtención de plantas como recurso en hábitats de alta montaña (Espinoza et al., 2004).
En este estudio, machos y hembras exhiben diferencias jerárquicas en el consumo de algunos ítems alimentarios, sin embargo, compartieron como ítem fundamental las “flores” de A. pinnifolia. En general, el consumo, variedad y aporte volumétrico de “flores” fue mayor en machos, contribuyendo a una mayor diversidad trófica en la dieta respecto a hembras. Esta observación podría relacionarse a una mayor selectividad por parte de los machos hacia las “flores”, o podría deberse a una mayor movilidad o mayor área de acción en los machos (Halloy y Robles, 2002). Generalmente los machos de Iguania tienen mayor área de acción que las hembras (Perry y Garland, 2002), característica posiblemente relacionada con los requerimientos energéticos, o con la necesidad de maximizar el acceso hacia las hembras. De esta manera, los machos podrían tener la posibilidad de seleccionar más variedad y cantidad de ítems para su consumo.
Cualitativamente, ambos sexos compartieron sólo el 14% de la dieta (Ij= 0.14), lo que indicaría que en esta población existe una baja competencia intraespecífica por el recurso alimenticio entre sexos. Por otro lado, en términos cuantitativos, las estimaciones de similitud a partir del IRI entre estaciones indicaron que todas las estaciones fueron altamente similares permitiendo afirmar que no existen grandes variaciones cuantitativas en la contribución de los diferentes ítems entre sexos.
En reptiles herbívoros, cambios ontogénicos han sido reportados en la dieta, concluyendo principalmente que los juveniles serían incapaces de consumir plantas por no tener la capacidad volumétrica necesaria para el procesamiento de una dieta vegetal (Fialho et al., 2000, Durstche, 2004). Contrario a esto, en este estudio, la dieta de los juveniles al igual que la de los adultos, fue una dieta estrictamente herbívora. No obstante, a diferencia de los adultos machos, ésta fue menos diversa. En términos generales, el ítem alimentario “flores” fue fundamental en la dieta. “Flores” de Skhuria pinnata se presentó como ítem exclusivo en juveniles.
Estacionalmente P. cf. palluma, únicamente presentó diferencias en el componente volumétrico y numérico en verano 2009 y otoño 2010, respectivamente, en ambos casos, las “semillas” resultaron de mayor contribución en la dieta, no obstante, el ítem fundamental fue “flores” entre fechas. Las estaciones de verano fueron las más diversas, lo que podría coincidir con épocas de mayor disponibilidad y oferta de alimento; y a nivel fenológico, son épocas de floración y fructificación de las distintas especies vegetales. La baja similitud encontrada en la dieta durante las estaciones de verano y la falta de similitud en el resto de las estaciones nos permite deducir que, la competencia estacional por el recurso alimentario sería muy leve dado el amplio espectro de ítems encontrados en las distintas fechas. Por otro lado, el análisis de Morisita-Horn para el IRI y %N mostró valores elevados en todas las estaciones, sin permitir establecer grandes variaciones cuantitativas en el tiempo.
Cabe destacar, que la información obtenida en este estudio representa un aporte substancial en el conocimiento de los hábitos tróficos de P. cf. palluma, especie que habita en ambientes rigurosos como son los de alta montaña, generando un avance en el conocimiento de la biología y ecología de la especie en ecosistemas andinos.
Agradecemos a la Subsecretaría de Medio Ambiente de la provincia de San juan por los permisos autorizados. A la Secretaría de Ciencia y Técnica de la UNSJ por el subsidio otorgado (CICITCA-JCAcosta). A Gustavo Fava por su asistencia en los trabajos de campo. A Antonio Dalmasso y Justo Márquez por su ayuda en la identificación de especies vegetales. A José Rogelio Cedeño Vázquez por su asistencia en el proceso editorial del manuscrito y a 2 revisores anónimos que contribuyeron a mejorar el trabajo.