



Se describe e ilustra una nueva especie de orquídea, Maxillaria pinasensis, de los bosques siempreverde piemontano y semideciduo montano del suroccidente de Ecuador. Se proporciona información sobre su distribución, hábitat, fenología y estatus de conservación. La nueva especie se compara con Maxillaria estradae de Ecuador, M. flava, M. lankesteri, M. microphyton y M. wercklei de Costa Rica y Panamá, a las cuales se parece morfológicamente.
Maxillaria pinasensis, a new orchid species from evergreen premontane and semi-deciduous montane forests from Southwestern Ecuador, is herein described and illustrated. Information about its distribution, habitat, phenology and conservation status is provided. The new species is compared with Maxillaria estradae from Ecuador, M. flava, M. lankesteri, M. microphyton and M. wercklei from Costa Rica and Panama, morphologically similar species.
El género Maxillaria Ruiz et. Pav. incluye aproximadamente 660 taxones (Govaerts, 2014) y es considerado como el grupo más numeroso de la subtribu Maxillariinae. Se distribuye desde México hasta Perú y Bolivia, incluyendo las Antillas, donde la mayoría de las especies crecen como epífitas en los bosques húmedos. En su circunscripción más amplia (Chase et al., 2015; Schuiteman y Chase, 2015), Maxillaria se diferencia de otros géneros de su subtribu porque sus integrantes, en muchos de los casos, presentan hojas con desarrollo no plicado, generalmente conduplicado, inflorescencias unifloras que surgen de la base del pseudobulbo y un labelo articulado al pie de la columna (Chase et al., 2015; Schuiteman y Chase, 2015).
Las relaciones filogenéticas de la subtribu Maxillariinae, o algunos de sus grupos, han sido evaluadas a partir del análisis de secuencias del ADN (Dathe y Dietrich, 2006; Ojeda, Carnevali, Williams y Whitten, 2003; Singer y Koehler, 2003; Szlachetko, Sitko, Tukallo y Mytnik-Ejsmont, 2012; Whitten et al., 2007). Estos estudios revelaron que Maxillaria era un grupo polifilético dentro de un clado altamente apoyado que incluía a otros géneros preliminarmente reconocidos en la subtribu, como Anthosiphon Schltr., Chrysocycnis Linden et. Rchb. f., Cryptocentrum Benth., Cyrtidiorchis Rauschert, Mormolyca Fenzl, Pityphyllum Schltr. y Trigonidium Lindl (Chase, Cameron, Barrett y Freudenstein, 2003). En su estudio sobre las relaciones filogenéticas de Maxillaria, Whitten et al. (2007) revelaron que los representantes de Maxillaria y los grupos antes mencionados se dividían en 17 clados. A partir de esos resultados, Blanco et al. (2007) redefinieron a Maxillaria con una delimitación estricta y elevaron a nivel genérico los otros grupos monofiléticos reconocidos en el análisis de Whitten et al. (2007). De esa forma, esos autores consideraron como géneros a Brasiliorchis R.B. Singer, S. Koehler et. Carnevali, Camaridium Lindl., Christensonella Szlach., Mytnik, Górniak et. Smiszek, Heterotaxis Lindl., Hylaeorchis Carnevali et. G.A. Romero, Inti M.A. Blanco, Mapinguari Carnevali et. R.B. Singer, Cyrtidiorchis Rauschert, Cryptocentrum Benth., Pityphyllum Schltr., Maxillariella M.A. Blanco et. Carnevali, Nitidobulbon Ojeda, Carnevali et. G.A. Romero, Rhetinantha M.A. Blanco y Sauvetrea Szlach. Esta clasificación de Maxillaria y grupos afines fue adoptada por Pridgeon, Cribb, Chase y Rasmussen (2009). Pese a tener una delimitación más estrecha, Maxillaria continuó siendo el género más numeroso en su subtribu.
Por su parte, Szlachetko et al. (2012) también evaluaron las relaciones filogenéticas de Maxillariinae y plantearon una nueva clasificación, aún más fraccionada para la subtribu, y elevaron a nivel genérico varios de los clados reconocidos dentro de los géneros más diversos considerados por Pridgeon et al. (2009), como Camaridium, Maxillaria y Ornithidium Salisb. ex R. Br. De esta manera, Szlachetko et al. (2012) concluyen reconociendo un total de 36 géneros dentro de la subtribu. Recientemente, Schuiteman y Chase (2015) propusieron una clasificación de Maxillaria a partir de la delimitación ampliada para el género, la cual no solo incluye a los grupos previamente segregados por Blanco et al. (2007), sino que se extendió para incluir también a Mormolyca, Ornithidium y Trigonidium. Con esta delimitación, Maxillaria resulta ser un megagénero que, además de los caracteres arriba mencionados, se puede reconocer porque sus integrantes, de manera general, presentan fibras esclerotizadas en las flores, la bráctea floral más larga que el ovario pedicelado y frutos con dehiscencia lateral. Por otra parte, los 17 grupos monofiléticos determinados en el análisis de Whitten et al. (2007) son considerados como secciones de Maxillaria por Schuiteman y Chase (2015): Maxillaria sect. Camaridium (Lindl.) Christenson, M. sect. Cryptocentrum (Benth.) Schuit. et. M.W. Chase, M. sect. Cyrtidiorchis (Rauschert) Schuit. et. M.W. Chase, M. sect. Digammae Christenson, M. sect. Erectae Pfitzer, M. sect. Iridifoliae Pfitzer, M. sect. Polyphyllae Christenson, M. sect. Mapinguari (Carnevali et. R.B. Singer) Schuit. et. M.W. Chase, M. sect. Maxillaria Ruiz et. Pav., M. sect. Nitidobulbon (Ojeda, Carnevali et. G.A. Romero) Schuit. et. M.W. Chase, M. sect. Ornithidium (Salisb. ex R.Br.) Christenson, M. sect. Pityphyllum (Schltr.) Schuit. et. M.W. Chase, M. sect. Repentes Pfitzer, M. sect. Rufescens Christenson, M. sect. Trigonae Christenson, M. sect. Trigonidium (Lindl.) Schuit. et. M.W. Chase y M. sect. Urceolatae Christenson. En este trabajo se sigue esta clasificación de Maxillaria, pues si bien ahora es un grupo más diverso y morfológicamente heterogéneo, es también monofilético, presenta un alto valor de soporte y, adicionalmente, se evitaría la proliferación de nombres genéricos que solo crearían confusión para el entendimiento del género (Bennett y Christenson, 2009; Chase et al., 2015; Schuiteman y Chase, 2015).
En Ecuador se ha registrado hasta el momento la presencia de más de 200 taxones de Maxillaria (Dodson, 2002), lo cual representa casi el 30% de la diversidad del género. Aproximadamente 50 de esas especies son endémicas de Ecuador (Jørgensen y León-Yánez, 1999). No obstante, excepto por Dodson (2002) y Dodson y Dodson (1980), los estudios taxonómicos para Maxillaria son escasos en el país. Recientemente, en la colección de Orquiecuador y Gloxinias, florecieron ejemplares que superficialmente son similares a Maxillaria estradae Dodson —endémica de Ecuador—, Maxillaria flava Ames, Hubbard et. C. Schweinf., Maxillaria lankesteri Ames, Maxillaria microphyton Schltr. y Maxillaria wercklei (Schltr.) L. O. Williams —especies de Costa Rica y Panamá—. Sin embargo, al examinar y comparar el material cultivado con esos taxones, se comprobó que correspondía a una planta diferente y desconocida, la cual se describe e ilustra aquí a partir de material vivo y herborizado.
Materiales y métodosLa descripción y la ilustración del nuevo taxón se elaboraron a partir de ejemplares cultivados en el centro de tenencia Orquiecuador y Gloxinias (Piñas-El Oro, Ecuador). Por otro lado, se tomaron fotografías a los individuos en floración usando una cámara Nikon 8700. Se realizó un dibujo a línea de uno de los ejemplares, usando para ello un microscopio USB 50-500x sobre una lámina milimetrada y una cámara Nikon D7100. El dibujo incluyó el hábito de la planta, la flor y la disección floral; las flores frescas fueron examinadas y diseccionadas en un microscopio. Se prepararon muestras de respaldo que fueron depositadas en el Herbario Ecuatoriano de Ciencias Naturales (QCNE) y en la colección particular de Orquiecuador y Gloxinias (J. Zambrano).
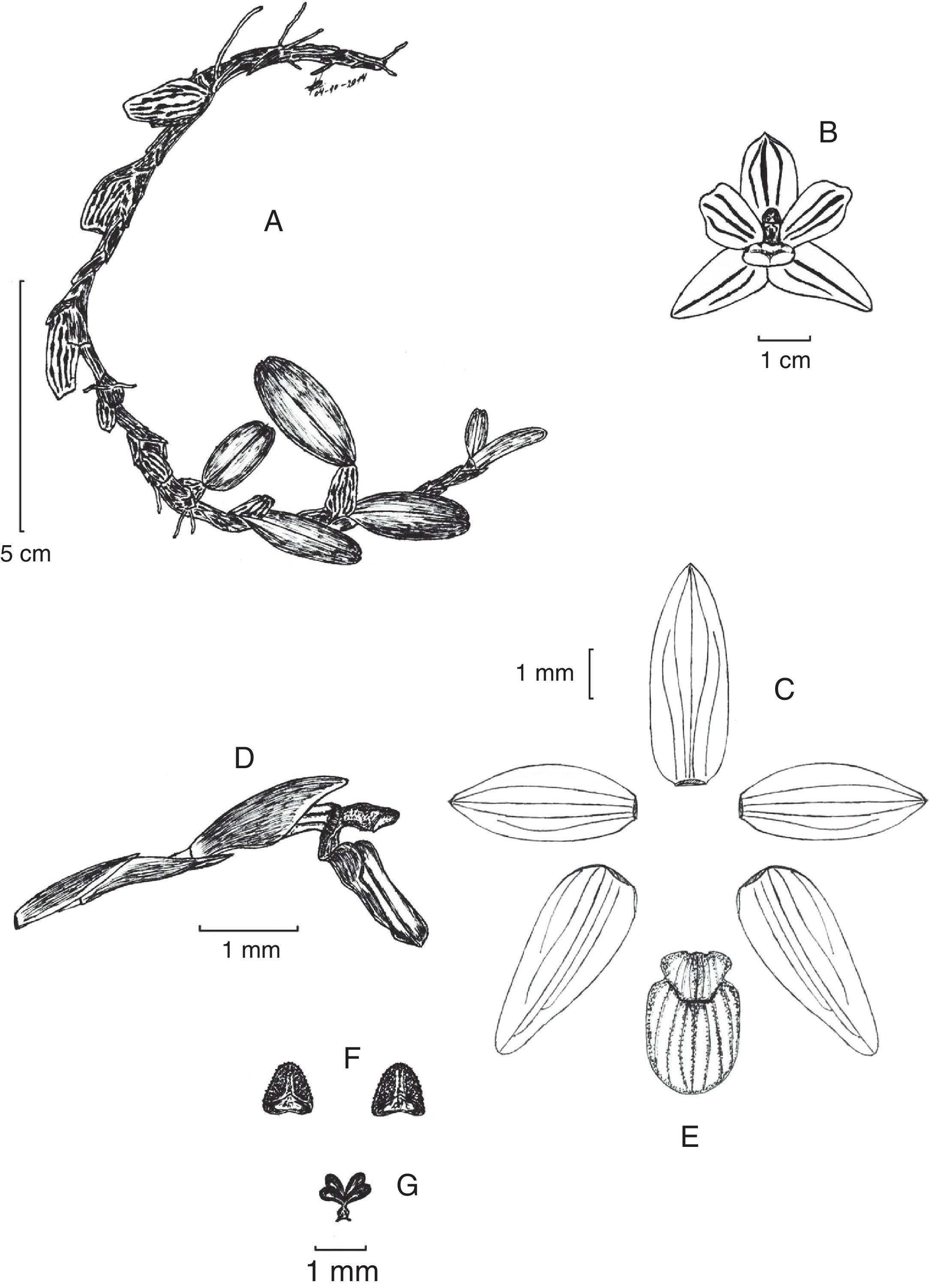
DescripciónMaxillaria pinasensis Zambrano et. Solano sp. nov. (figs. 1 y 2)
 hábito; B) vista de la flor; C) disección floral; D) labelo y columna en vista lateral; E) labelo en vista frontal; F) antera; G) polinario. Dibujo de B. Zambrano basado en el holotipo.")

Tipo. Ecuador. Provincia El Oro: Piñas, cordillera de Curitejo, 1,200 m snm, cultivado en Orquiecuador y Gloxinias, 20/XI/2007, J. Zambrano B. 432 (holotipo en QCNE; isotipo en Herbario J. Zambrano [flores conservadas en alcohol]).
Herba epiphytica, pseudobulbis ovoideis, striatis, unifoliatis. Folia oblongo-elliptica. Sepalum intermedium oblongo-lanceolatum; sepala lateralia ovato-lanceolata. Petala ovato-elliptica. Labellum 3-lobum, lobis lateralibus orbicularibus, leviter erectis, lobo intermedio orbiculari-obovato, apice rotundato. Columna leviter curvata.
Planta epifítica, de 25-30cm de longitud, rizomatosa, algo péndula, de 3-5cm de altura; rizoma cilíndrico, de 2-4.5cm de largo entre pseudobulbos adyacentes, cubierto por varias vainas agudas, escariosas, traslapadas. Raíces blanquecinas, delgadas. Pseudobulbos ovoides, longitudinalmente estriados, de 10-20mm de largo y 8-9mm de grosor, formados por un entrenudo, unifoliados en el ápice y rodeados en la base por 2-3 vainas foliáceas, conduplicadas, traslapadas. Hojas elípticas a oblongo-elípticas, ligeramente coriáceas, oblicuas, sésiles, articuladas al ápice del pseudobulbo, de 15-30×8-12mm, de ápice mucronado-emarginado. Inflorescencia pequeña, uniflora, desde la base del pseudobulbo maduro, 2-3 por pseudobulbo; pedúnculo erecto, cilíndrico, de 7mm de largo y 1mm de diámetro, envuelto por 3-4 brácteas agudas, escariosas, traslapadas, de 5-6×2mm —4mm cuando extendidas—. Bráctea floral más larga que el ovario pedicelado, cubriendo la base de la flor, conduplicada, obtusa, carinada, de 7-8mm de largo. Flores abiertas a semiabiertas, en forma de estrella; sépalos y pétalos amarillo-ámbar, con marcas rojo-púrpura a lo largo de las nervaduras, labelo rosado con manchas púrpura, columna blanquecina, antera crema con un ligera mancha púrpura en la cofia. Ovario pedicelado, ligeramente arqueado, cilíndrico, ensanchado hacia el ápice, de 6mm de largo y 1mm de diámetro. Sépalo dorsal de 9×3mm, ligeramente cóncavo, oblongo-lanceolado, obtuso, 5-nervados; sépalos laterales de 9×3.5mm, extendidos, ovado-lanceolados, ligeramente oblicuos, agudos, 5-nervados. Pétalos de 7.5×3mm, ligeramente cóncavos, elípticos a ovado-elípticos, agudos, 3-4-nervados. Labelo 3-lobulado, de 5.5×3.5mm, articulado al pie de la columna, 7-nervados, lóbulos laterales basales, orbiculares, ligeramente erectos, con un callo ligeramente ligulado a la mitad del labelo, el cual conecta los lóbulos laterales, lóbulo medio orbicular-obovado, con ápice redondeado. Columna corta, ligeramente arqueada, de 2mm de largo y 1mm de diámetro, con un pie prominente casi tan largo como el cuerpo de la columna. Antera subapical, ovoide, diminutamente papilosa; polinias 4, ovoides.
Ejemplares adicionales. Ecuador. Piñas, sector Panupali, vía a Machala, 458 m snm. 3/VIII/2013. Cultivado en Orquiecuador y Gloxinias, Zambrano B. 1060 (Herbario J. Zambrano); El Placer, km 12 vía a Machala, 567 m snm. 30/VI/2014. Cultivado en Orquiecuador y Gloxinias, Zambrano B. 1473 (Herbario J. Zambrano); sector Piñas Grande, 1,100 m snm. 5/VII2014. Cultivado en Orquiecuador y Gloxinias, Zambrano B. 1597 (Herbario J. Zambrano).
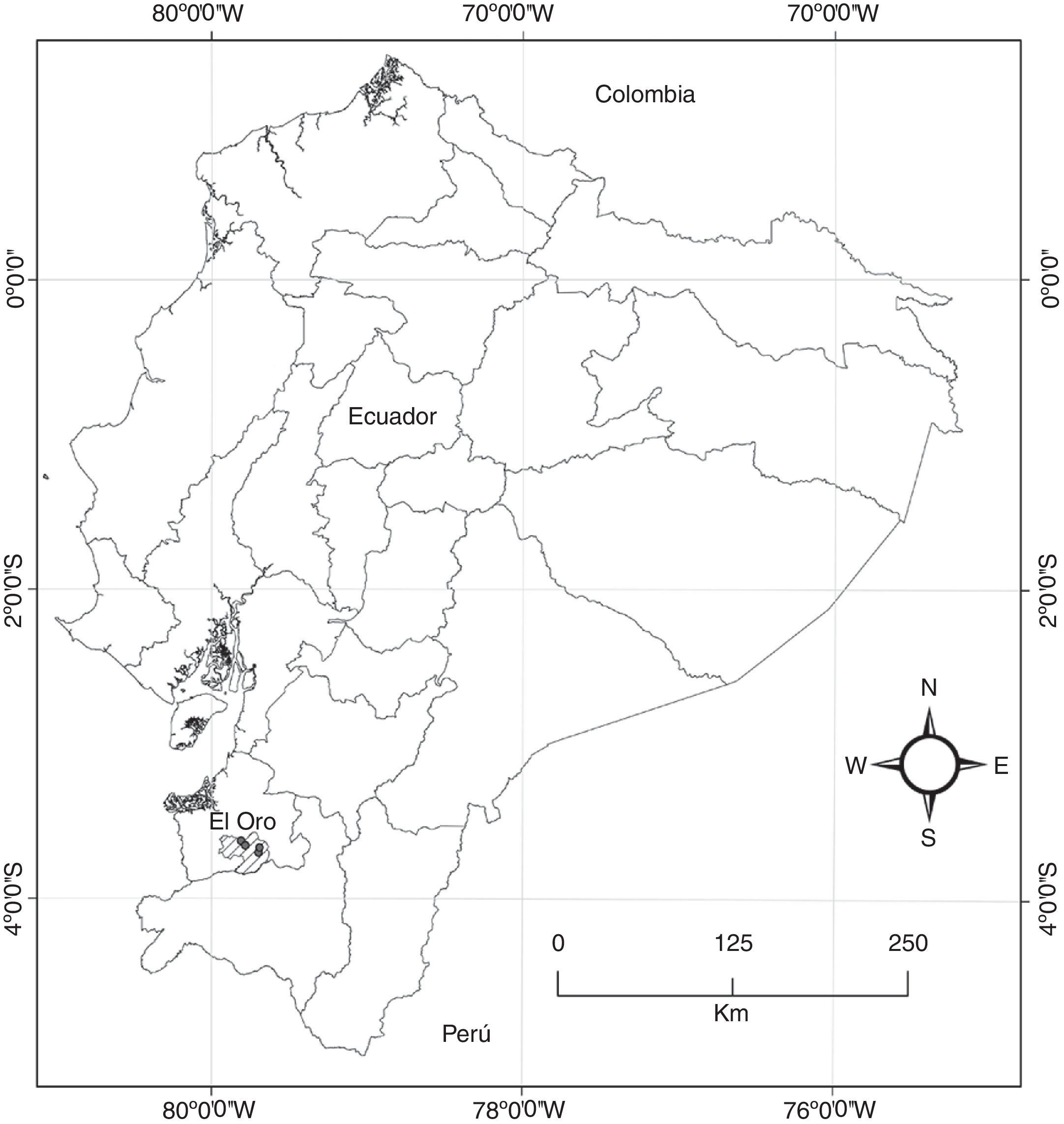
Distribución y hábitat. Hasta el momento esta especie es endémica de Ecuador, donde solo ha sido registrada en las montañas del suroccidente de la provincia de El Oro, entre los 400 y 1,100 m snm (fig. 3). Crece como epífita a medio dosel, sobre capas de musgos y materia orgánica acumulada en los troncos y ramas de arbustos (Citrus sp.) y árboles (Carapa sp., Ficus sp., Clusia sp. y Miconia sp.). El taxón habita en los bosques siempreverde piemontano y semideciduo montano localizados entre los 400 y 1,500 m snm, donde las neblinas son frecuentes durante todo el año, pero especialmente en la época seca.

Fenología. Florece entre junio y noviembre.
Etimología. El nombre de la especie es en alusión al cantón Piñas, localidad donde fue descubierta.
Estado de conservación. Esta especie presenta una distribución geográfica muy restringida. Hasta ahora se ha localizado en remanentes boscosos perturbados por la actividad humana, donde sus poblaciones están muy limitadas, presentan densidades bajas (2-11 individuos/árbol) y, además, parecen estar sujetas a eventos de extinción-recolonización frecuente. Debido a la fragmentación de su hábitat, las poblaciones hasta ahora conocidas —Curitejo, Panupali, El Placer, Piñas Grande— parecen estar aisladas y poco conectadas entre sí, por lo que se podría esperar que se presentase una estructura genética entre ellas. La única población cuya conservación podría estar garantizada es la que se encuentra en las zonas de influencia de la Reserva Buenaventura-El Placer; las otras poblaciones se localizan en áreas no protegidas.
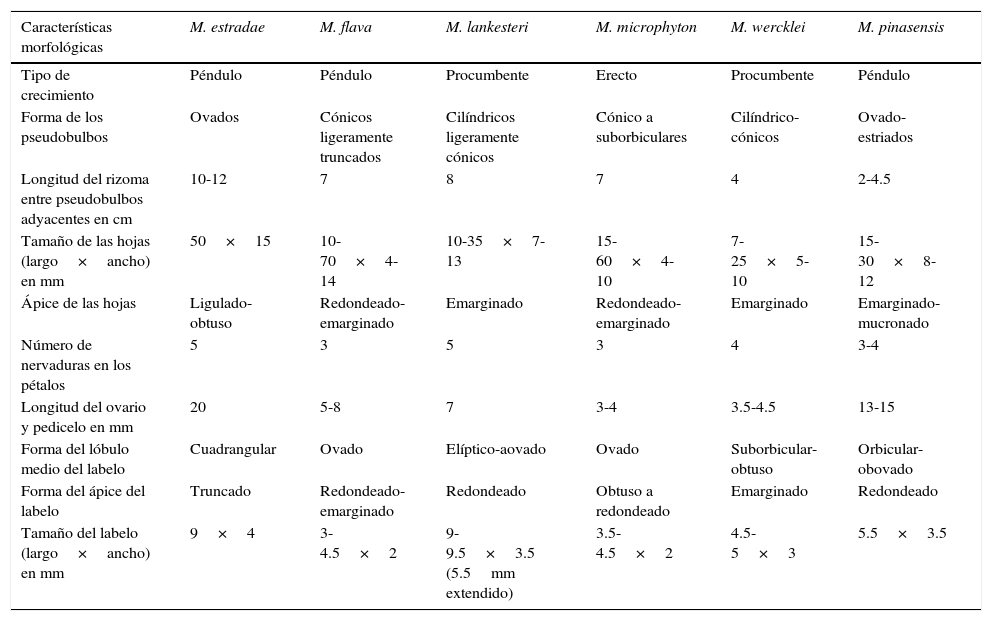
Comentarios taxonómicos. La especie del Ecuador más similar es la endémica M. estradae, la cual es diferente por su hábito más robusto, y evidentemente péndulo, pseudobulbos bifoliados, hojas más anchas (15 vs. 8-12mm), flores amarillo-rosadas (vs. amarillo-ámbar), segmentos del perianto más grandes y columna evidentemente arqueada (Dodson, 1980). Otros taxones morfológicamente afines son M. flava, M. lankesteri, M. microphyton y M. wercklei, originarios de Costa Rica y Panamá; sin embargo, la primera es diferente por sus pseudobulbos cónicos y algo truncados, hojas más largas (hasta 70 vs. 15-30mm), pedúnculo más corto (5-8 vs. 13-15mm, incluyendo el ovario pedicelado) y labelo ovado de 3-4.5×2mm (vs. orbicular-ovado de 5.5×3.5mm). Por su parte, M. lankesteri se distingue por sus plantas largamente rizomatosas (segmento de rizoma de 8cm entre pseudobulbos adyacentes vs. 2-4.5cm), pseudobulbos cilíndricos, pedúnculo más corto (7 vs. 13-15mm, incluyendo el ovario pedicelado) y el labelo elíptico-ovado (vs. orbicular-ovado). En cambio, M. microphyton posee pseudobulbos cónicos a suborbiculares, regularmente no surcados en ejemplares secos (vs. ovoides estriados), pétalos elíptico-lanceolados, 3-nervados (vs. elípticos a ovado-elípticos, 3-4-nervados). Finalmente, M. wercklei es diferente por sus pseudobulbos cilíndricos a ligeramente cónicos (vs. ovoides estriados), pedúnculo más corto (3.5-4.5 vs. 13-15mm, incluyendo el ovario pedicelado), pétalos elípticos-lanceolados (vs. elípticos a ovado-elípticos), labelo emarginado en el ápice (vs. redondeado) con los lóbulos laterales muy recurvados (vs. ligeramente erectos) (Atwood y Mora de Retana, 1999). La tabla 1 presenta un resumen de las principales diferencias entre las especies ya enunciadas.
Diferencias morfológicas de Maxillaria estradae, M. flava, M. lankesteri, M. microphyton, M. wercklei y M. pinasensis.
| Características morfológicas | M. estradae | M. flava | M. lankesteri | M. microphyton | M. wercklei | M. pinasensis |
|---|---|---|---|---|---|---|
| Tipo de crecimiento | Péndulo | Péndulo | Procumbente | Erecto | Procumbente | Péndulo |
| Forma de los pseudobulbos | Ovados | Cónicos ligeramente truncados | Cilíndricos ligeramente cónicos | Cónico a suborbiculares | Cilíndrico-cónicos | Ovado-estriados |
| Longitud del rizoma entre pseudobulbos adyacentes en cm | 10-12 | 7 | 8 | 7 | 4 | 2-4.5 |
| Tamaño de las hojas (largo×ancho) en mm | 50×15 | 10-70×4-14 | 10-35×7-13 | 15-60×4-10 | 7-25×5-10 | 15-30×8-12 |
| Ápice de las hojas | Ligulado-obtuso | Redondeado-emarginado | Emarginado | Redondeado-emarginado | Emarginado | Emarginado-mucronado |
| Número de nervaduras en los pétalos | 5 | 3 | 5 | 3 | 4 | 3-4 |
| Longitud del ovario y pedicelo en mm | 20 | 5-8 | 7 | 3-4 | 3.5-4.5 | 13-15 |
| Forma del lóbulo medio del labelo | Cuadrangular | Ovado | Elíptico-aovado | Ovado | Suborbicular-obtuso | Orbicular-obovado |
| Forma del ápice del labelo | Truncado | Redondeado-emarginado | Redondeado | Obtuso a redondeado | Emarginado | Redondeado |
| Tamaño del labelo (largo×ancho) en mm | 9×4 | 3-4.5×2 | 9-9.5×3.5 (5.5mm extendido) | 3.5-4.5×2 | 4.5-5×3 | 5.5×3.5 |
Los taxones de Centroamérica arriba mencionados son morfológicamente más similares a M. pinasensis que la ecuatoriana M. estradae, pero la gran disyunción geográfica entre sus poblaciones y los ecosistemas diferentes en los que estas crecen tanto en Ecuador como en Panamá y Costa Rica son criterios para considerar a los ejemplares ecuatorianos como una especie diferente. Por otro lado, hasta ahora no se sabe que la planta descrita aquí como nueva, o alguna muy similar, hayan sido registradas en zonas adyacentes de Colombia y Perú. Cabe recalcar que la especie aquí descrita solo ha sido encontrada al suroccidente del Ecuador, en una zona muy restringida de bosques húmedos rodeados en su mayoría por bosques secos.
Maxillaria pinasensis pertenece a M. sect. Camaridium (Lindl.) Christenson, la cual se distingue por su pequeño labelo y perianto ampliado, con una bráctea floral que se extiende hasta sobreponerse a la base del sépalo dorsal. Tiene los pétalos similares en tamaño y forma a los sépalos, la columna corta y arqueada, un labelo 3-lobulado, con un callo prominente dirigido hacia la base del labelo, cóncavo, de aspecto no brillante (Blanco et al., 2007; Schuiteman y Chase, 2015; Szlachetko et al., 2012; Whitten et al., 2007).
Agradecemos a los revisores anónimos por los comentarios y sugerencias que mejoraron la calidad del manuscrito, al Dr. Fernando Chiang por sus valiosas observaciones y corrección de la descripción en latín, así como al Ministerio del Ambiente Ecuatoriano (MAE) por la autorización de investigación científica N.° 006-IC-FLO-DPAEO-MAE.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.