





Fungal infections have become a concern for health professionals, and the emergence of resistant strains has been reported for all known classes of antifungal drugs. Among the fungi causing disease, we highlight those that belong to the genus Aspergillus. For these reasons, the search for new antifungals is important. This study examines the effects of a coumarin derivative, 4-acetatecoumarin (Cou-UMB16) both alone and together with antifungal drugs, and its mode of action against Aspergillus spp. Cou-UMB16 was tested to evaluate its effects on mycelia growth, and germination of Aspergillus spp. fungal conidia. We investigated its possible action on cell walls, on the cell membrane, and also the capacity of this coumarin derivative to enhance the activity of antifungal drugs. Our results suggest that Cou-UMB16 inhibits Aspergillus spp. virulence factors (mycelia growth and germination of conidia) and affects the structure of the fungal cell wall. When applying Cou-UMB16 in combination with azoles, both synergistic and additive effects were observed. This study concludes that Cou-UMB16 inhibits mycelial growth and spore germination, and that the activity is due to its action on the fungal cell wall, and that Cou-UMB16 could act as an antifungal modifier.
In recent years, fungal infections have become a concern for health professionals, and the emergence of resistant strains has been reported for all known classes of antifungal drugs. As a result, especially in immunocompromised patients, mortality, morbidity, and the cost of treatment against fungal pathogens have all increased. Among the disease-causing fungi, we highlight those that belong to the genus Aspergillus. Aspergillus fumigatus is the most common species isolated in cases of invasive aspergillosis, the species Aspergillus flavus, Aspergillus niger, and Aspergillus terreus follow close behind. In certain locations, A. flavus and A. terreus species are frequently isolated.1,2
A. fumigatus, being saprotrophic, plays an essential role in recycling carbon and nitrogen worldwide. The fungus has a high capacity to sporulate which results in an ubiquitous presence, both indoors and outdoors, of high atmospheric conidia concentrations (1–100conidia/m3). A. fumigatus is a prevalent airborne fungal pathogen, and causes severe and usually fatal invasive infections in immunocompromised hosts.3
A. flavus is the second leading cause of invasive aspergillosis and is the most common cause of superficial fungal infections. Particularly common clinical syndromes associated with A. flavus include chronic granulomatous sinusitis, keratitis, cutaneous aspergillosis, wound infections, and osteomyelitis (following trauma and inoculation). In addition, A. flavus produces aflatoxins, the most toxic and potent natural hepatocarcinogenic compounds ever characterized.4
Until recently, the only drugs available to treat aspergillosis were amphotericin B and itraconazole, the latter in oral and intravenous formulations. Recently voriconazole, posaconazole, and caspofungin have also been approved for treatment. Although resistance to antifungal drugs is not as common as resistance to antibacterial agents, there has been a significant increase in the number of reported cases of fungal resistance. Some investigators have isolated strains of A. flavus from patient samples that are resistant to amphotericin B and itraconazole.1,4
For these reasons, the search for new antifungals is important. In work recently reported by our group, the antifungal activity of 24 coumarin derivatives against A. fumigatus and A. flavus were described. Certain derivatives showed significant antifungal activity with Minimum Inhibition Concentration (MIC) values ranging from 16 to 1024μg/mL, including 4-acetatecoumarin (Fig. 1) with an MIC value of 16μg/mL.6
.")
These results motivated us to assess more thoroughly the activity of this coumarin derivative on the fungal cell. Another promising alternative is the possibility of combining coumarin derivatives with antifungals against strains of Aspergillus spp. Combination therapy of the available antifungal drugs with natural products and their derivatives is even less explored, especially in the case of coumarin derivatives.6,7 The aim of this study was to examine the effects of coumarin derivative, 4-acetetatecoumarin, both alone and together with antifungal drugs and its mode of action against Aspergillus spp.
Material and methodsMicroorganismsAspergillus spp. used in the antifungal assay was obtained from the archival collection of the Federal University of Paraíba Laboratory of Mycology (LM). They included A. fumigatus ATCC 46913 and A. flavus ATCC 16013. Stock inoculates (suspensions) of Aspergillus spp. were prepared from 8-day old potato dextrose agar (Difco Lab., USA); the cultures grown at room temperature. Fungal colonies were covered with 5mL of sterile saline solution (0.9%), the surface was gently agitated with vortexes. The fungal elements with saline solution were transferred to sterile tubes. Inoculate was standardized at 0.5 tube of McFarland scale (106CFU/mL). The final concentration confirmation was done by counting the microorganisms in a Neubauer chamber.8,9
ChemicalsThe product tested was the coumarin derivative, 4-acetatecoumarin (Cou-UMB16), obtained by biosynthesis.5 Amphotericin B, fluconazole, itraconazole and voriconazole were obtained from Sigma Aldrich®, Brazil. The drugs were dissolved in DMSO (dimethylsulfoxide), and sterile distilled water was used to obtain solutions of 1024μg/mL for each antifungal. The concentration of DMSO did not exceed 0.5% in the assays.
Culture mediaTo test the biological activity of the products, Potato Agar (AP), Sabouraud dextrose agar (SDA) (purchased from Difco Laboratories, Detroit, MI, USA), and RPMI-1640-L glutamine (without sodium bicarbonate) (Sigma–Aldrich, São Paulo, SP, Brazil) culture media were used. They were prepared and used according to the manufacturers’ instructions.
Effects on mycelia growthAnalysis of the interference of the coumarin derivative, 4-acetatecoumarin, on mycelia growth was performed by determining the dry mycelia weights of A. fumigatus (ATCC 46913) and A. flavus (ATCC 16013).11,12 Flasks containing MIC (16μg/mL) and MIC×2 (32μg/mL) of coumarin derivative in RPMI-1640 medium, were inoculated with suspension of the A. fumigatus (ATCC 46913) and A. flavus (ATCC 16013) test strains. In the corresponding control, the amount of coumarin derivative was substituted by distilled water. The system was incubated at 28°C for 8 days. Flasks containing mycelia were filtered through Whatman® Grade 1 Qualitative Filtration Paper (particle retention: 11μm), and then washed with distilled water. The mycelia were dried at 60°C for 6h, and kept at 40°C overnight. The filter paper containing dry mycelia from two independent assays were weighed, and the mean values obtained. The percentage growth inhibition based on the dry weight of each at time of analysis, was calculated according to Sharma and Tripathi.10
Conidial germination assayThe coumarin derivative, 4-acetatecoumarin (Cou-UMB16), and amphotericin B were tested to evaluate effects on the germination of fungal conidia of A. fumigatus (ATCC 46913), and A. flavus (ATCC 16013). Flasks containing MIC (16μg/mL) and MIC×2 (32μg/mL) of coumarin derivative and a control with distilled water, were used. In sterile test tubes, 500μL of RPMI-1640 plus the Cou-UMB16 were evenly mixed with 50μL of fungal conidia suspension and immediately incubated at 28°C. Samples of this mixture were taken after 24h of incubation for analysis. The experiment was performed in duplicate, where the 200 conidia was analyzed in a Neubauer chamber, and the inhibition percentage of spore germination at each time point was calculated by comparing the results obtained in the test experiments with the results of the control experiment. The analysis was conducted under an optical microscope (Zeiss® Primo Star).13,14
The effects of sorbitol on growth after exposure to antifungal agentsThe assay was performed using medium with and without sorbitol to evaluate possible mechanisms involved in the antifungal activity of the test product on the Aspergillus spp. cell wall. The sorbitol was added to the culture medium in a final concentration of 0.8M. The assay was performed by microdilution method in 96-well plates in a “U” (Alamar, Diadema, SP, Brazil).8,9 The plates were sealed aseptically, incubated at 28°C, and readings were taken at 3 days. Based on the ability of sorbitol to act as a fungal cell wall osmotic protective agent, the higher MIC values observed in the medium with sorbitol added as compared to the standard medium suggest that the cell wall is one of the possible cell targets for the product tested.15,16 Amphotericin B was used as the control drug. The assay was performed in duplicate and expressed as the geometric mean of the results.
Ergosterol binding assay – MIC value determination in the presence of ergosterolTo assess if the product binds to fungal membrane sterols, an experiment was performed according to the method described by Escalante,16 with some modifications. The ergosterol was prepared as was described by Leite et al.15 The MIC of coumarin derivative, 4-acetatecoumarin (Cou-UMB16) against Aspergillus spp. was determined by using broth microdilution techniques,8,9 in the presence and absence of exogenous ergosterol (Sigma–Aldrich, São Paulo, SP, Brazil) added to the assay medium, in different lines of the same micro-plate. Briefly, a solution of Cou-UMB16 was doubly diluted serially with RPMI-1640 (volume=100μL) containing ergosterol at a concentration of 400μg/mL. A volume of 10μL yeast suspension (0, 5 McFarland) was added to each well. The same procedure was realized for amphotericin B, whose interaction with membrane ergosterol is already known, serving as a control drug. The plates were sealed and incubated at 28°C. The plates were read after 3 days of incubation, and the MIC was determined as the lowest concentration of test agent inhibiting the visible growth. The assay was carried out in duplicate and the geometric mean values were calculated. Thus, this binding assay reflected the ability of the compound to bind with ergosterol.
Checkerboard assayA checkerboard micro-titer test was performed to evaluate the interaction of coumarin derivative 4-acetatecoumarin with the antifungal drugs (azoles and amphotericin B) against A. fumigatus ATCC 46913, and A. flavus ATCC 16013. A series of 2-fold dilutions in eight for each coumarin derivative and antifungal drug were made in RPMI-1640 to obtain four times the final concentration in micro-titer wells. Then, 50μL of each dilution of coumarin derivative was added to the 96-well micro-titer plates in the vertical direction, while 50μL of each dilution of the antifungal drugs was added in the horizontal direction, so that various combinations of coumarin derivative and antifungal drugs could be achieved. In addition, 10μL of inoculum from the spore suspension (1.5×105CFU/mL) was added to each well, and the plates were incubated at 30°C for 3 days. In order to evaluate the activity of the combinations of drugs, fractional inhibitory concentration (FIC) indices were calculated as FICA+FICB, where FICA and FICB represent the minimum concentrations inhibiting the fungal growth for drugs A and B, respectively: FICA=MICA combination/MICA alone and FICB=MICB combination/MICB alone. A mean FIC index was calculated based on the following equation:
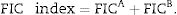
In addition, the interpretation was made as follows: synergistic (<0.5), additive (0.5–1.), indifferent (>1), or antagonistic (>4).5,17–19
Data analysisThe results were expressed as mean±SE. Statistical analyses were performed with t-test. p<0.05 was considered significant.
ResultsThe coumarin derivative, 4-acetatecoumarin, Fig. 1, with a minimum inhibitory concentration (MIC) (16μg/mL) has been previously observed by our group,6 and displayed inhibitory activity against spore germination and mycelial growth of A. fumigatus ATCC 46913 and A. flavus ATCC 16013.
The percentage of conidia germinated by A. fumigatus ATCC 46913 and A. flavus ATCC 16013 in the presence and absence (control) of test products is shown in Fig. 2. In the test with two concentrations (MIC and MIC×2), Cou-UMB16 showed significant inhibitory action against Aspergillus spp. spore germination (p<0.05) when compared to the control. However, the activity was less potent (p<0.05) compared with respective concentrations of amphotericin B.
, A, and A. flavus (ATCC 16013), B, in the absence (control) and presence of 4-acetatecoumarin (MIC: 16μg/mL; 2×MIC: 32μg/mL) and amphotericin B (MIC: 2μg/mL; 2×MIC: 4μg/mL). a: p<0.05 compared to control. b: p<0.05 compared to amphotericin B with respective concentration.")
Percentage of conidial germination of A. fumigatus (ATCC 46913), A, and A. flavus (ATCC 16013), B, in the absence (control) and presence of 4-acetatecoumarin (MIC: 16μg/mL; 2×MIC: 32μg/mL) and amphotericin B (MIC: 2μg/mL; 2×MIC: 4μg/mL).
a: p<0.05 compared to control. b: p<0.05 compared to amphotericin B with respective concentration.
The effect of different concentrations of test drugs (at MIC and MIC×2) on mycelial growth was determined by dry mycelial mass quantification; the results are shown in Fig. 3. With respect to effects on A. fumigatus ATCC 46913 and A. flavus ATCC 16013, concentrations of Cou-UMB16 at 16μg/mL (MIC), and 32mL−1 (MIC×2) inhibited mycelial growth of normal strains (p<0.05) when compared to the control. Further analysis demonstrated that Cou-UMB16 at its MIC×2 was more potent than amphotericin B at its respective MIC×2 (p<0.05).
, A, and A. flavus (ATCC 16013), B, in the absence (control) and presence of 4-acetatecoumarin (MIC: 16μg/mL; 2×MIC: 32μg/mL) and amphotericin B (MIC: 2μg/mL; 2×MIC: 4μg/mL). Control produced 100% of dry mycelia weight. a: p<0.05 compared to control. b: p<0.05 compared to amphotericin B with respective concentration.")
Percentage of dry mycelia weight produced by A. fumigatus (ATCC 46913), A, and A. flavus (ATCC 16013), B, in the absence (control) and presence of 4-acetatecoumarin (MIC: 16μg/mL; 2×MIC: 32μg/mL) and amphotericin B (MIC: 2μg/mL; 2×MIC: 4μg/mL). Control produced 100% of dry mycelia weight.
a: p<0.05 compared to control. b: p<0.05 compared to amphotericin B with respective concentration.
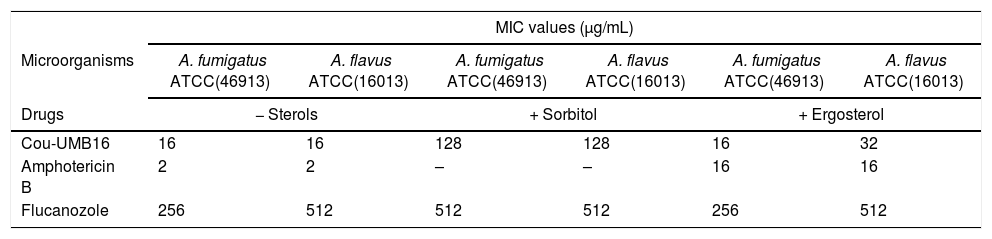
In this study, the MIC values of Cou-UMB16 in culture media with or without exogenous ergosterol were identical in both experiments, suggesting that the coumarin derivative tested does not act via binding to plasma membrane ergosterol; results shown in Table 1.
MIC values (μg/mL) of coumarin derivative, 4-acetatecoumarin (Cou-UMB16), in the absence and presence of sorbitol (0.8M) and ergosterol (400μg/mL) against Aspergillus spp.
| MIC values (μg/mL) | ||||||
|---|---|---|---|---|---|---|
| Microorganisms | A. fumigatus ATCC(46913) | A. flavus ATCC(16013) | A. fumigatus ATCC(46913) | A. flavus ATCC(16013) | A. fumigatus ATCC(46913) | A. flavus ATCC(16013) |
| Drugs | − Sterols | + Sorbitol | + Ergosterol | |||
| Cou-UMB16 | 16 | 16 | 128 | 128 | 16 | 32 |
| Amphotericin B | 2 | 2 | – | – | 16 | 16 |
| Flucanozole | 256 | 512 | 512 | 512 | 256 | 512 |
To investigate the action on the cell wall of the coumarin derivative Cou-UMB16 we carried out a test with sorbitol (as an osmotic shield); the test results are shown in Table 1. The Cou-UMB16 MIC values increased 8 times in the presence of culture medium with sorbitol compared to culture medium without sorbitol, suggesting that Cou-UMB16 indeed acts on the structure of the fungal cell wall, results similar to other coumarins analyzed by our group.20
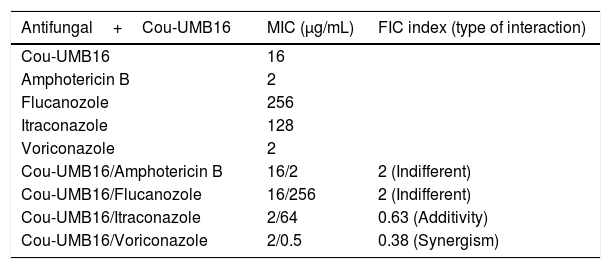
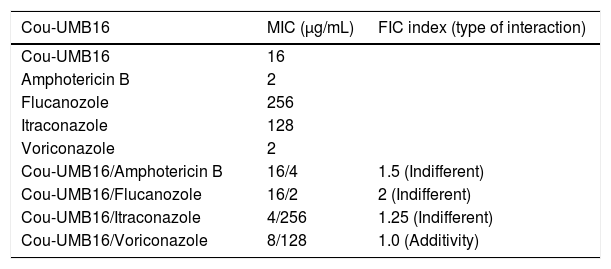
Tables 2 and 3 show the results of Cou-UMB16 in combination with the antifungal drugs (amphotericin B, and azole derivatives), against A. flavus ATCC 16013 and A. fumigatus ATCC 46913. The combination of Cou-UMB16 with voriconazole against A. flavus ATCC 16013 revealed synergistic effects. Against A. flavus ATCC 16013 additive effects were observed for the combination of Cou-UMB16 with itraconazole; the fractional inhibitory concentration index (FIC) being equal to 0.63. With voriconazole against A. fumigatus ATCC 46913, the FIC index was 1.0. However, the combination of Cou-UMB16 and amphotericin B and fluconazole revealed an FIC index greater than 1.0 for all strains tested.
MIC of antifungal drugs and effect of combination with coumarin derivative, 4-acetatecoumarin (Cou-UMB16), against A. flavus, ATCC 16013.
| Antifungal+Cou-UMB16 | MIC (μg/mL) | FIC index (type of interaction) |
|---|---|---|
| Cou-UMB16 | 16 | |
| Amphotericin B | 2 | |
| Flucanozole | 256 | |
| Itraconazole | 128 | |
| Voriconazole | 2 | |
| Cou-UMB16/Amphotericin B | 16/2 | 2 (Indifferent) |
| Cou-UMB16/Flucanozole | 16/256 | 2 (Indifferent) |
| Cou-UMB16/Itraconazole | 2/64 | 0.63 (Additivity) |
| Cou-UMB16/Voriconazole | 2/0.5 | 0.38 (Synergism) |
Note: Cou-UMB16, 4-acetatecoumarin; FIC, fractional inhibitory concentration; MIC, minimal concentration inhibitory.
MIC of antifungal drugs and effect of combination with coumarin derivative, 4-acetatecoumarin (Cou-UMB16), against A. fumigatus, ATCC 46913.
| Cou-UMB16 | MIC (μg/mL) | FIC index (type of interaction) |
|---|---|---|
| Cou-UMB16 | 16 | |
| Amphotericin B | 2 | |
| Flucanozole | 256 | |
| Itraconazole | 128 | |
| Voriconazole | 2 | |
| Cou-UMB16/Amphotericin B | 16/4 | 1.5 (Indifferent) |
| Cou-UMB16/Flucanozole | 16/2 | 2 (Indifferent) |
| Cou-UMB16/Itraconazole | 4/256 | 1.25 (Indifferent) |
| Cou-UMB16/Voriconazole | 8/128 | 1.0 (Additivity) |
Note: Cou-UMB16, 4-acetatecoumarin; FIC, fractional inhibitory concentration; MIC: minimal concentration inhibitory.
Current antifungal treatments against Aspergillus spp. are still effective, but resistant strains and intrinsically resistant species are emerging fast. Current antifungal drugs suffer from a number of limitations that can render their use difficult; e.g. amphotericin B's dose-limiting nephrotoxicity; and azoles’ simple fungistatic mode of action (with the development of fungal resistance). This reveals the urgent need for new antifungals with broad fungicidal spectrums of action, and with fewer dose-limiting side effects.22,23
An important field of study for new antifungal drugs is plant biosynthesis of secondary metabolites (and their derivatives). The coumarin derivative, 4-acetatecoumarin (Cou-UMB16); a biosynthetic compound, has reported antifungal activity.6
Coumarins are a group of compounds are derived from phenylalanine metabolism. They reveal a benzene ring attached to a pyran ring, the simplest and the most common representative found being coumarin (1,2-benzopyrone). These metabolites are found throughout the world. Coumarins have been isolated in various plant parts; the leaves, roots, and fruits of several different families such as: Fabaceae, Asteraceae, Rutaceae, Saxifragaceae and Thymelaceae.23
Various coumarins and their derivatives have already proven their antifungal activity. It is believed that these properties are usually a reflection of their initial roles in vegetables; such as the phytoalexins.24–27 The interesting biological activities of the coumarins have made them attractive targets for organic synthesis.6,27,28
This study shows that 4-acetatecoumarin (Cou-UMB16) has activity against mycelial growth and spore germination for A. fumigatus ATCC 46913 and A. flavus ATCC 16013. Aspergillus spp. are ubiquitous to the soil and produce large numbers of asexual conidia (conveyed by the environment), which then become important factors for the spread of the fungus and consequent host infections.29
The coumarin derivative, Cou-UMB16, before the assays, was dissolved in DMSO (dimethylsulfoxide), and sterile distilled water was used to obtain initial solution of 1024μg/mL. The concentration of DMSO did not exceed 0.5% in the assays, secured that it does not interfere with antifungal activity of coumarin.29,30
The coumarin derivative, Cou-UMB16 displayed inhibitory action against Aspergillus spp. spore germination, thus interfering in this important virulence factor. Hyphae are difficult to phagocytize, and can induce apoptosis in macrophages (after phagocytosis) if forming inside. The resulting mycelium formation is an important virulence factor for Aspergillus spp.31,32 Reductions in mycelial growth (produced by Cou-UMB16) interfere with Aspergillus spp. virulence. This result was similar to that found in our previous study with other coumarin derivatives.21
The MIC values for Cou-UMB16 were multiplied 4 times in the presence of sorbitol dosed culture medium as compared to normal culture medium, suggesting that Cou-UMB16 acts on the structure of the fungal cell wall. Studies show that coumarin acts via formation of pores in the cell wall, a consequent release of cytoplasmic contents, and cell death; this corroborates the results found in our study.21,24 However, other studies show that coumarins alter the mitochondrial morphology of the fungal cell, thus inducing apoptosis.23
The synergistic effect observed against A. flavus ATCC 16013 when combining Cou-UMB16 and voriconazole; and the additive effects against A. flavus ATCC 16013 when combined with itraconazole; and against A. fumigatus ATCC 46913 when combined with voriconazole, suggest that Cou-UMB16 may be used in future combinations with these drugs. Several reports have indicated yet other antifungal combinations, being assayed in vitro and then applied in clinics,33–35 and with differing plant derivatives,7 but against Aspergillus spp., combinations of synthetic drugs with coumarin derivatives are little explored.
ConclusionOur results suggest that the coumarin derivative 4-acetatecoumarin (Cou-UMB16) has antifungal activity which inhibits virulence factors in the Aspergillus species tested. The results also demonstrate that the action of the coumarin derivative is against the structure of the fungal cell wall. When applying Cou-UMB16 in combination with azoles (voriconazole and itraconazole), both synergistic and additive effects were observed, depending on the fungal strain used.
Conflicts of interestThe author(s) declare(s) that there are no conflicts of interest regarding the publication of this paper.
The authors are grateful to CAPES (Coordenação de Aperfeiçoamento Pessoal de Nível Superior), the Federal University of Paraíba (UFPB) for financial support, and to David Harding who edited the English version.