





La enfermedad cardiovascular es la manifestación clínica de un proceso que se inicia en las primeras décadas de la vida, la aterosclerosis1, y que evoluciona de forma asintomática y, en general, sin expresión clínica hasta la edad adulta. La presencia de factores de riesgo en la edad infantil se ha relacionado con la incidencia de cardiopatía isquémica
(CI) en el adulto2. En el Estudio Cuatro Provincias hemos analizado diversos factores de riesgo cardiovascular en niños de 6 a 8 años de 4 zonas españolas que presentan diferencias importantes en las tasas de mortalidad por CI en la edad adulta: Madrid, Orense, Cádiz y Murcia3. Uno de los datos interesantes de nuestro estudio es el referente a los elevados valores plasmáticos de colesterol unido a lipoproteínas de alta densidad (cHDL) en nuestros niños4, que han permanecido altos y estables en los últimos años5. Estos elevados valores de cHDL en niños pueden estar contribuyendo a explicar la baja mortalidad cardiovascular existente en España a pesar de la elevada presencia de factores de riesgo6.
Al efecto protector asociado a las HDL por su participación en el transporte inverso del colesterol, se añade el efecto antioxidante que las HDL ejercen sobre las lipoproteínas de baja densidad (LDL)7,8. Estas propiedades antioxidantes están vinculadas a la presencia de enzimas antioxidantes en la partícula HDL, tales como la paraoxonasa, capaces de frenar el proceso oxidativo de las LDL, retrasando o previniendo el desarrollo del proceso aterosclerótico9. La paraoxonasa 1 (PON1) es una éster hidrolasa que destruye los fosfolípidos e hidroperóxidos oxidados presentes en la LDL10-12, destruyendo de esta forma su actividad biológica y restaurando una LDL normal para acoplarse a su receptor. La región codificante del gen de la PON1 presenta 2 polimorfismos mayoritarios: uno de ellos originado por una sustitución arginina por glutamina en el codón 192 (192 Q/R)13, considerado como el determinante mayor de la actividad sé-rica de la paraoxonasa14, y el otro originado por una sustitución metionina/leucina en la posición 55 (55 M/L)15.
La actividad PON1 del suero se ha encontrado disminuida en sujetos con infartos de miocardio16,17 y con hipercolesterolemia familiar18, y se ha asociado a riesgo cardiovascular aumentado19. Sin embargo, la asociación de los polimorfismos con la enfermedad no es tan consistente; así, hemos encontrado un grupo de población en Canadá donde la mortalidad por CI es baja y el alelo PON-192 R está aumentado20, y sin embargo, no se observa esta asociación en poblaciones estudiadas en Italia21 o Francia22. La discrepancia en los resultados hace necesario estudiar los distintos aspectos relacionados con la paraoxonasa en cada población para evaluar su relación con el riesgo cardiovascular, así como analizar los factores que puedan estar relacionados con su actividad.
Por todo ello, en el presente trabajo nos propusimos analizar los valores de actividad PON1 y los polimorfismos PON1 192Q/R y PON1 55M/L en los niños de edad prepuberal que integran el Estudio Cuatro Provincias y evaluar la relación de estos aspectos con las variables antropométricas y bioquímicas en nuestra población.
Material y métodosDiseño y sujetos de estudio
El estudio se llevó a cabo en muestras de suero y ADN de 1.275 niños de 6 a 8 años (media 6,7 años) integrantes del Estudio Cuatro Provincias, estudio transversal, destinado a analizar factores de riesgo cardiovascular en población infantil de 4 provincias españolas con distintas tasas de mortalidad por CI3. La selección de los niños del Estudio Cuatro Provincias se realizó mediante muestreo aleatorio, por conglomerados de los centros escolares, estratificados por sexo y nivel socioeconómico. El protocolo del estudio cumple con las normas de Helsinki y la legislación española sobre investigación clínica enhumanos, y ha sido aprobado por el Comité Ético de Investigación Clínica de la Fundación Jiménez Díaz de Madrid. Se remitió una carta a los padres de los niños con información sobre los objetivos y procedimientos del estudio y se obtuvo la autorización escrita de los padres. Se excluyó a los niños que, según comunicado de los padres, padecían alteraciones metabólicas, endocrinas, hepáticas o renales.
Determinación de la actividad paraoxonasa
La actividad paraoxonasa en suero se determinó mediante espectrofotometría, según el método descrito por Mackness et al18 con mínimas modificaciones. La mezcla de reacción consta de 500 µl de una solución con una concentración de sustrato paraoxón de 2,22 mmol/l en buffer 0,1 mol/l Tris-HCl a pH 8,0 que contenía 2 mmol/l de CaCl2 y 50 µl de suero. La generación de p-nitrofenol se mide según su absorbancia a una longitud de onda de 405 nm a 37 °C, de forma continua en un espectrofotómetro. La actividad PON1 se expresó en U/l.
Determinación de los genotipos de paraoxonasa
Polimorfismo 192. Se determinó según la técnica descrita por Humbert et al23 en 1993. Como producto de la reacción en cadena de la polimerasa (PCR) se obtiene un fragmento de 99 pares de bases que es digerido con la enzima de restricción Alw I. Los productos de esta digestión se separan por medio de electroforesis en gel de acrilamida al 12%. En la posición 192 la sustitución puntual de arginina por glutamina crea un único sitio de corte para la enzima Alw I; el alelo Q (glutamina) corresponde a un fragmento sin digerir de 99 pares de bases, mientras que el alelo R (arginina) origina 2 fragmentos, uno de 65 pb y otro de 34 pb.
Polimorfismo 55. Se determinó, al igual que el polimorfismo 192, por la técnica descrita por Humbert et al23; como producto de PCR se obtiene un fragmento de 170 pares de bases que será digerido con la enzima Hsp 92 II; los productos de la digestión serán separados por electroforesis en gel de acrilamida al 8%. En la posición 55 puede aparecer una metionina o una leucina, el alelo M (metionina) corresponde a un fragmento no digerido de 170 pares de bases, y el alelo L (leucina) da lugar a 2 fragmentos, uno de 126 y otro de 44 pares de bases15.
Método estadístico
La comparación de las frecuencias de genotipos y alelos entre provincias se realizó mediante la prueba de la µ2. Las diferencias entre provincias se evaluaron mediante el análisis ANOVA. Dado que las distribuciones de los valores de actividad PON1 en suero no siguieron una distribución normal, los análisis estadísticos se llevaron a cabo después de su transformación logarítmica. Las correlaciones entre actividad paraoxonasa y las variables bioquímicas se estimaron mediante coeficientes de correlación de Spearman después de ajustar por peso. El análisis de cómo las distintas variables o su interacción afectan a los valores de actividad paraoxonasa se realizó mediante modelos de regresión lineal. Los análisis se realizaron con el programa SPSS versión 9.0.
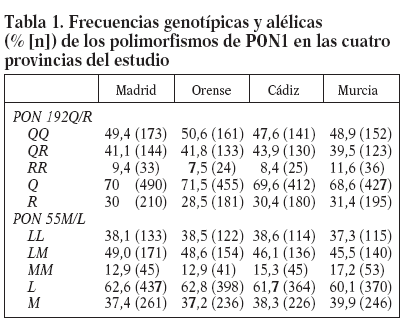
ResultadosLas frecuencias genotípicas y alélicas de los polimorfismos PON1 192Q/R y PON155 M/L en los niños del estudio se muestran en la tabla 1. No existen diferencias significativas en la distribución de ninguno de los genotipos o alelos en las 4 provincias. La frecuencia media del alelo Q fue del 70% y la del alelo R, del 30%. Las frecuencias medias de los alelos L y M fueron del 62 y el 38%, respectivamente.
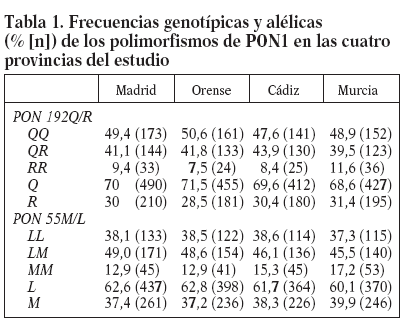
Tabla 1. Frecuencias genotípicas y alélicas (% [n]) de los polimorfismos de PON1 en las cuatro provincias del estudio
Los valores de actividad paraoxonasa de nuestra población fueron de 280,5 ± 168,6 U/l, con valores comprendidos entre 48,1 y 871,9 U/l. No hemos observado diferencias significativas entre sexos, con valores medios de actividad de 288,5 ± 173,9 U/l en niños y de 272,5 ± 162,8 U/l en niñas. Hemos observado que la actividad paraoxonasa era significativamente más elevada en niños obesos (310,7 ± 161,4 U/l) que en niños con peso normal (275,2 ± 169,1 U/l).
La actividad paraoxonasa presentó una distribución bimodal en las 4 provincias del estudio. Tal como era de esperar, la actividad paraoxonasa, tanto en el grupo total como en cada una de las provincias por separado, fue significativamente superior (p < 0,001) en los portadores del genotipo RR que en los portadores del genotipo QR y, en ambos casos, significativamente superior a los portadores del genotipo QQ. De igual modo, los portadores del alelo L presentaban valores de actividad paraoxonasa significativamente más elevados que los porres de actividad paraoxonasa en función de la provincia, observamos diferencias importantes en esas actividades, tanto al comparar los valores totales como los de cada uno de los genotipos en cada una de las provincias (tabla 2). Los valores de actividad paraoxonasa en Orense fueron significativamente más elevados que en el resto de las provincias, tanto en el conjunto de niños como al comparar los portadores de los genotipos QQ, QR, RR y LL por separado. Por el contrario, Murcia presenta los valores de actividad paraoxonasa más bajos tanto en el total de la población como al comparar los valores de cada genotipo.

Tabla 2. Actividad enzimática (U/l) asociada a los distintos genotipos de PON1 en las cuatro provincias del estudio
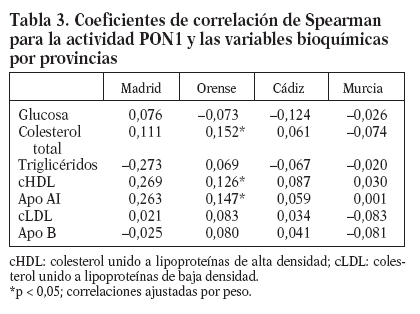
Mediante el análisis de correlación de Spearman, observamos que en la provincia de Orense existen correlaciones significativas (p < 0,05) entre la actividad PON1 en suero y las concentraciones de CT, cHDL y apo-AI después de ajustar por peso (tabla 3).
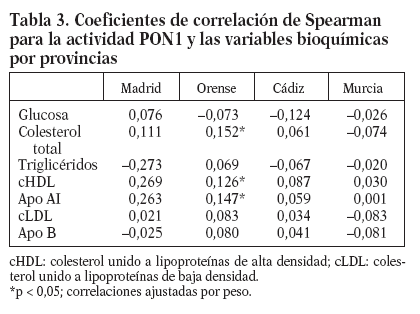
Tabla 3. Coeficientes de correlación de Spearman para la actividad PON1 y las variables bioquímicas por provincias
El análisis de regresión ha mostrado que los polimorfismos PON 192Q/R y PON 55 M/L son los principales determinantes de la actividad paraoxonasa en nuestra población infantil en las cuatro provincias del estudio. De modo que los polimorfismos PON 192Q/R y PON 55 M/L aparecen como predictores significativos en Orense (en modelos con R2 de 0,814 incluyendo solamente el polimorfismo PON 192Q/R y con R2 de 0,832 al incorporar el polimorfismo PON 55 M/L) y en Murcia (en un modelo con R2 de 0,830 con PON 192Q/R como único predictor y de 0,841 incluyendo PON 192Q/R y PON 55 M/L). En Madrid y Cádiz el polimorfismo PON 192Q/R aparece como predictor único en modelos con R2 de 0,862 y 0,802, respectivamente.
DiscusiónEn nuestro estudio de factores de riesgo cardiovascular en la edad infantil (Estudio Cuatro Pro-vincias)3, y ante los elevados valores de cHDL observados en nuestros niños prepuberales4,5, decidimos analizar las implicaciones de este hallazgo no solamente en cuanto a la concentración de cHDL en sí, sino en cuanto a las características antioxidantes de la partícula, estudiando los valores de actividad paraoxonasa 1 (PON1) y los polimorfismos de PON1 en nuestra población prepuberal.
El valor medio de actividad paraoxonasa de nuestra población es de 280,5 U/l, similar en ambos sexos. Se trata de valores inferiores a los observados en adultos españoles, para los que se han descrito valores medios de actividad paraoxonasa de 410,9 U/l24, pero más elevados que los descritos en otras poblaciones tanto de adultos como de niños. Por ejemplo, se han descrito valores medios de 219,6 U/l en adultos sanos del Reino Unido25, de 162 U/l en la población control japonesa26 y de 79,5 U/l en la adultos de China27 o valores medios de 124,8 U/l en niños griegos de 4 a 6 años28 y de 97,5 U/l en niños de 11-12 años en Eslovaquia29. En nuestro estudio hemos observado que la actividad paraoxonasa presenta una distribución bimodal, idéntica a la descrita para otras poblaciones16. El polimorfismo PON1 192Q/R se considera el determinante mayor de la actividad sérica de la paraoxonasa30 y, por tanto, sería la causa de la determinación genética de esta distribución bimodal31.
Al analizar la prevalencia del alelo menos común para este polimorfismo PON192R, hemos encontrado una frecuencia del 30%. Se trata de una frecuencia idéntica a la descrita en nuestro país en estudios en adultos32 y similar a las observadas en otros países europeos33 pero muy diferente de las descritas en poblaciones asiáticas como la japonesa (64%) o la china (63%)33. Sin embargo, no hemos encontrado diferencias en la frecuencia de este polimorfismo, ni del polimorfismo PON1 55M/L entre las provincias de nuestro estudio. Como se ha comentado previamente los estudios que analizan la relación de este polimorfismo con la enfermedad cardiovascular son discordantes incluso dentro de una misma población34,35. En la actualidad se piensa que la actividad paraoxonasa se relaciona de forma más evidente con la enfermedad que los polimorfismos genéticos36.
Al comparar los grados de actividad paraoxonasa en las provincias de nuestro estudio, hemos observado que Orense presenta los valores más elevados y Murcia los más bajos. Curiosamente Orense es la provincia donde los valores de actividad PON1 correlacionan con los valores plasmáticos de colesterol total, cHDL y apo AI. La asociación entre la actividad PON1 y los valores plasmáticos de cHDL y apo AI se ha descrito en estudios en adultos27, y, sin embargo, no se ha encontrado en estudios en niños29. Además, se ha descrito que la correlación entre la actividad PON1 y la concentración de cHDL es más débil en población con enfermedad coronaria que en población control37, sugiriendo que existen factores que modulan la relación entre la enzima y la HDL en distintas situaciones o que la asociación puede depender de los valores de actividad PON1.
Análisis de regresión han confirmado que los niveles de actividad PON1 en nuestra población están determinados genéticamente, de modo que los polimorfismos, especialmente el polimorfismo PON1 192Q/R, explican un alto porcentaje (alrededor del 80%) de la variabilidad en la actividad PON1. A pesar de que puedan existir factores ambientales que influyan sobre la actividad PON138, nuestros datos están de acuerdo con datos muy recientes de la literatura científica que atribuyen a los determinantes genéticos un impacto mucho mayor sobre PON1 que el de los determinantes ambientales39, con estudios en adultos que atribuyen al genotipo el 76% de la variación de actividad PON130.
En resumen, aunque existe una importante determinación genética, la diferente actividad PON1 observada en niños de las provincias de Orense y Murcia, para todos los genotipos en el rango determinado por el propio genotipo, sugiere la existencia de factores que modulan la actividad paraoxonasa en de cada genotipo que deben ser investigados.
Investigadores del Estudio Cuatro Provincias:J.L. del Barrio, I. de Oya (Fundación Jiménez Díaz,Universidad Autónoma de Madrid); M.A. Lasunción, H. Ortega (Hospital Ramón y Cajal); J.M. Martín Moreno, L. Gorgojo (Universidad de Valencia); M.A. Royo (Instituto de Salud Carlos III); A. Gil (Universidad Rey Juan Carlos); F. Rodríguez Artalejo (Universidad Autónoma de Madrid); O. Fernández (Hospital Cristal Piñol, Orense); A. Mangas y A. Macías (Universidad de Cádiz) y J. Fernández Pardo (Hospital de la Cruz Roja de Murcia).
*Al final del artículo se relacionan los investigadores del Estu dio Cuatro Provincias. Este trabajo ha sido parcialmente subvencionado por la beca concedida por la FEA/SEA para la Investigación Clínico-Epi- demiológica en el año 2003, en el XVI Congreso Nacional de la Sociedad Española de Arteriosclerosis, celebrado en Marbella.
Correspondencia: Dra. C. Garcés. Unidad de Lípidos. Fundación Jiménez Díaz. Avda. Reyes Católicos 2, 28040 Madrid. España. Correo electrónico: cgarces@fjd.es
Recibido el 1 de agosto de 2007 y aceptado el 7 de septiembre de 2007.