





La apolipoproteína CIII (apoCIII) inhibe la hidrólisis de triglicéridos de la VLDL y quilomicrones por la lipasa lipoproteica. Se ha pensado que la hipertrigliceridemia resultante explica la asociación entre el aumento de la apoCIII con exageración del riesgo cardiovascular. Sin embargo, estudios clínicos indican que un aumento de la apoCIII asociada a las LDL es un factor de riesgo independiente de la hipertrigliceridemia. Experimentos recientes indican que la apoCIII incrementa la adhesión de monocitos al endotelio, un requisito para la acumulación de macrófagos en la aterogénesis. Además, LDL ricas en apoCIII tienen una alta afinidad por los proteoglicanos de la íntima, lo cual podría aumentar su deposición subendotelial. Estas dos propiedades y efectos de la LDL rica en apoCIII pueden ser la base de los hallazgos indicativos de que la apoCIII es un agente causal de la aterogénesis y un factor de riesgo cardiovascular independiente.
The apolipoprotein CIII (apoCIII) is an inhibitor of the hydrolysis of triglycerides in chylomicrons and VLDL by lipoprotein lipase. It has been accepted that the hypertriglyceridemia induced by high apoCIII can explain its association with increased cardiovascular risk. However, several studies indicate that increased apoCIII is a risk factor independent of hypertriglyceridemia. Recent experimental evidence in vitro and in vivo indicates that apoCIII by itself, or associated with lipoproteins, increase expression of cell adhesion molecules in the endothelium and monocytes. Thus causing associations between these cells. In addition, it has been shown that LDL rich in apoCIII have a high affinity for proteoglycans of the arterial intima. A property that could increase the rate of LDL entrapment in the subendothelial intima. These two properties of LDL rich in apoCIII indicate that apoCIII is a significant risk factor because it can have a causal role in atherogenesis.
La apolipoproteína CIII (apoCIII) es una proteína pequeña sintetizada por el hígado y los enterocitos en el intestino delgado. La apoCIII es secretada por el hígado asociada a las lipoproteínas de muy baja densidad (VLDL) y por el intestino en la linfa como parte de los quilomicrones. Esta apolipoproteína fue aislada y caracterizada inicialmente por Brown, Levy y Fredrickson1. La apoCIII es una de las apolipoproteínas más abundantes del plasma y es un inhibidor fisiológico de la enzima lipoproteína lipasa (LPL), la cual circula por el plasma y además se encuentra asociada a la superficie del endotelio del tejido adiposo, del músculo esquelético y del músculo cardiaco. La LPL es la principal responsable de la hidrólisis de los triglicéridos (TG) de las VLDL y de los quilomicrones2. Aumentos de la secreción de la apoCIII pueden inducir hipertrigliceridemias, que se consideran factores de riesgo de la enfermedad cardiovascular ateroesclerótica2. Por muchos años se ha aceptado que la asociación significativa de niveles elevados de apoCIII circulante con el aumento de riesgo cardiovascular está relacionada con la hipertrigliceridemia. Sin embargo, hallazgos recientes indican que la apoCIII por sí misma, o unida a las lipoproteínas, aumenta la aterogenicidad de estos complejos2,3. En esta revisión discutiremos los resultados que apoyan la interpretación de estos hallazgos, así como las hipótesis propuestas acerca de los mecanismos por los cuales la apoCIII confiere una mayor aterogenicidad a las VLDL y especialmente a las lipoproteínas de baja densidad (LDL). Haremos hincapié en estudios recientes que muestran cómo la apoCIII puede tener efectos aterógenos directos sobre los monocitos y el endotelio, así como en la deposición de la LDL en la íntima de la pared arterial. Por último, mencionaremos los esfuerzos para el desarrollo de fármacos que puedan modular la contribución de la apoCIII a la ateroesclerosis y sus manifestaciones clínicas.
Evidencia clínica que relaciona la apoCIII con dislipidemias y aterogénesisEl hallazgo de que la apoCIII puede inhibir la hidrólisis de los TG de la VLDL y de los quilomicrones circulantes por la LPL, y con ello inducir hipertrigliceridemias, fue el estímulo inicial para la extensa investigación dedicada a esta apolipoproteína4. La inhibición de la LPL lleva a una disminución de la producción de ácidos grasos a partir de los TG circulantes. Tal inhibición causa el aumento de la vida media metabólica de los TG en el plasma y genera hipertrigliceridemia posprandial y también postabsortiva, lo cual hace de la apoCIII un modulador crítico en la generación de ácidos grasos a partir de los TG provenientes de la dieta (quilomicrones) y del hígado (VLDL). Como consecuencia, aumentos de la apoCIII pueden limitar el acceso y la utilización de los ácidos grasos requeridos para las funciones fisiológicas de los adipocitos, el músculo esquelético y el músculo cardíaco2,4-6.
La apoCIII circula en la sangre como isoformas de una cadena proteica de 8.920dalton (Da) de peso molecular, que puede tener unidas covalentemente una o 2 moléculas de ácido siálico y adquirir pesos moleculares de 9.120 y 9.420Da, respectivamente, y que se designan apoCIII0, apoCIII1 y apoCIII22. El contenido de la apolipoproteína en plasma se puede medir fácilmente usando anticuerpos específicos y nefelometría o ELISA. Los valores normales son de 10-15mg/dl7.
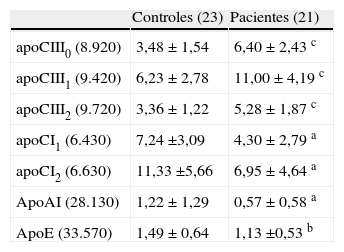
Estudios en humanos publicados a partir de 1990 han mostrado una asociación significativa entre valores elevados (mayores de 15mg/dl) de la apoCIII en la circulación con dislipidemias caracterizadas por cambios a todo lo largo del espectro de las lipoproteínas circulantes tales como aumento de las VLDL y, por consiguiente, hipertrigliceridemia, disminución de las lipoproteínas de alta densidad (HDL), especialmente de las partículas grandes (HDL-α), y aumento de LDL densa y pequeña (LDLdp). Este perfil lipoproteico es común en la insulinorresistencia (IR), el síndrome metabólico (SM) y en la diabetes tipo 2, y se considera la principal causa del aumento del riesgo cardiovascular presente en sujetos con estos trastornos metabólicos8,9. Los resultados de los principales trabajos clínicos publicados hasta diciembre de 2011 se resumen en la tabla 110-20. La mayoría de los estudios citados son relativamente pequeños, pero la consistencia de sus resultados sugiere que la apoCIII es un factor de riesgo cardiovascular real. Sin embargo, no es fácil distinguir si las asociaciones encontradas son causadas por la misma apoCIII o si son producto de que la apoCIII elevada casi siempre acompaña a los cambios múltiples de la dislipidemia de la IR descrita. No obstante, en los estudios de Blakenhorn et al.10, Chivot et al.11, Hodis et al.12 y Sacks et al.15,16 los niveles de apoCIII mostraron una correlación con el riesgo cardiovascular estadísticamente independiente de la hipertrigliceridemia y de la hipercolesterolemia. Es importante resaltar que la contribución aparente de los niveles de apoCIII al riesgo cardiovascular es mucho más significativa cuando se mide la apolipoproteína asociada a las partículas contentivas de apoB-100 (VLDL, remanentes de la VLDL y en la LDL) que cuando se mide su contenido total en plasma. Más aún, estudios de nuestros laboratorios mostraron que en pacientes con el síndrome metabólico y enfermedad ateroesclerótica periférica la presencia de niveles altos de la apoCIII caracterizan a una subclase de las LDL, las LDL pequeñas y densas (LDLpd) de los pacientes, comparándolas con controles aparentemente sanos y apareados17. Utilizando técnicas proteómicas pudimos demostrar que el aumento de la apoCIII por partícula de LDLpd incluye todas las isoformas de la apolipoproteína (tabla 2). Por otra parte, las apolipoproteínas CI, CII, AI y E mostraron estar más elevadas en los controles comparados con los pacientes. Debemos señalar que en nuestro estudio el número de pacientes y de controles fue relativamente pequeño debido a lo laborioso de la separación de las subclases de las LDL y de los análisis proteómicos17.
Estudios que muestran la asociación de niveles altos de apoCIII con dislipidemias y/o aumento del riesgo de enfermedad cardiovascular
| Referencia | Pacientes | Estudio | Resultado |
| Blakenhorn et al. (1990)10 | 162 H con EC | Aleatorizado, placebo-control, angiografía | Aumento apoCIII aumento progresión |
| Chivot et al (1990)11 | 74 H con EC; 76 controles | Caso-control | Aumento apoCIII aumento hipertrigliceridemia |
| Hodis et al (1994)12 | 220 H/M con EC y dislipidemia | Aleatorizado, placebo-control sección-cruzada | Aumento apoCIII-apoB predicción independiente de la progresión |
| Luc et al. (1996)13 | 360 H/M post IM; 489 controles sanos | Caso-control | Aumento apoCIII-en infarto |
| Gervaise et al. (2000)14 | 188 hombres con diabetes tipo 2 | Sección-cruzada | Riesgo por EC aumentado 3,95 con apoCIII >17mg/dl |
| Sacks et al.(2000)15 | 418 (87% H) con IM; 370 controles (87% H) | Prospectivo, 5 años | Riesgo episodio coronario aumentado 2,25x quint. 5 de apoCIII (VLDL+LDL) |
| Lee et al. (2003)16 | 121 diabéticos con IM; 121 sin IM (83% H) | Prospectivo, 5 años | LDL con alta apoCIII aumento riesgo de IM 9,3x |
| Davidsson et al. (2005)17 | 31 M con IR y ateroesclerosis periférica; 31 controles sanas | Sección-cruzada | LDL pequeña con aumento apoCIII marcador de aterosclerosis |
| Onat et al. (2009)18 | 395 H, 464 M, 42% con síndrome metabólico | Prospectivo, 5 años | ApoCIII en HDL y no-HDL significativamente asociado con MS y EC |
| Olivieri et al. (2010)19 | 633 (81% H) con EC (angiograf.) | Prospectivo, 5 años; 91 fallecimientos, 64 IM | apoCIII >10,5mg/dl aumenta el riesgo 2,2x |
| Mendivil et al. (2011)20 | 749 casos; 749 controles (76% M) originalmente sin EC | Prospectivo, 10-14 años | Riesgo episodio coronario aumentado 2,38x,quintil mas alto de apoCIII en LDL |
H: hombres; M: mujeres; IM: infarto de miocardio; IR: insulinorresistencia; EC: enfermedad coronaria.
Apolipoproteínas en las LDL pequeñas y densas de pacientes (mujeres) con insulinorresistencia y ateroesclerosis periférica y controles sanos
| Controles (23) | Pacientes (21) | |
| apoCIII0 (8.920) | 3,48 ± 1,54 | 6,40 ± 2,43 c |
| apoCIII1 (9.420) | 6,23 ± 2,78 | 11,00 ± 4,19 c |
| apoCIII2 (9.720) | 3,36 ± 1,22 | 5,28 ± 1,87 c |
| apoCI1 (6.430) | 7,24 ±3,09 | 4,30 ± 2,79 a |
| apoCI2 (6.630) | 11,33 ±5,66 | 6,95 ± 4,64 a |
| ApoAI (28.130) | 1,22 ± 1,29 | 0,57 ± 0,58 a |
| ApoE (33.570) | 1,49 ± 0,64 | 1,13 ±0,53 b |
Los valores representan microgramos de las apolipoproteínas individuales por miligramo de apoB100 en la fracción de LDL±desviación estándar.
a p<0,05; b p<0,01; c p≤0,001 (significación estadística evaluada con el test de Student).
Modificada de Davidsson et al.17.
En diciembre del 2011 Sacks et al. publicaron los resultados de un estudio prospectivo que documenta sólidamente la asociación de niveles altos de apoCIII en las LDL con el aumento del riesgo coronario por enfermedad ateroesclerótica20. Estos investigadores siguieron durante 10-14 años a 640 mujeres y a 838 hombres, libres de enfermedad coronaria al inicio del estudio. Al final del estudio se seleccionaron 320 mujeres y 419 hombres que desarrollaron infartos del miocardio fatales y no fatales. Este grupo se comparó con igual número de controles apareados libres de infartos al fin del estudio. Utilizando un análisis multifactorial se encontró que el quintil de los pacientes con más alto nivel de apoCIII en las LDL experimentó un significativo aumento del riesgo de 2,38 (1,54-3,68; p<0,001) respecto al quintil con más baja apoCIII en la LDL. Tal aumento mostró ser independiente del nivel de TG y del cociente de TG/HDL-c. Los datos indican que la aterogenidad de la apoCIII es independiente de la dislipidemia de la IR y de otras causas de hipertrigliceridemia e hipercolesterolemia20.
Estudios genéticos recientes también dan apoyo a la hipótesis de la aterogenicidad intrínseca de la apoCIII. Pollin et al.21 encontraron que una mutación asociada con disminución de la apoCIII en humanos se asocia a niveles de TG muy bajos. Esta mutación tiene una prevalencia alta (5%) en una comunidad Amish de Lancaster (Pensilvania). Análisis de dicha población mostraron además que los sujetos con la mutación nula del gen de la apoCIII presentan niveles de calcificación de las arterias coronarias menores que los presentes en sujetos de la misma comunidad sin la mutación, lo cual se considera un indicio de resistencia al desarrollo de ateroesclerosis21.
Función fisiológica de la apoCIIIEn el plasma, la apoCIII se encuentra unida a las VLDL, a las LDL, a las HDL y a los quilomicrones. Además, una fracción pequeña parece existir no ligada a lipoproteínas. No obstante, esto puede ser un artefacto de los métodos usados para separar las partículas lipoproteicas del resto de las proteínas plasmáticas utilizando ultracentrifugación con soluciones con altas concentraciones de sales neutras (KBr), las cuales pueden despegar apolipoproteínas de las partículas lipoproteicas22. El efecto inhibidor de la apoCIII sobre la hidrólisis de los TG de las VLDL y de los quilomicrones se confirmó con estudios en sujetos con deficiencia en la expresión del gen de la apoCIII. Estas personas en ayunas muestran niveles de TG de las VLDL muy bajos2. Más pruebas del papel de la apoCIII como inhibidora de la LPL se obtuvieron con ratones transgénicos con el gen de la apoCIII humana y con sobreexpresión de la apolipoproteína. Tales animales muestran un aumento importante de las VLDL y de sus TG, mientras que ratones con disrupción del gen de la apoCIII endógena tienen niveles muy bajos de TG circulantes. Estos datos confirman que la apolipoproteína es un inhibidor fisiológico de la actividad de la LPL2,23,24. Aún no se conoce en detalle cómo la apoCIII inhibe a la LPL. Se ha propuesto que el bloqueo de la hidrólisis de los TG de lipoproteínas ricas en TG (VLDL y quilomicrones), así como de la hidrólisis de emulsiones sintéticas contentivas de TG, se debe a un desplazamiento de otra apolipoproteína, la apoCII, de la superficie de partículas ricas en TG. La apoCII, al contrario que la apoCIII, es un activador de la hidrólisis de los TG de las VLDL y de los quilomicrones mediada por la LPL. La apoCII posiblemente activa a la LPL al mejorar el contacto entre la enzima (LPL) y los TG en la superficie de las lipoproteínas. Dicho efecto activador se pierde cuando la apoCII es desplazada por la apoCIII2,5. La apoCI y la apoE también favorecen la asociación de la LPL con las VLDL y las LDL, y es posible que la apoCIII también las desplace. Nuestros datos, resumidos en la tabla 2, apuntan a esta posibilidad, ya que las LDL de los pacientes con alto contenido de apoCIII tienen menor contenido de apoCI y de apoE17.
Bases bioquímicas de las propiedades de la apoCIII y otras apolipoproteínasAntes de la discusión de los posibles mecanismos de la aterogenicidad de la apoCIII, es útil resumir algunos aspectos básicos de las bases moleculares de las funciones de las apolipoproteínas. Como todas las apolipoproteínas, la apoCIII reside en la superficie de las VLDL y de otras lipoproteínas. La propiedad de adherirse a las partículas por interacciones no covalentes se debe a que las apolipoproteínas reconocen, en la microemulsión esférica constituida por los lípidos, la estructura de la superficie donde residen los grupos polares de estos lípidos. Dicha superficie es la región de contacto con el medio acuoso. La mayoría de las apolipoproteínas tienen en sus cadenas peptídicas regiones ricas en aminoácidos polares alternadas con regiones ricas en aminoácidos no polares. Estos segmentos peptídicos, al ponerse en contacto con microemulsiones lipídicas, se orientan en estructuras helicoidales de 18 a 25 aminoácidos de longitud con una cara hidrofóbica adherida a los lípidos y una cara polar en contacto con el medio exterior acuoso. Las estructuras reciben el nombre de hélices anfipáticas (del griego anfi, «ambas», y pathos, «afinidad»: una cara polar y una cara no polar), que han sido caracterizadas en gran detalle por Segrest, Phllips y Lund-Katz25,26. En la microemulsión lipídica constituyente de las lipoproteínas las cadenas hidrofóbicas de los ácidos grasos esterificados al glicerol en los TG y fosfolípidos, así como los esterificados al colesterol, se orientan hacia el interior de la microemulsión. Por su parte, las apolipoproteínas se extienden sobre la superficie de las microemulsiones en contacto con los grupos polares de los lípidos que miran a la superficie; estas regiones de las apolipoproteínas son ricas en aminoácidos hidrofóbicos (alanina, leucina, isoleucina, tirosina y triptófano). Hacia el medio exterior acuoso se orienta la otra cara hecha de segmentos ricos en aminoácidos polares con cadenas hidroxiladas (serna treonina) o con cargas negativas (aspártico, glutámico) o cargas positivas (lisina y arginina)25,26.
La asociación de las apolipoproteínas CI, CII y CIII y de las apolipoproteínas AI, AII y E con las partículas es reversible y pueden intercambiarse entre las diferentes lipoproteínas. Este no es el caso de la apoB-100 ni de la apoB-48, las cuales, debido a sus largas cadenas de más de 4.000 y 2.000 aminoácidos, respectivamente, y a la presencia de numerosas regiones hidrofóbicas, permanecen unidas a las microemulsiones lipídicas y son parte permanente de las VLDL, de las LDL y de los quilomicrones. Más aún, la apoB-100 y la apoB-48 se mantienen unidas a las partículas durante la conversión metabólica de las VLDL en LDL y durante la conversión de los quilomicrones a remanentes de quilomicrones. Esto no ocurre con las apolipoproteínas apo-CI, apo-CII, apo-CIII, apo-E, apo-AI y apo-AII, las cuales se redistribuyen entre los quilomicrones VLDL, LDL y HDL durante la remodelación continua de estas partículas que tiene lugar en la circulación. Esta propiedad ha llevado a estas últimas apolipoproteínas a ser designadas como «intercambiables»25,26. Los principales agentes de la remodelación intravascular de las lipoproteínas son enzimas que hidrolizan a los TG y a los fosfolípidos de las lipoproteínas, tales como la LPL, la lipasa hepática (HL) y diversas fosfolipasas presentes en el plasma y asociadas a la superficie endotelial. Además, otras proteínas que redistribuyen lípidos, como la proteína transportadora de ésteres de colesterol (CETP) y la proteína transportadora de fosfolípidos (PLTP), contribuyen a la remodelación y a la redistribución de las apolipoproteínas intercambiables.
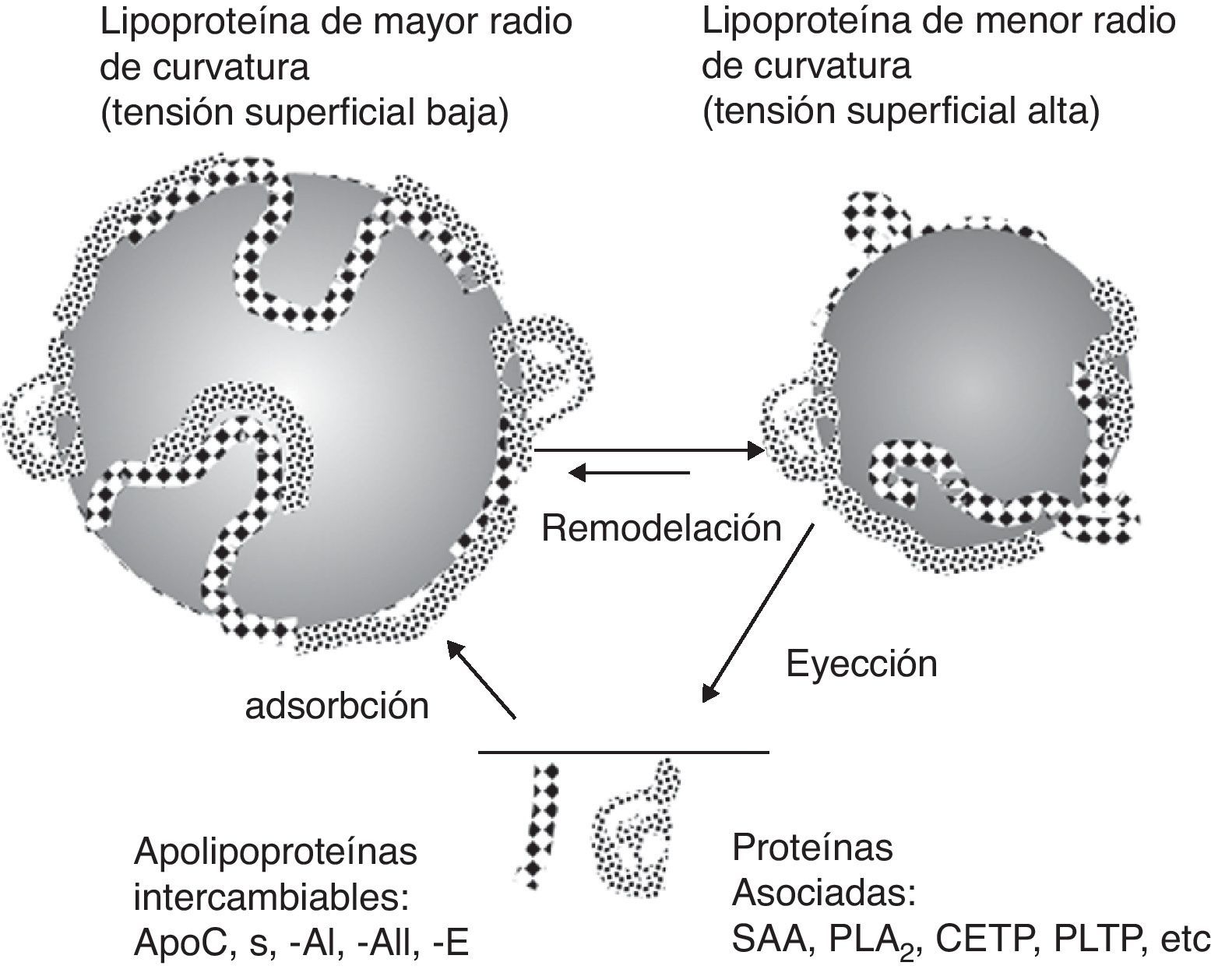
La afinidad de las apolipoproteínas intercambiables por la superficie de las lipoproteínas disminuye cuando el radio de curvatura de las partículas aumenta al decrecer su volumen, lo cual lleva a un aumento de la presión lateral (interfacial) de la capa monomolecular que forma la superficie de las partículas, provocando la separación de las apolipoproteínas intercambiables y de otras proteínas asociadas, permitiendo así el intercambio entre lipoproteínas o el intercambio con membranas de células endoteliales y de otros tejidos27. Es decir, la distribución de la apoCIII y de otras apolipoproteínas entre las diversas lipoproteínas es un fenómeno dinámico modulado constantemente por las modificaciones a las que están sometidas todas las lipoproteínas durante su secreción, circulación y metabolismo en el plasma y en la linfa. Además, a las lipoproteínas se adhieren diversas proteínas que no se consideran apolipoproteínas, tales como las fosfolipasas secretoras, la proteína amiloide del suero y múltiples componentes del sistema immunológico27,28. La figura 1 presenta un diagrama de los fenómenos tratados28.
")
Diagrama de cambios en el contenido de apolipoproteínas y de proteínas asociadas a lipoproteínas. Los cambios son causados por la remodelación de lipoproteínas y por la alteración de la tensión interfacial y de la estructura superficial al cambiar de volumen las partículas. Las flechas dobles indican que los procesos son reversibles. (Modificado de Davidsson et al.28.)
Las células endoteliales que separan la circulación sanguínea y linfática del parénquima de órganos y tejidos en general están cubiertas en su superficie por una alfombra de proteoglicanos y proteínas que constituyen el glicocálix29. Es en este ambiente donde tiene lugar la hidrólisis de los TG de los quilomicrones y de las VLDL por la LPL en la superficie de células endoteliales que cubren al tejido adiposo, al músculo cardiaco, al espacio de Disse en los hepatocitos y a las células del músculo esquelético. En esta superficie, las lipoproteínas ricas en TG y contentivas de apolipoproteínas B-100, B-48 y apoE son inmovilizadas transitoriamente al asociarse sus regiones ricas en aminoácidos con carga positiva (lisina y arginina) con proteoglicanos ricos en glucosaminoglicanos sulfatados, heparán sulfato y condroitín sulfatos. Los glucosaminoglicanos sulfatados del glicocálix confieren a tal superficie del endotelio una carga negativa, y por ello retiene a las lipoproteínas contentivas de apoB y apoE. Además retiene varias enzimas lipolíticas, como la LPL, la HL y varias fosfolipasas29,30. Es decir, el glicocálix crea un medio en donde se retiene a las lipoproteínas ricas en TG, quilomicrones, VLDL y a sus respectivos remanentes, así como a las enzimas que actúan sobre ellas. Se ha propuesto que la apoCIII inhibe a la acción de la LPL sobre las VLDL y los quilomicrones porque desplaza a la apoCII y a la apoE de la superficie de las partículas, proteínas estas últimas que son activadoras de la LPL. La primera se requiere para la unión de la enzima con su sustrato, los TG, y la apoE contribuye a la inmovilización de las lipoproteínas por los glucosaminoglicanos sulfatados en el glicocálix al incrementar las cargas positivas en la superficie de ellas. La apoCIII también inhibe a la LPL porque en la superficie de las VLDL y de los quilomicrones se orienta y expone sus segmentos cargados negativamente ricos en aspártico y glutámico hacia el medio acuoso, lo cual resulta en una disminución de la carga positiva de la superficie de las lipoproteínas. La carga positiva de las lipoproteínas, como se ha mencionado, es requerida para la inmovilización de las partículas por los glucosaminoglicanos de los proteoglicanos del glicocálix5,30. Es decir, un aumento de las apoCIII acompañado de la disminución relativa de las apoCII y apoE en las lipoproteínas ricas en TG disminuirá su fijación al glicocálix del endotelio, limitando el tiempo para la acción de la LPL sobre las partículas y prolongando así su vida media en la circulación. Un fenómeno análogo al descrito de reconocimiento entre las cargas positivas en la superficie de los remanentes de las VLDL y de los quilomicrones con cargas negativas en el glicocálix de los hepatocitos se requiere para que el hígado capture a estas partículas mediante una acción concertada con receptores apoB/E en las membranas plasmáticas de las células. Este proceso se dificulta cuando un exceso de apoCIII acompaña a los remanentes de VLDL y de quilomicrones30.
Mecanismos contribuyentes a la aterogenicidad de la apoCIII asociada a las LDL y las VLDLRetención de LDL rica en apoCIII por la íntima arterialLos estudios clínicos discutidos indican que un aumento de la apoCIII asociada a las LDL contribuye al aumento del riesgo cardiovascular. Para discutir las posibles razones de este incremento en la aterogenicidad es útil usar como guía la llamada «hipótesis de la respuesta a la retención»31-35. Tal hipótesis, extensamente documentada por estudios in vivo e in vitro, propone que el desarrollo de lesiones ateroescleróticas tiene como paso inicial la captura de lipoproteínas contentivas de la apoB-100, LDL y posiblemente remanentes de VLDL, en la íntima arterial. Una vez depositadas en la íntima, las lipoproteínas son modificadas por procesos enzimáticos y oxidativos, los cuales generan potentes agentes inflamatorios como lisofosfolípidos, colesterol libre, ácidos grasos y fosfolípidos oxidados, así como fragmentos productos de la proteólisis de la apoB-100 con propiedades antigénicas. Se cree que dichos productos perpetúan un ciclo de deposición de lipoproteínas apoB-100 y generación de productos inflamatorios. La captura de las LDL tiene lugar por la formación de complejos insolubles con proteoglicanos de la íntima, principalmente con el versican y el biglican35. Y es en este paso inicial de la aterogénesis donde un aumento de la apoCIII en la LDL puede incrementar la expansión de lesiones. Experimentos de Chait et al. muestran que la afinidad de la LDL y de la VLDL por los proteoglicanos arteriales se incrementa con el contenido de apoCIII en las lipoproteínas34,36,37. Nuestro laboratorio, por otra parte, ha mostrado que las LDL, cuando son pequeñas y densas (LDLpd), tienen una mayor afinidad por los proteoglicanos arteriales que las LDL más grandes17. Las LDLpd son especialmente ricas en apoCIII y son abundantes en pacientes con síndrome metabólico y diabetes, y se cree que son una de las causas del aumento del riesgo cardiovascular asociado con estos trastornos metabólicos3,25. Nuestros datos han mostrado que aumentando in vitro el contenido de apoCIII en las LDL del plasma se aumenta proporcionalmente su afinidad por los proteoglicanos arteriales17 Más aún, utilizando análisis proteómico, encontramos que las LDLpd de sujetos con IR y ateroesclerosis periférica tienen un contenido más alto de apoCIII y una mayor afinidad por los proteoglicanos arteriales que las LDLpd de controles sanos, lo cual extiende las observaciones de Chait et al. a experimentos ex vivo17. Además, tales datos indican que es el contenido de apoCIII y no el tamaño de las LDL lo que aumenta la afinidad por los proteoglicanos. En la tabla 2 se presentan los resultados de la evaluación con espectroscopia de masas de las principales apolipoproteínas identificadas en las LDLpd. Esta metodología permitió medidas de las diferentes isoformas de la apoCIII (apoCIII0, apoCIII1 y apoCIII2), y encontramos que todas ellas estaban elevadas en las LDL de los pacientes en comparación con los sujetos controles. La tabla 2 presenta asimismo las medidas del contenido de apoCI, apoCII, apoAI y apoE, que son más altos en las LDLpd de los controles que en la fracción análoga de los pacientes. Basándonos en estos datos y en los publicados por Chait et al.36 hemos sugerido que en los pacientes estas últimas apolipoproteínas intercambiables en las LDLpd han sido desplazadas de la superficie de las partículas por el incremento de las apoCIII17. Debemos señalar que nuestros resultados tienen la limitación de que el número de sujetos estudiado es limitado. Esto se debe a que aunque los análisis proteómicos tienen sensibilidad y precisión, son todavía laboriosos y por ello solo aplicables hasta el momento a grupos pequeños de muestras. No obstante, resultados recientes usando grupos más numerosos de sujetos y con ayuda de métodos inmunológicos han confirmado la presencia de LDLpd ricas en apoCIII y con alta afinidad por los proteoglicanos en pacientes con diabetes tipo 2 pero no en controles sanos37.
No conocemos en detalle cómo un aumento de la apoCIII en las LDL aumenta su afinidad por los proteoglicanos, pero resultados obtenidos en distintos laboratorios permiten especular que al orientarse la apoCIII sobre la superficie de las LDL, expone regiones ricas en aminoácidos cargados positivamente (lisina y arginina) hacia la superficie, aumentando así la carga positiva neta de la LDL y, por lo tanto, su afinidad por las cargas negativas de los glucosaminoglicanos (grupos sulfato y carboxilo) de los proteoglicanos5,17,36. Si esto ocurre en la dislipidemia de la IR con un aumento de la LDLpd rica en apoCIII, esta partícula pudiera ser más aterógena por tener una mayor afinidad por los proteoglicanos arteriales, lo cual podría resultar en una mayor tendencia a ser depositada en la íntima arterial, facilitando allí las modificaciones oxidativas y enzimáticas generadoras de la respuesta inflamatoria disparada por la LDL.
La apoCIII por sí misma contribuye a una respuesta inflamatoria aterógenaExiste consenso en que la respuesta inflamatoria a la deposición de lipoproteínas apoB-100 en la íntima es requerida para el desarrollo de placas ateroescleróticas32,34,35,38. La apoCIII parece estar directamente implicada en estos mecanismos aterógenos. La adhesión de monocitos al endotelio, el paso hacia la íntima y su conversión a macrófagos residentes precede a su eventual evolución a células espumosas causada por la captación no regulada de lipoproteínas apoB depositadas en la íntima38. Akio Kawakami, junto con Frank Sacks y Peter Libby en Boston, mostraron que la apoCIII por sí sola o unida a la VLDL es capaz de estimular la expresión de proteínas de adhesión celular (VCAM, ICAM) en el endotelio, las cuales son requeridas para la fijación de monocitos39. La apoCIII estimula la secreción de las proteínas de adhesión molecular activando una cascada fosforiladora iniciada por la proteína-cinasa C-beta, y esta a su vez activa el factor nuclear NF-kapa-B. El resultado de dicha cascada activadora es la adhesión de monocitos al endotelio y su eventual penetración en la íntima40. Los datos inicialmente reportados en cultivos de células endoteliales y usando monocitos aislados han sido confirmados in vivo usando ratones41,42. Kawakami et al. mostraron además que la apoCIII, por sí misma o unida a las VLDL, induce en los monocitos circulantes las integrinas requeridas para la interacción con las VCAM e ICAM de la superficie del endotelio. In vivo la apoCIII estimula la respuesta de adhesión también vía activación de la proteína-cinasa C-alfa con el subsiguiente estímulo del factor nuclear NF-kapa-B. Kawakami et al. mostraron también que la apoCIII por sí misma o unida a lipoproteínas inhibe la activación por la insulina del receptor sustrato de la insulina (IRS-1) a través de la estimulación de la isoforma PKCβ de la proteína-cinasa C. Tal efecto en células endoteliales en cultivo y en ratones se acompaña de una disminución de la producción de óxido nítrico (NO)41. No está claro si los efectos de la apoCIII sobre los monocitos y las células endoteliales se inician con una interacción con un receptor específico. Kawakami y Yoshida han sugerido que pueden estar implicados receptores acoplados a proteínas G (GCPR) o los receptores que reconocen a la apoB y apoE en lipoproteínas40-43. Los mecanismos moleculares de los efectos descritos todavía no están claros, pero estos importantes experimentos confirman que la apoCIII estimula una respuesta aterógena en los dos tipos de células envueltas en la eventual acreción de macrófagos en sitios de progresión de lesiones42.
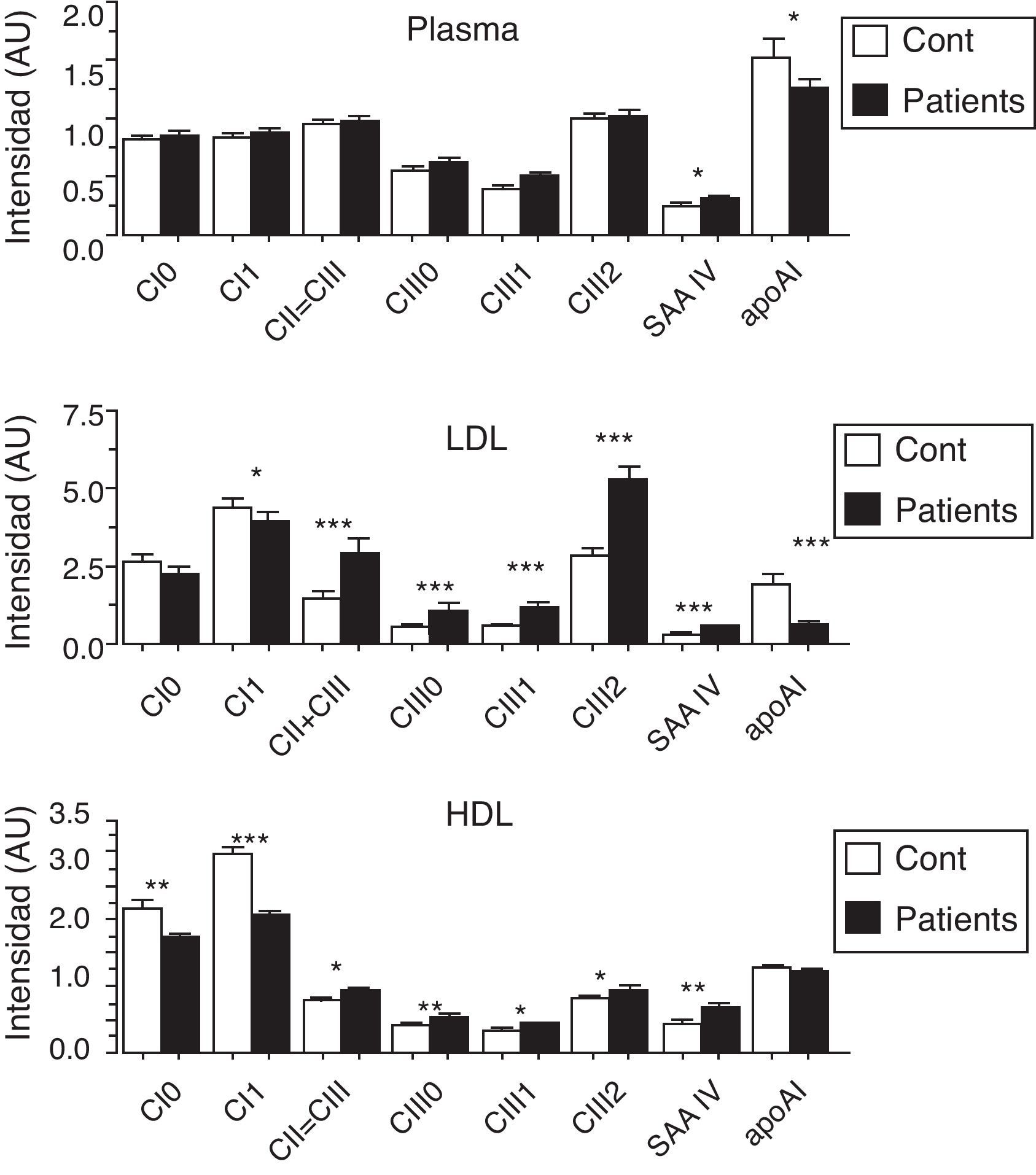
La apoCIII está presente en distintas cantidades en todas las lipoproteínas, incluyendo las HDL. Múltiples estudios indican que niveles elevados de HDL, especialmente las partículas grandes, designadas HDL2 o HDL-α, disminuyen el riesgo cardiovascular44. Diversas líneas de investigación sugieren que estas propiedades anti-aterógenas se deben a efectos antiinflamatorios y antioxidantes45. Sin embargo, existen alteraciones estructurales, relacionadas con cambios en el contenido de apolipoproteínas y de otras proteínas asociadas de las HDL que pueden comprometer sus acciones protectoras. Las HDL, además de sus principales apolipoproteínas (apoAI, apoAII y apoE), tienen asociadas pequeñas cantidades de las apoCI, CII y CIII y de proteínas del sistema inmune28,45. En experimentos in vitro la HDL ha mostrado que inhibe la adhesión de leucocitos a células endoteliales mediados por las VCAM y las integrinas, lo cual se ha postulado es uno de sus efectos protectores45. Recientemente, Kawakami et al.42 demostraron que cuando la HDL se enriquece en apoCIII, su efecto de inhibir la adhesión de leucocitos al endotelio se disminuye significativamente, lo cual, según los autores, podría comprometer una de sus principales propiedades antiinflamatorias HDL in vivo42. En relación con el contenido de apoCIII en la HDL, nuestro laboratorio reportó resultados de un análisis proteómico de las apolipoproteínas de las LDL y HDL y del plasma total en pacientes con diabetes tipo 2, IR y ateroesclerosis periférica, así como en controles sanos apareados por sexo y edad28. La figura 2, tomada de esta publicación, muestra el contenido mayor de apoCIII en la HDL de los pacientes respecto a los controles. Estos hallazgos, y los citados de Kawakami et al.42, sugieren que el aumento de apoCIII en HDL observado en la dislipidemia de la IR podría comprometer las propiedades antiinflamatorias de la lipoproteína y, junto con un aumento de la acumulación de la LDLpd en la íntima, contribuir a la aterogenicidad de esta dislipidemia.
")
Apolipoproteínas intercambiables y otras proteínas en las lipoproteínas de pacientes con ateroesclerosis periférica y diabetes y de controles sanos apareados por sexo y edad. Las proteínas de menos de 30kDa fueron evaluadas con espectrometría de masas usando un análisis proteómico. Los datos muestran que los niveles de todas las isoformas de apoCIII son más altos en los pacientes que en los controles. Nótese que las diferencias observadas son mucho más significativas cuando los análisis se hacen en las lipoproteínas aisladas una vez separadas. (Modificado de Davidsson et al.28.)
Fármacos activadores de los receptores nucleares PPARα son efectivos en la disminución de la concentración de la apoCIII en circulación. Los fibratos en uso clínico disminuyen la secreción hepática de las VLDL, lo cual resulta en una reducción de la apoCIII unida a estas partículas y a otras lipoproteínas. Además el pioglitazone, un fármaco con efectos en los receptores PPARγ y PPARα, es especialmente eficaz en reducir las VLDL y, como consecuencia, la apoCIII. Es muy posible que esto contribuya a sus efectos de reducción del progreso de placas ateroescleróticas documentados recientemente en pacientes diabéticos utilizando ultrasonidos intravasculares46. Varias sustancias que actúan sobre diferentes PPAR(s) están en fase de desarrollo clínico, pero datos preliminares presentados en congresos indican que reducen ya efectivamente los niveles de apoCIII. Es importante señalar que el gen de la apoCIII está bajo el control parcial de un elemento regulador que responde a la insulina en el hígado, lo cual indica que cualquier aproximación terapéutica que mejore la acción de la insulina en el hígado, incluyendo la actividad física y el control de la ingesta calórica, debe resultar en una disminución de la apoCIII que acompaña a la secreción de las VLDL. Por último, agentes específicos por la expresión del gen de la apoCIII (antisense) también están en desarrollo clínico y, basándose en resultados en animales, se espera que sean muy eficaces.
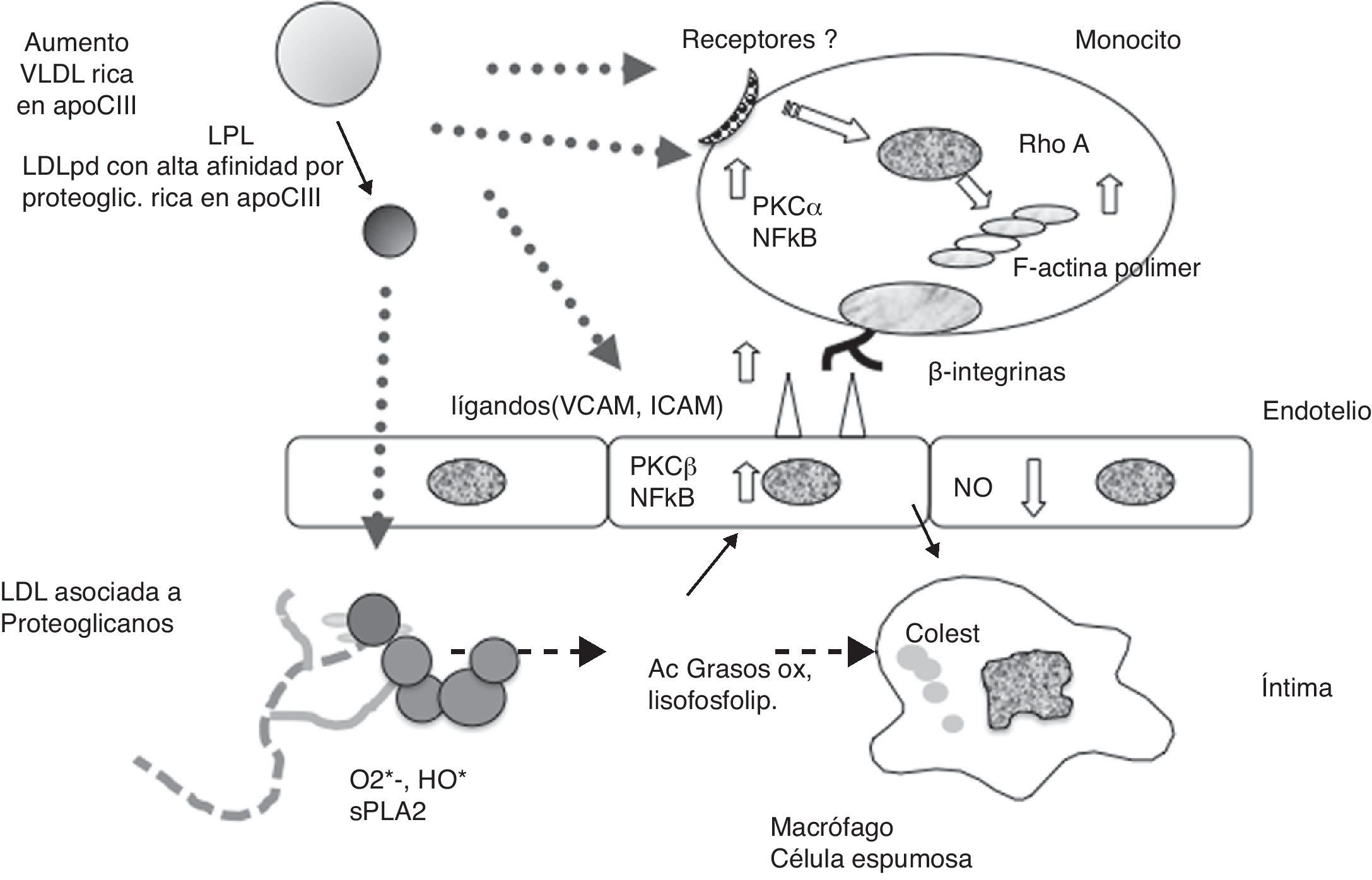
ConclusionesLos resultados discutidos sugieren que las dislipidemias acompañadas de un aumento de la apoCIII en las VLDL LDL y HDL, como la inducida por la IR y la diabetes, son agentes generadores de los dos aspectos que la hipótesis de la «respuesta inflamatoria a la retención de LDL» implica como aterógenos. Pero, además, trabajos clínicos muestran que la LDL rica en apoCIII es un factor de riesgo cardiovascular asociado o no a hipertrigliceridemia o a la dislipidemia de la IR. Experimentos que hemos discutido indican que, por una parte, la LDL contentiva de niveles altos de la apoCIII tiene una afinidad elevada por los proteoglicanos de la matriz extracelular de la íntima, lo cual podría ser la principal causa de la retención preferencial de estas partículas en regiones propensas a desarrollar lesiones ateroescleróticas. Además, las lipoproteínas contentivas de apoB-100 (VLDL; remanentes de VLDL y LDL) ricas en apoCIII parecen disparar la cascada inflamatoria considerada como la respuesta de la pared arterial a la retención de dichas partículas. Estos dos mecanismos podrían explicar el papel de factor de riesgo cardiovascular independiente adscrito a la apoCIII. La figura 3 presenta un diagrama en el que se resumen los dos mecanismos aterógenos de las lipoproteínas apoB100 ricas en apoCIII.
, tienen una alta afinidad por los proteoglicanos de la íntima arterial, y ello contribuye a su retención. Tanto las VLDL como LDL ricas en apoCIII activan los mecanismos de la respuesta inflamatoria de leucocitos en circulación y del endotelio, incrementando la expresión de moléculas de adhesión celular. La apo CIII posiblemente utiliza receptores acoplados a proteínas G y/o los receptores en monocitos y endotelio que reconocen la apoB y apoE en las lipoproteínas. Esto ocasiona la penetración de monocitos hacia la íntima, su conversión a macrófagos y eventualmente a células espumosas. En el endotelio, la apoCIII compromete la producción de óxido nítrico (NO) (De Kawakami et al.39-43).")
Posible origen de la aterogenicidad de las lipoproteínas contentivas de la apoB-100 y ricas en apoCIII dentro de los conceptos de la hipótesis de la ateroesclerosis «respuesta a la retención». Las LDL ricas en apoCIII, especialmente las pequeñas y densas (LDLpd), tienen una alta afinidad por los proteoglicanos de la íntima arterial, y ello contribuye a su retención. Tanto las VLDL como LDL ricas en apoCIII activan los mecanismos de la respuesta inflamatoria de leucocitos en circulación y del endotelio, incrementando la expresión de moléculas de adhesión celular. La apo CIII posiblemente utiliza receptores acoplados a proteínas G y/o los receptores en monocitos y endotelio que reconocen la apoB y apoE en las lipoproteínas. Esto ocasiona la penetración de monocitos hacia la íntima, su conversión a macrófagos y eventualmente a células espumosas. En el endotelio, la apoCIII compromete la producción de óxido nítrico (NO) (De Kawakami et al.39-43).
Los autores consideran que el contenido de esta revisión está libre de conflicto de intereses.