





: The aim of the present study was to evaluate the effects of ovariectomy on the secretory apparatus of natriuretic peptides in right atrial cardiomyocytes.
METHODS:Nine-month-old mice underwent bilateral ovariectomy or sham surgery. The blood exam of the ovariectomized mice showed results consistent with castrated females. Systolic blood pressure was measured after ovariectomy (9 mo of age) and at the moment of sacrifice (12 mo of age). Fragments of the right atrium were collected and prepared for electron microscopy examination. The following variables were quantified: the quantitative density and area of the natriuretic peptide granules, the relative volume of euchromatin in the nucleus, the number of pores per 10 μm of the nuclear membrane and the relative volumes of the mitochondria and Golgi complex.
RESULTS:The cardiomyocytes obtained from ovariectomized mice indicated that the quantitative density and the area of secretory granules of natriuretic peptides were significantly lower compared with the sham-operated mice. Furthermore, there was a decrease in the relative volume of euchromatin, a lower density of nuclear pores, and lower relative volumes of the mitochondria and Golgi complex in the ovariectomized mice compared with the sham-operated mice. These findings suggest a pool with a low turnover rate, i.e., low synthesis and elimination of natriuretic peptides.
CONCLUSION:A lack of estrogen caused hypotrophy of the secretory apparatus in right atrial cardiomyocytes that could explain the weak synthesis of natriuretic peptides in mice. Furthermore, one of the mechanisms of blood pressure control was lost, which may explain, in part, the elevated blood pressure in ovariectomized mice.
Natriuretic peptides (NPs) are synthesized, stored and secreted by the cardiac atria and play an important role in blood volume homeostasis and in the normal regulation of arterial blood pressure (1-2). Ultrastructural histochemical studies have shown that specific atrial granules contain NP prohormones (3). The production and secretion of NPs depend on the structural components of the cardiomyocytes, including the number of pores in the nuclear membrane, the euchromatin, the Golgi complex and the mitochondria, which have collectively been termed the secretory apparatus of the cardiomyocyte (4). NPs are primarily released in response to stretching of the atria wall, which occurs during the elevation of blood pressure (1).
The influence of estrogens on the production and secretion of NPs has been well established in humans (5), rats (6) and mice (7). Estrogens act by stimulating the production and secretion of NPs; the levels of estrogen are low in ovariectomized (OVX) rats and mice (8) but are restored by 17β-estradiol administration (9-16). However, to the best of our knowledge, no studies in the literature have examined the relationship between estrogen, arterial pressure and the secretory apparatus of NPs in hypertensive mice.
The purpose of the present study was to extend previous findings in two important ways. First, the effects of estrogen deprivation on blood pressure were determined. The number and size of NP granules were assessed by a comparison of electron photomicrographs of right atrial (RA) cardiomyocytes obtained from two sets of 12-mo-old animals, i.e., sham-operated and OVX mice.
Second, a quantitative evaluation of the secretory apparatus of the RA cardiomyocytes obtained from the OVX and sham-operated mice was performed: the number of pores per 10 μm of nuclear membrane and the relative volumes (%) of the euchromatin, mitochondria and Golgi complex in the cardiomyocytes obtained from the two groups were estimated and compared.
METHODSAnimalsFourteen female virgin C57BL mice (9 mo of age) were obtained from the Animal Care Facility of São Judas Tadeu University (São Paulo, Brazil). The animals were housed in individual cages in a temperature-controlled room (22°C) with a 12-hour dark/light cycle.
Experimental designAt the end of the 9th month, the mice were randomly assigned to the sham-operated (Sham, n = 7) or OVX (n = 7) groups. The animals in the OVX group underwent a bilateral ovariectomy under anesthesia, which was induced by an intraperitoneal injection of ketamine (75 mg/kg) and diazepam (5 mg/kg) (17). Following the surgery, the animals received an injection of antibiotics (40,000 U/kg penicillin G procaine IM).
Three months after the ovariectomy (i.e., 12 months of age), the animals from both groups were euthanized with an intraperitoneal injection overdose of sodium pentobarbital. Approximately 1 ml of blood was collected from each mouse during the euthanasia procedure.
Determination of blood estradiol levels and blood pressure measurementsBlood levels of 17β-estradiol were determined by a competitive enzyme immunoassay in a specialized laboratory. All samples were measured in the same assay. Systolic blood pressure was measured at two time points: after ovariectomy (9 mo of age) and at the moment of sacrifice (12 mo of age). The mice were anesthetized (ketamine-xylazine 80:40 mg/kg ip) and the blood pressure was evaluated by indirect measurement using the tail-cuff method (18).
Determination of the number and size of NP granules and the relative volumes of the mitochondria and Golgi complexFragments of the right atrium were fixed in 5% glutaraldehyde in a phosphate buffer solution (0.2 M, pH 7.3) for three hours. The samples were then washed with the same buffer solution for 15 minutes and placed in a solution of osmium tetroxide in 1% phosphate buffer for 2 hours. The fragments remained in 0.5% uranyl acetate for 12 hours and were dehydrated in an ascending series of alcohols and propylene oxide for 8 hours under rotation. The fragments were included in pure resin for 5 hours and maintained on the same resin at 60°C for 5 days.
Semi-thin sections were used to locate longitudinally oriented cardiomyocytes with a central nucleus and to obtain ultra-thin sections using an ultra-microtome (Sorvall MT-2). The ultra-thin sections were contrasted with uranyl acetate and lead citrate and examined under a transmission electron microscope (Jeol, ICB, São Paulo University). Five sections per animal were photographed with a final magnification of x7,500 to count the NP granules (number of granules per cardiomyocyte) (19-24). Five electron micrographs per animal, which were examined with a final magnification of x15,000, were used to determine the relative volumes of the mitochondria and Golgi complex and to estimate the size (section area) of the secretory granules.
Determination of the number of pores in the nuclear membrane and the relative volume of euchromatinFive electron micrographs per animal, which were examined with a final magnification of x15,000, were used to obtain the number of pores per 10 μm of nuclear membrane and the relative volume (%) of euchromatin in the cardiomyocyte nucleus.
The relative volume corresponds to the area occupied by the structure presented as a %. The relative volume was obtained using a test system equipped with 81 points (considered as 100%), which were allocated on each electronic image where the points for each component were counted.
The following formula was used to calculate the relative volumesVv[struc] = ΣP[struct] .100/PT; where Vv[struct] = Relative volume, ΣP[struct] = Number of points on the structure in question, and PT = Total number of points (81) of the test system (25-26). All measurements were performed using an image analysis program (Axio Vision, Zeiss, 2009) obtained from the Laboratory of Quantitative Studies of São Judas Tadeu University, São Paulo.
Statistical analysisThe data obtained for each parameter were tabulated and the means were calculated. Significant differences between the means were evaluated using Student's t test. P<0.05 was considered significant. The GraphPad Prism 5 program (GraphPad Software, San Diego, CA, USA) for Windows was used for data analysis.
Ethical informationAll surgical procedures and protocols outlined in this study were approved by the Experimental Animal Use Committee of São Judas University and were conducted in accordance with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (Ethical Protocol number A003/2010).
RESULTSEstradiol plasma levels and systolic blood pressureThe analysis of blood obtained from the OVX mice indicated results that were consistent with castrated females: the plasma estradiol levels were significantly lower compared with the sham-operated mice (p<0.05). The mean systolic blood pressure at rest was significantly higher in the OVX compared with the sham-operated mice at the time of sacrifice (p<0.05) (Table1).
Plasma estradiol levels and systolic blood pressure (SBP) in the two studied groups of mice.
| Parameters | Sham-operated | OVX | p-value |
|---|---|---|---|
| Estradiol (pg/ml) | 95±3.2 | 32±0.4* | <0.05 |
| SBP (mm Hg) | 112±3.0 | 127.3±2.0* | <0.05 |
Values represent the means ± SD. *p<0.05 vs. sham group. Student's t test was used for the statistical analysis.
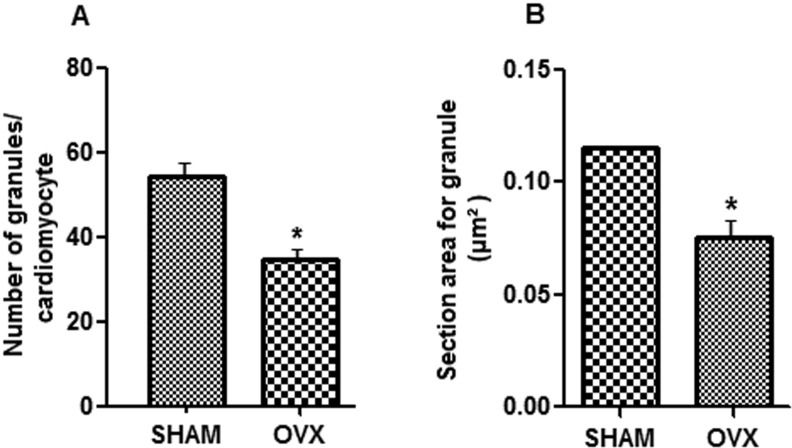
Cytoplasmic secretory granules were predominantly arranged in groups among the mitochondria (Figure1A) and near the Golgi complex (Figure1B). The number of NP granules in the cytoplasm of the cardiomyocyte was lower in the OVX mice compared with the sham-operated mice (p<0.05). Ovariectomy also significantly reduced the size of the NP granules (p<0.05) (Figure2).
 the number of NP granules (white arrows) dispersed among mitochondria (M) and (B) the mitochondria with varied forms (M) and NP granules near the Golgi complex (G).")
 and size (B) of NP granules in RA cardiomyocytes. *p<0.05 vs. sham. Values represent the means ± SD. Student")
The Golgi complex exhibited widened cisternae predominantly localized among the cytoplasmic secretory granules; the mitochondria appeared as a heterogeneous mixture of elongated, tubular forms and short vesicular forms (Figure1). The OVX mice exhibited lower relative volumes of the Golgi complex and mitochondria (%) compared with the sham group (p<0.05) (Table2).
- Morphometric indexes of the cardiomyocyte cytoplasm and nucleus in the RA of sham and OVX mice.
| Morphometric indexes | Sham-operated | OVX | p-value |
|---|---|---|---|
| RVGC (%) | 2.2±0.2 | 1.7±0.2* | <0.05 |
| RVM (%) | 23±3.0 | 19±1.0* | <0.05 |
| RVE (%) | 66±5.0 | 58±2* | <0.05 |
| NPs/10 μm NM | 4.1±0.3 | 3.0±0.1* | <0.05 |
Values represent the means ± SD. RVGC, relative volume of Golgi complex; RVM, relative volume of mitochondria; RVE, relative volume of euchromatin; NP/10 μm NM, number of pores/10 μm of nuclear membrane. *Significant vs. sham group. Student's t test was used for the statistical analysis.
In the cardiomyocytes obtained from the mice in the two groups, the euchromatin appeared disperse and not readily stainable, whereas the heterochromatin appeared as a narrow peripheral band near the nuclear membrane (Figure3). The cross sections show the typical structure of the nuclear envelope with two parallel membranes and the nuclear pores as electron-dense structures embedded in the spaces where the inner and outer nuclear membranes join together (Figure3). The density of the pores in the nuclear membrane and the relative volume of nuclear euchromatin were significantly lower in the RA cardiomyocyte obtained from the OVX group compared with the sham group (p<0.05) (Table2).
DISCUSSION and OVX (B) groups showing the nucleus (N) with euchromatin (white arrows) and the pores of the nuclear membrane (black arrows).")
There are two major findings in the present study. First, the middle-aged mice subjected to ovariectomy exhibited a significant decrease in the number and size of NP granules of RA cardiomyocytes compared with the sham-operated controls. These findings indicate that estrogens enhance NP production, which has previously been reported in other animal models (9-14). Second, the number of nuclear pores and the volumetric fractions of euchromatin, mitochondria and the Golgi apparatus were significantly reduced in the mice 3 months after ovariectomy but not in the sham-operated mice. This is the first demonstration that lower levels of estrogens are associated with a morphometric reduction of the secretory apparatus in atrial cardiomyocytes.
In the present study, the mice subjected to ovariectomy exhibited a significant decrease in the number and size of NP granules of the RA cardiomyocytes compared with the sham-operated controls. The reduction in NP production was associated with increased blood pressure in accordance with previous results (27-29). This increase demonstrated the importance of NPs and estrogen in the control of blood pressure.
It is known that after menopause, blood pressure increases in women to levels even higher than found in men (28-30). The reduction in NPs after menopause could explain why hormone replacement therapy does not significantly reduce blood pressure in most postmenopausal women, which suggests that the loss of estrogens may not be the only component involved in the higher blood pressure in women after menopause. It has been suggested that estradiol protects rats from developing hypertension by increasing NO and NP levels while decreasing renin activity (15).
The present results are in accordance with recent studies in rats which have suggested that estrogen controls the production of NPs (13,31,32) and their release into the bloodstream (10,13,33,34). The mechanism for the reduction of NP synthesis observed in ovariectomized mice might be related to gene expression in RA. In a recent study, it was shown that ovariectomy significantly down-regulated NP expression in the RA, which indicates that this gene is under estrogenic control (10).
The present study demonstrated adverse effects on nuclear and cytoplasmic cardiomyocyte parameters induced by ovariectomy. The cardiomyocytes of mice that underwent ovariectomy presented a low density of nuclear pores and decreased volumetric fractions of euchromatin, mitochondria and Golgi complex. These findings are consistent with previous research that indicated that the production and secretion of NPs depend on the secretory apparatus of atrial cardiomyocytes (17), which, in turn, depends on the action of estrogens (10).
Thus, the decreased euchromatin volumetric fractions in the OVX mice and subsequent reduced RNA transcription rates associated with the decreased volume in the Golgi complex provide evidence that the rate of NP synthesis may decrease as a response to the lack of stimulation by estrogen.
Estrogens belong to a class of steroid hormones that regulate target cells upon binding to intracellular and membrane receptors. The existence of estrogen receptors has been demonstrated in cardiomyocytes (35). Estrogen binding causes conformational changes to estrogens receptors with attachment to cellular proteins that serve the transcriptional coactivators (36). In adult rats, ovariectomy has been shown to decrease atrial estrogen receptor and NP transcription (37). It is possible that the same effects occurred in the present study. In summary, in the present study, the lack of estrogens resulted in decreased functioning of the cardiomyocytes in RA mice as a result of the hypotrophy of their nuclear and cytoplasmic components, including the nuclear pores, Golgi apparatus and mitochondria, which was related to the production and secretion of NPs.
AUTHOR CONTRIBUTIONSDe Souza RR and Pithon-Curi TC were responsible for the study design, statistical analysis, data evaluation and collection, manuscript writing and critical review. De Oliveira VC contributed to the statistical analysis and data evaluation and collection. Maldonado DC contributed to the data evaluation, manuscript writing and critical review.
No potential conflict of interest was reported.