




Fenitrothion residue is found primarily in soil, water and food products and can lead to a variety of toxic effects on the immune, hepatobiliary and hematological systems. However, the effects of fenitrothion on the male reproductive system remain unclear. This study aimed to evaluate the effects of fenitrothion on the sperm and testes of male Sprague-Dawley rats.
METHODS:A 20 mg/kg dose of fenitrothion was administered orally by gavages for 28 consecutive days. Blood sample was obtained by cardiac puncture and dissection of the testes and cauda epididymis was performed to obtain sperm. The effects of fenitrothion on the body and organ weight, biochemical and oxidative stress, sperm characteristics, histology and ultrastructural changes in the testes were evaluated.
RESULTS:Fenitrothion significantly decreased the body weight gain and weight of the epididymis compared with the control group. Fenitrothion also decreased plasma cholinesterase activity compared with the control group. Fenitrothion altered the sperm characteristics, such as sperm concentration, sperm viability and normal sperm morphology, compared with the control group. Oxidative stress markers, such as malondialdehyde, protein carbonyl, total glutathione and glutathione S-transferase, were significantly increased and superoxide dismutase activity was significantly decreased in the fenitrothion-treated group compared with the control group. The histopathological and ultrastructural examination of the testes of the fenitrothion-treated group revealed alterations corresponding with the biochemical changes compared with the control group.
CONCLUSION:A 20 mg/kg dose of fenitrothion caused deleterious effects on the sperm and testes of Sprague-Dawley rats.
Pesticides such as organophosphates have been widely used in agriculture to enhance the quality of food products. They have also been utilized in public health efforts to control vector diseases by eradicating unwanted insects. Fenitrothion [O,O-dimethyl-O-(3-methyl-4-nitrophenyl) phosphorothioate] (FNT) is a broad-spectrum organophosphate insecticide that distresses the nervous system by inhibiting acetyl cholinesterase activity (1). It is employed in agriculture to control insects and mites that affect cereals, rice, fruits, vegetables, stored grains and cotton (2). FNT also acts against flies, mosquitoes and cockroaches and has been successfully used as a vector control agent for malaria in public health programs (1).
The widespread use of FNT insecticides has also caused the release of the parent compound and its metabolites into natural water sources, such as rivers and ponds, thus categorizing it as a common river water pollutant (3). It has been classified on the “red list” by the United Kingdom Environment Agency as one of the most dangerous substances to the aquatic environment (4). The presence of FNT in the environment, such as in soil, water and food products, can also lead to toxic effects on non-target organisms (3).
Humans are potentially exposed to FNT either directly through occupational exposure or indirectly via food consumption. Previous research has found that various concentrations of FNT caused histopathological effects on the liver and kidneys of rats, cytotoxic effects on the lungs of rats and immunosuppressive effects in rats (5,6). Furthermore, after a seven-day exposure of FNT at doses of 15 and 30 mg/kg, the compound acted as an anti-androgen that could reduce the growth of androgen-dependent tissues in rats (7). However, the pathophysiological effect of FNT on sperm and testes remains unclear and underreported.
These toxic effects likely occur through the generation of reactive oxygen species (ROS) that may be produced as a result of the FNT metabolism by cytochrome P450s or due to the high-energy consumption coupled with the inhibition of oxidative phosphorylation (8). The imbalance between the formation of ROS and mechanism of enzymatic and non-enzymatic antioxidants as a body defense system can lead to oxidative stress. Oxidative stress has been reported to be the primary mechanism of organophosphate toxicity after prolonged exposure (9). In addition, oxidative stress conditions may cause alterations in sperm cells due to the high levels of polyunsaturated fatty acid (PUFA) in their plasma membrane (10).
FNT at a dose of 20 mg/kg body weight (bw) by oral administration caused significant damage to the liver and kidneys (5) and changes in the histology of lungs (6) in male albino rats. Therefore, this study aimed to assess the potential effects of FNT at a dose of 20 mg/kg bw on the reproductive parameters of male Sprague-Dawley rats to provide a potential extrapolation of the findings to humans. The purpose of this study was to determine the effects of FNT on sperm characteristics, sperm and testicular morphology, biochemical and oxidative stress markers in testes of male Sprague-Dawley rats.
MATERIALS AND METHODSAnimalsTwenty adult male Sprague-Dawley rats weighing 230-250 g were used in this study. All rats were obtained from the Laboratory Animal Resource Unit, Faculty of Medicine, Universiti Kebangsaan Malaysia, one week prior to the study for acclimatization purposes. The rats were kept in plastic cages, with two rats per cage under standard environmental conditions (12 h light/dark cycles, 25-28°C) and maintained on a standard pellet and water ad libitum diet for the duration of the study. All of the animal handling protocols were approved by the Animal Ethics Committee of Laboratory Animal Resource Unit, Faculty of Medicine, Universiti Kebangsaan Malaysia.
ChemicalsThe chemicals and reagents used in this study were of high purity. FNT with a purity of 99.9% was purchased from SUPELCO Analytical, USA with lot number: LB75917. Other chemicals used in this study were purchased from Sigma Chemicals Co., St. Louis, Missouri, USA, with the exception of 2,4-nitrophenylhydrazine, which was purchased from Mallinckrodt Chemicals, Lenzing AG, Salzburg, Austria. Nitro blue tetrazolium, reduced glutathione and other reagents were purchased from Merck, Darmstadt, Bundesland of Hesse, Germany.
Animal treatment scheduleAfter the one-week acclimatization, the rats were divided randomly into control and treated groups, with ten rats per group. FNT (dissolved in corn oil) was administered at a dose of 20 mg/kg bw (1/3 LD50) using oral gavages for 28 consecutive days (five days/week), whereas the control group received only corn oil (1 mg/kg bw). Toxicity signs were observed and recorded daily. Body weight was recorded weekly throughout the study duration.
At the end of the study, the rats were fasted overnight and anaesthetized with diethyl ether. Blood was collected in an EDTA tube via cardiac puncture. Plasma was obtained by centrifugation of blood samples at 3,500 rpm at 4°C for 10 minutes. Plasma samples were maintained at -40°C until further biochemical analysis. At the end of the experiment, the rats were sacrificed and dissected. The weights of the cauda epididymis, epididymis and testes were recorded. Left and right of cauda epididymis were immediately placed in 2 ml of Hank's Buffered Salt Solution (HBSS) enriched with 0.5% BSA and pre-warmed at 37°C to obtain the sperm. The cauda epididymis was cut into small pieces and centrifuged at 1,000 rpm at 4°C for 3 minutes to allow the sperm to be released for further analysis. Testes were homogenized in 0.15 M KCl solution (1:2) at 4°C and centrifuged at 10,000 X g for 20 minutes at 4°C. The homogenated testes were maintained at -40°C until further biochemical analysis.
Biochemical analysisPlasma cholinesterase (ChE) was assayed using the Ellman method (11). The cholinesterase in the samples hydrolyzed the propionythiocholine to form thiocholine, which was then reacted with 5,5′-dithiobis-2-nitrobenzoic acid (DTNB) to form a yellow solution containing 5-thio-2-nitrobenzoate. The formation of the yellow color was monitored at 405 nm and the cholinesterase activity was calculated in U/l.
Protein and cholesterol levels in the testes were determined by the Bradford (12) and Franey and Amador (13) methods, respectively. Briefly, an aliquot of the homogenated testes was added to Bradford's reagent and incubated for 5 minutes at room temperature. The protein reacted with Bradford's reagent to produce colored complexes and the intensity of these complexes was measured at 595 nm using a microplate reader. Similarly, cholesterol detection was based on the reaction between cholesterol, ferric chloride and concentrated sulfuric acid. Briefly, 100 μl of homogenate testes was added to ethanol for cholesterol extraction purposes. The obtained supernatant was treated with ferric chloride and sulfuric acid. The concentration of cholesterol in each sample was determined at 560 nm, where it exhibited a color change from yellow to brown. The protein and cholesterol levels were calculated based on their standard curves in mg/dl and mg/ml, respectively.
The homogenated testes were evaluated for malondialdehyde (MDA), protein carbonyl (PC), total glutathione (GSH), glutathione S-transferase (GST) and superoxide dismutase (SOD) to assess the redox processes. The production of MDA was measured for lipid peroxidation based on the Stocks and Dormandy methods (14). Briefly, an aliquot of (0.5 ml) of homogenated testes was added to a TCA/HCl solution, vortexed and incubated at room temperature for 15 minutes. The mixture was added to TBA/NaOH, vortexed and heated in a boiling water bath (100°C) for 30 minutes. The MDA in the sample reacted with the thiobarbituric acid to form a pink chromogen containing thiobarbituric acid reactive substances (TBARS). The level of TBARS in the supernatant was measured using a spectrophotometer at 532 nm. The MDA concentration was expressed as mM/mg of tissue protein.
The PC content was measured for oxidative stress in the protein precipitate using the method described by Levine (15) with some modifications. Briefly, the homogenated testes were added to TCA (1:1) to precipitate the protein, incubated in ice for 15 minutes and then centrifuged at 15,000 X g for 5 minutes at 4°C. The obtained pellet was reacted with 10 mM DNPH and incubated for 1 hour at room temperature in the dark. TCA was added to the mixture to precipitate the protein; the mixture was incubated in ice for 5 minutes and then centrifuged at 15,000 X g for 5 minutes at 4°C. The obtained pellet was then washed twice using a mixture of ethanol and ethyl acetate and then diluted with 5 M urea. The derivatives obtained after centrifugation at 15,000 X g for 3 minutes at 4°C were monitored between 375 and 380 nm. PC formation was expressed as nmol/mg of tissue protein.
GSH content was measured in the homogenated testes based on the Ellman method (16) with some modification. Deproteinized homogenated testes were treated with a metaphosphoric acid solution and centrifuged at 3,000 X g for 10 minutes at 4°C. The obtained supernatant was mixed with a reaction buffer at pH 8.0 and DTNB for 15 minutes and measured at 412 nm by using a microplate reader. The GSH content was expressed as mmol/mg protein.
SOD and GST enzyme activity was determined using the Beyer and Fridovich (17) and Habig and Jacoby (18) methods, respectively. Briefly, an aliquot of (20 μl) of the homogenate was mixed with the substrate containing [PBS(EDTA); L-methionine; NBT.2HCl; Triton-X] and riboflavin. The mixture was incubated in an aluminum box under 20 watt lamp for 7 minutes. The SOD activity was measured spectrophotometrically by monitoring the inhibition of ferricytochrome reduction using a xanthine-xanthine oxidase as a source of peroxides. An SOD unit is defined as one unit of enzyme, which inhibits 50% of the nitro blue tetrazolium (NBT) reduction. SOD activity was calculated and expressed in U/mg protein. GST was assayed by measuring the rate that the enzyme catalyzed the conjugation of reduced glutathione with CDNB. Briefly, 50 μl of homogenated testes was added to 1 mM GSH and 75 μM of 1-chloro-2,4-dinitro benzene (CDNB). The rate of GST activity was measured at 340 nm and expressed as U μM/min/mg protein.
Sperm viabilitySperm viability was determined by a routine gold standard method suggested by WHO (19) involving the determination of eosin, which penetrates the membranes of damaged cells. Approximately 100 μl of sperm suspension was mixed with two drops of eosin 1% and allowed to remain undisturbed for 30 seconds. Then, three drops 10% nigrosin were mixed with the solution and within 30 seconds, a thick smear was performed in triplicate. The results were determined by counting both motile and immotile sperm and are presented as a percentage (%).
Sperm density and morphologySperm density was calculated by a Markler Counting Chamber based on a WHO method (19). Briefly, 10 μl of a sperm suspension was calculated for at least ten different microscopic fields under 40X magnification using a light microscope. Results are expressed as 106 cells/ml.
A smear was conducted by using a drop of sperm suspension. After the smear had dried, the slide was fixed with absolute ethanol for 5 minutes. Then, the slide was immersed in Diff-Quik Stain I and II for 5 minutes each. A cover slip was applied and two hundred sperms were calculated per animal to measure the morphological abnormalities under oil immersion. The abnormal sperm heads were calculated in triplicate. Data are shown as a percentage of sperm head abnormalities.
Morphological analysisThe testes in both groups were treated with 10% buffered formalin solution and a routine histological procedure was conducted. All testicular sections were stained with hematoxylin and eosin (H & E) stains and monitored for morphological changes under X10 and X40 magnifications. All changes were verified by a histopathological expert.
A section of the testes was cut into small pieces (1 mm3), treated with 2.5% glutaraldehyde 0.1 N PBS at room temperature for one hour and post-fixed with osmium tetraoxide for another hour. Testes tissue was dehydrated in 70, 90 and 100% (twice) acetone for five minutes each, followed by 1:1 (acetone: resin) for five minutes and then embedded in epoxy resin. Ultrathin slices (90 nm) were observed using a transmission electron microscope, Tecnai G2 (FEI, USA), at 100 kV.
Statistical analysisAll obtained data from the analysis were normally distributed. The differences between the treated and control groups were statistically evaluated using an independent Student's t-test. All data are expressed as the mean values±SD, with significant values at p<0.05.
RESULTSSigns of toxicityAdministration of a 20 mg/kg bw dose of FNT caused cholinergic signs, such as hypoactivity, lacrimation, piloerection and tremor. All of the toxicity signs occurred within one or two hours after FNT administration. The toxicity signs remained in the same animals for approximately 48 hours throughout the experimental period. Approximately 60% of treated rats showed signs of toxicity. No deaths were recorded in either experimental group.
Body and organ weightThe body and organ weight of the FNT and control groups are provided in Table 1. The treated and control groups showed increased body weight throughout the experimental period. However, the rats treated with FNT showed a significantly lower weight-gain compared with the control group (p<0.05). The absolute and relative epididymis weight decreased significantly for the FNT group compared with the control group, with p<0.05 and p<0.001, respectively. The relative weight of the testes was significantly higher in the FNT group than control group (p<0.05). No statistically significant differences were noted in absolute and relative weight of cauda epididymis between the treated and untreated groups.
Body and reproductive organ weights of the treated and untreated groups. Data are reported as the mean values±SD. Statistically significant differences compared with the control group are reported when p<0.05a and p<0.01b.
| Weight (g) | Experimental Groups | |
|---|---|---|
| Control | FNT(20 mg/kg bw) | |
| Initial Body Weight | 243.1±14.64 | 236.3±13.40 |
| End of Body Weight | 326.9±26.01 | 295.9±23.81a |
| Weight Gain | 83.30±7.59 | 59.60±8.57a |
| Absolute Weight | ||
| Testes | 2.54±0.45 | 2.68±0.23 |
| Epididymis | 0.94±0.35 | 0.53±0.08a |
| Cauda Epididymis | 0.40±0.15 | 0.39±0.03 |
| Relative Weight | ||
| Testes | 0.77±0.14 | 0.91±0.09a |
| Epididymis | 0.34±0.07 | 0.18±0.03b |
| Cauda Epididymis | 0.12±0.05 | 0.13±0.02 |
The results of the biochemical analysis are provided in Table 2. A marked reduction in the cholinesterase activity was found for the FNT group compared with the control group (p<0.05). However, testicular cholesterol and protein levels were not significantly different between the treated and untreated groups. FNT not only increased the formation of MDA (p<0.05) and PC (p<0.01) but also increased the content of GSH and activity of GST compared with the control group (p<0.05). However, SOD activity decreased in the FNT treated group compared with the control group (p<0.001).
Biochemical and oxidative stress analysis of the treated and untreated groups. Data are reported as the mean values±SD. Statistically significant differences compared with the control group are reported when p<0.05a, p<0.01b and p<0.001c.
| Experimental Groups | ||
|---|---|---|
| Control | FNT(20 mg/kg bw) | |
| Plasma cholinesterase (U/ml) | 0.975±0.39 | 0.208±0.13b |
| Protein Testes (mg/ml) | 34.81±1.90 | 31.96±0.98 |
| Cholesterol Testes (mg/ml) | 0.22±0.03 | 0.24±0.02 |
| SOD (U/mg protein) | 1.23±0.11 | 0.62±0.06c |
| GSH (mmol/mg protein) | 0.013±0.003 | 0.035±0.007a |
| GST (μM/min/mg protein) | 6.47±0.36 | 8.28±0.52a |
| MDA (mM/mg protein) | 0.060±0.008 | 0.130±0.020a |
| PC (nmol/mg protein) | 0.035±0.009 | 0.096±0.017b |
SOD: superoxide dismutase; GSH: total glutathione; GST: glutathione S-transferase; MDA: malondialdehyde; PC: protein carbonyl.
The sperm concentration, sperm viability and sperm head abnormality data are provided in Table 3. A significant decrease in sperm concentration was found in the FNT group (p<0.05). Furthermore, FNT significantly decreased sperm viability and normal sperm morphology compared with the control group (p<0.05).
Sperm analysis of the treated and untreated groups. Data are reported as the mean values±SD. Statistically significant differences compared with the control group are reported when p<0.05a.
| Experimental Groups | ||
|---|---|---|
| Control | FNT(20 mg/kg bw) | |
| Sperm density (X106 cells) | 68.77±3.69 | 35.71±3.13a |
| Sperm non-viability (%) | 7.24±1.46 | 38.61±5.75a |
| Abnormal sperm morphology (%) | 11.33±0.99 | 57.33±3.98a |
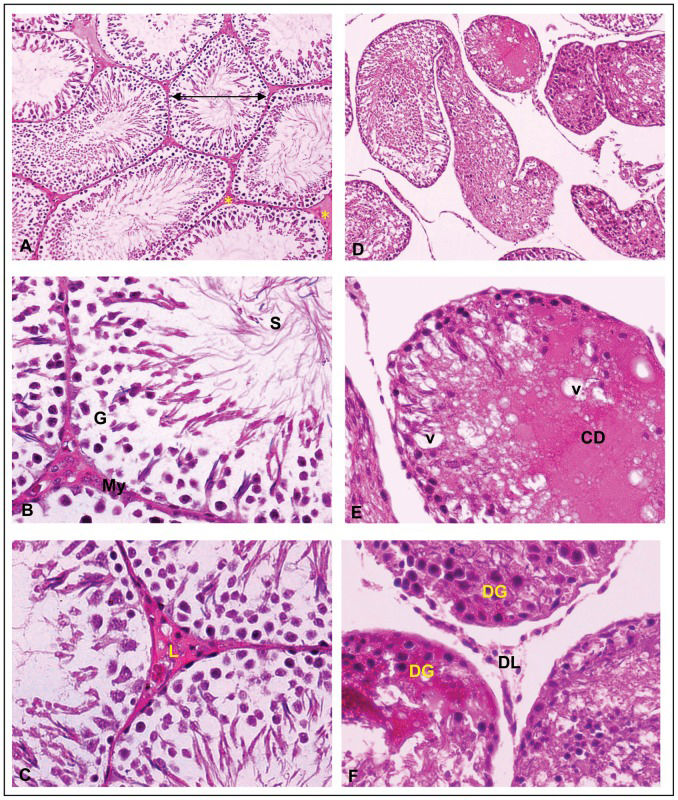
The normal histological structure of rat testes is illustrated in Figure 1 (A-C). The seminiferous tubule consists of normal somatic and spermatogenic cells and is surrounded by peritubular myoid cells (Figure 1B). Clusters of Leydig's cells were observed in the intertubular space that is in close contact with blood vessels and lymphatic channels (Figure 1C). However, histopathological observations such as degeneration of germ cells, expansion of interstitial space, disarrangement of spermatogonia in seminiferous tubule, degeneration of Leydig cells and the presence of cellular debris throughout the lumen of the seminiferous tubule of rat testes were observed in the FNT group (Figure 1 D–F).
. Figure 1 (A–C): Normal histological structure of rat")
Histology of the testes of treated and untreated group (H&E staining). Figure 1 (A–C): Normal histological structure of rat's testis. Normal structure of seminiferous tubules (⇒) and interstitial tissue (∗)(Figure 1A). The seminiferous tubule consists of normal somatic, sperm (S) and spermatogenic cells (G) surrounded with peritubularmyoid cells (My)(Figure 1B). Clusters of Leydig's cells (L) were observed in the intertubular space that was in close contact with blood vessels and lymphatic channels (Figure 1C). Figure 1(D–F): Histology of FNT rat testis showing: degenerative of germ cells (DG) and Leydig cells (DL), expansion of interstitsial space, cellular debris (CD), vacuoles (v) and disarrangement of spermatogonia throughout the lumen of seminiferous tubule. (Magnification A and D∼X10; Magnification B, C, E and F∼X40).
The ultra-structure of the rat testis showed morphological alterations in the treated groups compared with the control group (Figure 2). Abnormalities included the presence of a lipid droplet, increased number of mitochondria, less chromatin condensation in the late spermatid and a vacuolated mitochondrial helix (Figure 2 D-F). However, histopathological and ultra-structure alterations were only observed in approximately 40% of the treated rat population.
: The normal ultra structure of control rat testis showing: dense of chromatin (K) in late spermatid and normal structure of sperm tail. Figure 2 (D-F): The ultra structure of FNT rats testis showing: presence of lipid droplet (L), increased number of mitochondria (M), less chromatin (K) condensation in late spermatid and vacuolated of mitochondriae helix (red arrow) (Bar A & D∼5000 nm; Bar B & E∼2000 nm; Bar C & F∼1000 nm).")
Ultra structure of testes of treated and untreated group. Figure 2 (A–C): The normal ultra structure of control rat testis showing: dense of chromatin (K) in late spermatid and normal structure of sperm tail. Figure 2 (D-F): The ultra structure of FNT rats testis showing: presence of lipid droplet (L), increased number of mitochondria (M), less chromatin (K) condensation in late spermatid and vacuolated of mitochondriae helix (red arrow) (Bar A & D∼5000 nm; Bar B & E∼2000 nm; Bar C & F∼1000 nm).
The extensive use of organophosphate in the environment has affected many non-target organisms, including humans. The high lipid solubility of fenitrothion may lead to the bioaccumulation of the parent compound and its toxic metabolites in the adipose tissue. The release of this compound from the fat storage could be enhanced in stressful situations, such as during illness and diet changes (20). The current toxicity scenarios have attracted the attention of researchers studying the toxic effects of organophosphate on the male reproductive system. FNT has been found to cause some reproductive abnormalities in Japanese sprayers, with a reduction in the sperm quality (21).
This study showed that the administration of FNT at a dose of 20 mg/kg bw for 28 days in male Sprague-Dawley rats induced significant adverse effects, including toxicity signs and symptoms, alterations in the biochemical status, reductions in sperm quality and histopathological and ultrastructural changes in the testes. FNT administration inhibits the acetyl cholinesterase enzymes that cause acetylcholine accumulation in cholinergic synapses. Therefore, cholinergic signs were observed, including hypoactivity, lacrimation, piloerection and tremor. Significant inhibition of acetyl cholinesterase enzyme by FNT directly supports the common acute toxicity mechanism of organophosphate poisoning (1). These findings are generally consistent with the studies by Turner (22) who reported that the muscle tremor was observed in rats treated with FNT at doses of 20 and 25 mg/kg/day.
A significant decrease in body weight gain was observed in the FNT treated group; this might be associated with the toxic symptoms, such as cholinergic signs. The reduced weight gained could be attributed to systemic toxicity in male rats (23). FNT significantly decreased epididymis weight but increased testis weight. In this study, the reduction in the weight of reproductive organs, such as the epididymis, could be due to the reduction of androgen availability (23). The increase in the weight of the testes induced by chemicals can be correlated with testicular swelling, followed by atrophy (24).
Under pathological conditions, high levels of ROS induce lipid peroxidation in male reproductive system (25). FNT increased MDA and PC production, which could be used as important biomarkers of lipid and protein peroxidation. These findings support a previous study that found that most of the organophosphate insecticides altered MDA levels in experimental animals, especially in rats (5). PUFA peroxidation in the sperm head leads to a loss of membrane fluidity and decrease in the activity of the membrane enzyme and ion channels, which in turn increases the sperm permeability, causing sperm morphological abnormalities and decreased sperm function (26).
ROS may also initiate a chain of reactions that ultimately leads to apoptosis. The process of apoptosis may also be accelerated by ROS-induced DNA damage and ultimately decrease the sperm count (26). Decrease in spermatogenesis can explain the occurrence of sperm with excessive residual cytoplasm in the mid-piece, thus altering sperm morphology. The abnormal morphology of sperm may occur in the epididymis and seminiferous tubules (27). The above condition might explain the alterations in the sperm morphology of the FNT-treated group.
During the maturation process of sperm, cytoplasmic enzymes are extruded at the final stages of maturation. Therefore, mature sperm can attain their characteristic morphology and prevent oxidative stress, thus altering sperm functions, such as viability (27). In this study, FNT decreased the percentage of sperm viability. Decreases in sperm production and viability and increases in abnormal sperm morphology are common hazardous effects of organophosphate insecticides on sperm parameters (28). However, the supplementation of Eurycoma longifolia Jack increased the epididymal sperm quality, such as sperm count and motility, of estrogen-treated rats (29).
Due to the lack of the cytoplasmic enzyme in mature sperm, the male genital tract, including the testes and semen, are rich in both enzymatic and non-enzymatic antioxidants. Several mechanisms of enzymatic and non-enzymatic antioxidants in humans and animals can counteract the harmful effects caused by ROS (9). In this study, SOD activities significantly decreased in testicular tissues of the FNT-treated group. These findings have also been reported in rat tissues induced by other organophosphate insecticides (30). Furthermore, the activity of melatonin as an antioxidant was also found to decrease in rats exposed to 2.45 GHz microwave radiation (31). However, phosphodiesterase type 5 increased total antioxidant levels, explaining the acute beneficial effect of this compound in patients with erectile dysfunction (32).
This study also found that FNT increased GSH content and GST activity. Although GSH is an early consequence of oxidative stress, the increase in the GSH content of testes treated with FNT could be due to the early stage of the detoxification process by GST. GST is an enzymatic antioxidant that is involved in the detoxification of organophosphate insecticides to form a non-toxic product (33). Organophosphate insecticides have been reported to alter the biochemical status in experimental animals depending on the various types of organophosphates, exposures durations and doses (34). There is a slight difference in the total protein and cholesterol content in testes of the FNT group. Changes in protein and cholesterol levels in the testes have been attributed to the inhibition of the androgen concentration (34).
The above findings were supported by the histopathological changes in sperm and testis induced by FNT. FNT caused pathological changes in seminiferous tubules following a 28-day exposure. The apoptosis induced by ROS helped remove abnormal germ cells in the testes and prevent their overproduction by activating caspases (10). This explained the degenerative nature of germ and Leydig's cells found in the seminiferous tubule and interstitial tissues of the FNT-treated group. FNT also allowed the crossing of the blood-testes barrier, thereby decreasing the spermatogenesis. These findings supported the study performed by Turner (22) who found that FNT inhibited the maturation and differentiation of germ cells. Toxicants can directly disturb the function of Sertoli cells, thereby causing the disorganization of germ cells in the seminiferous tubule of the FNT-treated group. In this study, FNT increased the formation of vacuoles and number of mitochondria in the testes. Vacuoles can be described as an early stage of damage induced by any toxicants (35). In addition, the increase in the number of mitochondria may be attributed to the high-energy consumption of the redox process in testes (9). FNT also caused fewer chromatins in late spermatids and vacuolated mitochondria of the sperm tail. A decrease in the chromatin in late spermatids may be due to genetic abnormalities that occurred during the differential process in spermatogenesis (28).
In conclusion, FNT at a dose of 20 mg/kg bw has a detrimental effect on the sperm and testes of Sprague-Dawley rats.
The authors would like to thank the Ministry of Higher Education and Universiti Kebangsaan Malaysia for research funds (UKM-GUP-2011-126). We also extend our gratitude to the Director of Institute for Medical Research (IMR), Malaysia, for the permission to use the electron microscopy facilities. The authors gratefully acknowledge the staff of Programme of Biomedical Sciences, School of Diagnostic and Applied Health Sciences, Faculty of Health Sciences, Universiti Kebangsaan Malaysia, and Electron Microscopy Unit, Institute for Medical Research (IMR), Malaysia, for providing the research facilities. We would like to acknowledge all of the lecturers, researchers and those who directly or indirectly supported this research.
No potential conflict of interest was reported.
Taib IS performed the majority of the experiments and drafted the manuscript. Budin SB was the project leader and was responsible for the project design. Ghazali AR and Mohamed J revised the manuscript for important intellectual content. Jayusman PA and Louis SR performed some of the experiments. All authors read and approved the final manuscript.