







In this study, we tested the hypothesis that hypertonic saline exerts anti-inflammatory effects by modulating hepatic oxidative stress in pancreatitis.
INTRODUCTION:The incidence of hepatic injury is related to severe pancreatitis, and hypertonic saline reduces pancreatic injury and mortality in pancreatitis.
METHODS:Wistar rats were divided into four groups: control (not subjected to treatment), untreated pancreatitis (NT, pancreatitis induced by a retrograde transduodenal infusion of 2.5% sodium taurocholate into the pancreatic duct with no further treatment administered), pancreatitis with normal saline (NS, pancreatitis induced as described above and followed by resuscitation with 0.9% NaCl), and pancreatitis with hypertonic saline (HS, pancreatitis induced as described above and followed by resuscitation with 7.5% NaCl). At 4, 12, and 24 h after pancreatitis induction, liver levels of inducible nitric oxide synthase (iNOS), heat-shock protein 70, nitrotyrosine (formation of peroxynitrite), nitrite/nitrate production, lipid peroxidation, and alanine aminotransferase (ALT) release were determined.
RESULTS:Twelve hours after pancreatitis induction, animals in the HS group presented significantly lower iNOS expression (P<0.01 vs. NS), nitrite/nitrate levels (P<0.01 vs. NS), lipid peroxidation (P<0.05 vs. NT), and ALT release (P<0.01 vs. NS). Twenty-four hours after pancreatitis induction, nitrotyrosine expression was significantly lower in the HS group than in the NS group (P<0.05).
DISCUSSION:The protective effect of hypertonic saline was related to the establishment of a superoxide-NO balance that was unfavorable to nitrotyrosine formation.
CONCLUSIONS:Hypertonic saline decreases hepatic oxidative stress and thereby minimizes liver damage in pancreatitis.
Acute pancreatitis causes systemic inflammatory response syndrome (SIRS) and multiple organ failure (MOF). In severe cases, the mortality rate is high.1 Severe acute pancreatitis has been shown to correlate with hepatic injury.2 Although the liver is known to be a primary target of cytokines released in the blood by the pancreas, the liver itself releases inflammatory substances, such as reactive oxygen species, leading to injury of distant organs.3 In addition, hepatic oxidative stress plays an important role in the pathophysiology of many gastrointestinal diseases.4
Free radicals actively participate in the pathophysiological process that results in hepatocyte apoptosis.5 However, nitric oxide (NO) plays a dual role in inflammation. Studies suggest that the expression of inducible nitric oxide synthases (iNOS) increases hepatocellular injury in hemorrhagic shock and ischemia/reperfusion injury under conditions of increased superoxide production. Peroxynitrite is a highly reactive oxidant produced spontaneously by the combination of NO and superoxide anions at rates approaching the diffusion limit.6 In addition to its capacity to hydroxylate and nitrate aromatic rings, peroxynitrite is capable of oxidizing lipid membranes and sulfhydryl moieties.6 Peroxynitrite formation is thought to be a late event that contributes to cell death and tissue injury in human sepsis.7,8 These findings are consistent with the hypothesis that peroxynitrite is ultimately responsible for pathophysiology previously attributed to NO alone. Because NO has been shown to protect cells and tissues, the inhibition of NO can also result in negative outcomes.9 In particular, NO is hepatoprotective under conditions that do not involve redox stress.10
In experimental animal models of pancreatitis, hypertonic saline has been shown to alter circulating plasma volume, reduce trypsinogen levels, prevent acinar necrosis, reduce inflammatory cytokine levels, and avert pancreatic infection, which minimizes injury and reduces mortality.11,12 The administration of hypertonic saline in a rat model of acute pancreatitis reduced systemic inflammation rather than protecting local (pancreatic) tissue.13 Previous studies showed that hypertonic saline modulates pancreatitis-related injury to the lungs14 and liver.15 This effect was attributed to the modulated expression and activity of various proteins, such as metalloproteinases, collagen, and members of the heat-shock protein (Hsp) family.
Although hepatic injury is known to play a major role during pancreatitis16, the effect of hypertonic saline on oxidative stress in the liver has not been reported in the literature. However, hypertonicity is known to regulate protein expression,17 oxidative stress,18 and intracellular signaling cascades.19 We therefore examined the effects of hypertonic saline on hepatic oxidative stress in experimental pancreatitis.
MATERIAL AND METHODSAnimalsMale Wistar rats weighing 270–320 g were obtained from the animal facilities of the University of São Paulo School of Medicine, which is located in São Paulo, Brazil. The experimental protocol was approved by the Research Ethics Committee of the School of Medicine.
Pancreatitis inductionThe rats were anesthetized using subcutaneous injections of ketamine (10 mg/kg) and xylazine (8 mg/kg). Acute pancreatitis was then induced by a retrograde transduodenal infusion of 2.5% sodium taurocholate (1.0 ml/kg; Sigma, St. Louis, MO, USA) into the pancreatic duct via a 24-gauge angiocatheter at a constant infusion rate of 1 ml/min (Machado et al. 2006). The bile duct was clamped with a small bulldog clamp at the hepatic hilum to prevent the sodium taurocholate from leaking into the duodenum or refluxing into the liver. The hepatic hilar clamp was released after injection. The animals were divided into four groups: control (C), consisting of animals that were not subjected to treatment; untreated pancreatitis (NT), consisting of animals that received induction of pancreatitis (as described above) with no further treatment; pancreatitis with normal saline (NS), consisting of animals that received induction of pancreatitis (as described above) followed by resuscitation with 34 ml/kg of 0.9% NaCl given as an intravenous bolus over a period of 1 h in 5 min increments; and pancreatitis with hypertonic saline (HS), consisting of animals that received induction of pancreatitis (as described above) followed by resuscitation with 4 ml/kg of 7.5% NaCl administered via the internal jugular vein over a period of 1 hr in 5 min increments. Our main objective was to study the effects of tonicity. Therefore, the sodium content of the total volume of normal saline infused was equivalent to that of 4 ml/kg of hypertonic saline. At 4, 12, and 24 h after the induction of pancreatitis, animals were sacrificed, and the livers were collected.
Semi-quantitative reverse transcriptase-polymerase chain reaction (RT-PCR)Semi-quantitative RT-PCR was used to determine iNOS mRNA levels in liver tissue. Total RNA was extracted from frozen rat livers with TRIzol reagent (Invitrogen, Carlsbad, CA, USA), which was in accordance with the manufacturer's instructions. The RNA was dissolved in diethyl pyrocarbonate-treated water and the concentration was quantified spectrophotometrically at 260 nm. First-strand cDNA was generated by adding 1 μg of RNA to a mixture containing 1 μl of reverse transcriptase (ImProm-II™; Promega, Madison, WI, USA), 1 μl (0.5 μg/μl) of oligo(dT), 20 U/μl recombinant RNAsin® RNAse inhibitor, 3 mM MgCl2, 6 μl of ImProm-II™ 5× reaction buffer (Promega), and 1 μl (0.5 mM) of deoxynucleoside triphosphate (dNTP) mix (Invitrogen), which resulted in a final volume of 20 μl. Reverse transcription was performed at 42°C for 50 min and was followed by heat inactivation of the reverse transcriptase at 70°C for 10 min. Amplification via PCR was performed using a programmable thermal controller (PTC-200; MJ Research, Watertown, MA, USA). The PCR solution contained 1 μl of first-strand cDNA, 2.5 μl of 10× PCR buffer, 2 mM MgCl2, 0.5 mM dNTP mix, 1 pmol/μl each of specific primers, and 2.5 U/μl TaqDNA polymerase (Invitrogen), which resulted in a final volume of 25 μl. To compare the relative abundance of a transcript between samples, RT-PCR with 18S rRNA primers was performed as an internal control. PCR products were resolved by electrophoresis on an ethidium bromide-stained 1% agarose gel in a Horizon 58 electrophoresis unit (Life Technologies, Gaithersburg, MD, USA) and visualized under ultraviolet light with a video imaging system (Syngene Photo Image System (Syngene, Cambridge, United Kingdom). Densitometric analyses of ethidium bromide-stained gel bands were performed with Gene Tools analysis software (Syngene, Cambridge, UK). The data were plotted as a function of the log optical density of the target gene product in relation to the log optical density of the 18S rRNA. The sequences of the specific primers (Invitrogen) were as follows: GAAAGATGGTGAACTATGCC and TTACCAAAAGTGGCCCACTA, respectively, were the sequences for the sense and antisense rRNA primers (amplicon length: 320 bp), and TTCTTTGCTTCTGTGCTTAATGCG and GTTGTTGCTGAACTTCCAATCGT, respectively, were the sequences for the sense and antisense iNOS primers (amplicon length: 170 bp). Amplification consisted of 30 cycles at 95°C for 1 min, 60°C for 1 min, and 72°C for 1 min, followed by an extension at 70°C for 10 min. The results consist of the mean from 5 animals per group.
Western blottingTo analyze HSP70 protein expression, frozen tissue samples (100 mg) were pulverized in liquid nitrogen and then homogenized in a buffer containing 1% TX-100, 20 mM Tris (pH 8.0), 10% glycerol, 135 mM NaCl, and proteolytic enzyme inhibitors (40 μg/ml phenylmethylsulfonyl fluoride and 10 μg/ml pepstatin; Sigma). After the separation of the debris by centrifugation for 45 min at 14,000 g, the supernatants were preserved, and the protein concentration was determined using the Bradford method (Bradford protein assay kit; Bio-Rad, Hercules, CA). The samples were stored at −80°C until assayed. Protein expression was determined using electrophoresis on a sodium dodecyl sulfate (SDS)-polyacrylamide gel under reducing conditions. Liver tissue extracts (25–100 μg/ml) were boiled in equal volumes of loading buffer (150 mM Tris-HCl, pH 6.8; 4% SDS; 20% glycerol; 15% β-mercaptoethanol; and 0.01% bromophenol blue) and were electrophoresed on 10% polyacrylamide gels. After electrophoretic separation, proteins were transferred to Hybond-P membranes (Amersham Pharmacia Biotech, Buckinghamshire, UK) for Western blotting. Membranes were blocked with 5% non-fat dry milk in Tris-buffered saline and 0.5% Tween 20 (TBST) for 1 h. We employed the following primary antibodies: goat polyclonal anti-Hsp70 (1:1000, sc1060; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and mouse anti-β-actin (1:10000, A5441; Sigma). The primary antibodies were incubated overnight at 4°C, and the membrane was washed twice in TBST. A secondary horseradish-peroxidase-conjugated antibody (goat anti-rabbit polyclonal or rabbit anti-goat, sc2004 or sc2768, respectively; Santa Cruz Biotechnology) was then applied at a dilution of 1:5000 for 2 h. Over a 30-min period, the blots were washed twice in TBST, and then they were then incubated in enhanced chemiluminescence reagents (Super Signal Detection Kit; Pierce, Rockford, IL, USA) and exposed to photographic film (O-OMAT-AR; Eastman Kodak, Rochester, NY, USA). The band intensities of the original blots were quantified using Image J software (Research Services Branch, National Institutes of Health, Bethesda, MD, USA) and were normalized against control levels.20 The results consist of the mean from 4 animals per group.
ImmunohistochemistryPeroxynitrite formation was identified through immunohistochemical detection of tyrosine nitration (nitrotyrosine) in the liver. Formalin-fixed tissues were embedded in paraffin and sectioned at 4 μm. The paraffin was then removed from the sections and the sections were rehydrated. They were then digested with sodium citrate. Nonspecific adsorption was minimized by incubating the samples in 0.3% H2O2 for 50 min, followed by blocking with biotin-avidin for 10 min and 5% casein for 5 min. The sections were mounted on slides, which were then incubated overnight with a nitrotyrosine antibody (1:1000, A21285; Molecular Probe, Eugene, OR, USA) at 4°C. Anti-rabbit IgG (Vectastain® ABC kit, PK6101; Vector Laboratories, Burlingame, CA, USA) was used as a secondary antibody. We used 3,3′-diaminobenzidine (DAB kit, D5637; Sigma) to develop the positive reaction, which appeared as a brown color. Slides were counterstained with hematoxylin. Images were acquired using an image analysis system (Q500 iW; Leica Imaging Systems, Cambridge, UK) at a magnification of ×200. The percent amount of brown staining (staining for nitrotyrosine) in each area was quantified. The total tyrosine nitration was calculated as the mean of 10 areas per slide. The results represent mean nitration in 8 animals per group.
Nitrite and nitrateTissue concentrations of nitrite were determined using the Griess reaction to produce a color that would be detectable at the visible wavelength of 550 nm. The nitrate content of the sample was reduced to nitrite after enzymatic conversion by nitrate reductase; 25 μl of nitrate reductase and 25 μl of reduced nicotinamide adenine dinucleotide phosphate were added to 50-μl samples prepared with a Tx-100 buffer. A blank was prepared in the same buffer. The mixtures were incubated for 2 h at 25°C. We then added 100 μl of the Griess reagent, which consisted of 1% sulfanilamide in 1 N HCl, 15% N-(1-naphthyl) ethylenediamine dihydrochloride, and we incubated the samples for 10 min at room temperature. At the end of the reaction time, the absorbance was measured on a microplate reader (Infinite F500; Tecan, Salzburg, Austria) at a wavelength of 550 nm. Calibration curves were prepared by diluting sodium nitrite and potassium nitrate in distilled water. The results consist of the mean from 5 animals per group.
Lipid peroxidationLipid peroxidation was evaluated by determining the level of thiobarbituric acid reactive substances (TBARS). Liver tissue was homogenized in 1.15% KCl (100 mg/ml), transferred to clean tubes, and centrifuged at 14,000 g for 20 min at 4°C. The supernatant was collected and transferred to new receptacles. Aliquots of the tissue homogenates (100 μl each) were then added to the other reagents (750 μl of 20% acetic acid, 100 μl of 8.1% SDS, 300 μl of water, and 750 μl of 0.8% thiobarbituric acid). The samples were then placed in glass tubes and incubated at 95°C for 60 min. The absorbance of the final solution was measured at 532 nm. The results consist of the mean from 8 animals per group.
Serum enzyme activityPlasma was collected from the animals in all of the groups to determine the serum levels of alanine aminotransferase (ALT), which were measured with an automatic analyzer (ADVIA® 1200; WB Saunders Company, Philadelphia, PA, USA). The results consist of the mean from 6 animals per group.
Statistical analysisAll values are expressed as the means ± standard errors. The analyses were performed using Sigma Stat Statistical Software, version 3.1 (Sigma Stat Software Inc., Chicago, IL, USA). Comparisons among groups were performed by analysis of variance (ANOVA), which was followed by Tukey's post hoc tests to compare individual groups at the various time points. P<0.05 was considered significant.
RESULTSGene expression of iNOSAt 4, 12, and 24 h after the induction of pancreatitis, the gene (mRNA) expression of iNOS was analyzed by PCR. Four hours after the induction of pancreatitis (Figure 1A), no iNOS mRNA expression was detected in the control or HS groups, but expression was significantly elevated in the NT group (P<0.01 vs. HS and control). In the NS group, iNOS mRNA expression was noticeable but lower than that seen in the NT group, (although this result was not significant). Twelve and twenty-four hours after the induction of pancreatitis, no iNOS mRNA expression was observed in any of the groups.
, as assessed by PCR. iNOS mRNA expression was observed only at 4 h after the induction of pancreatitis. Representative mRNA iNOS (170-bp) and 18S rRNA (320-bp) bands (1B). Data are means ± SEM, n = 5 in each group. *P<0.05 vs. C and HS.")
Densitometric analysis of iNOS gene expression (1A), as assessed by PCR. iNOS mRNA expression was observed only at 4 h after the induction of pancreatitis. Representative mRNA iNOS (170-bp) and 18S rRNA (320-bp) bands (1B). Data are means ± SEM, n = 5 in each group. *P<0.05 vs. C and HS.
The levels of nitrite and nitrate in the liver are shown in Figure 2. Four hours after the induction of pancreatitis, no significant differences were observed between any of the experimental groups and the control group in terms of nitrite and nitrate levels in the liver samples. Twelve hours after the induction of pancreatitis, nitrite and nitrate levels were significantly elevated in the NS group (P<0.05 vs. NT; P<0.01 vs. control). However, 24 h after the induction of pancreatitis, nitrite and nitrate levels were slightly lower in the NS group and were significantly higher in the HS group than in the control group (P<0.01).
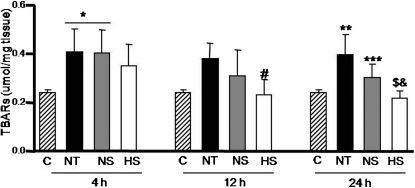
Lipid peroxidation, NS, HS, and NT groups. Data are means ± SEM, n = 5 in each group. *P<0.01 vs. C; $P<0.05 vs. NT; #P<0.05 vs. NS.")
Figure 3 shows that 4 h after the induction of pancreatitis, lipid peroxidation was significantly increased in the NT and NS groups (P<0.05 vs. control) but not in the HS group. Twelve and twenty-four hours after the induction of pancreatitis, lipid peroxidation remained lower in the HS group (P<0.05 vs. NT at 12 h; P<0.05 vs. NS at 24 h; P<0.001 vs. NT at 24 h).
 in tissue homogenates of rats in the control (C), NS, HS, and NT groups. Data are means ± SEM, n = 8 in each group. *P<0.05 vs. C; #P<0.05 vs. NT at 12 h; **P<0.01 vs. C; ***P<0.05 vs. NT at 24 h; $P<0.001 vs. NT at 24 h; &P<0.05 vs. NS at 24 h.")
Effect of hypertonic saline on hepatic lipid peroxidation. Thiobarbituric acid reactive substances (TBARS) in tissue homogenates of rats in the control (C), NS, HS, and NT groups. Data are means ± SEM, n = 8 in each group. *P<0.05 vs. C; #P<0.05 vs. NT at 12 h; **P<0.01 vs. C; ***P<0.05 vs. NT at 24 h; $P<0.001 vs. NT at 24 h; &P<0.05 vs. NS at 24 h.
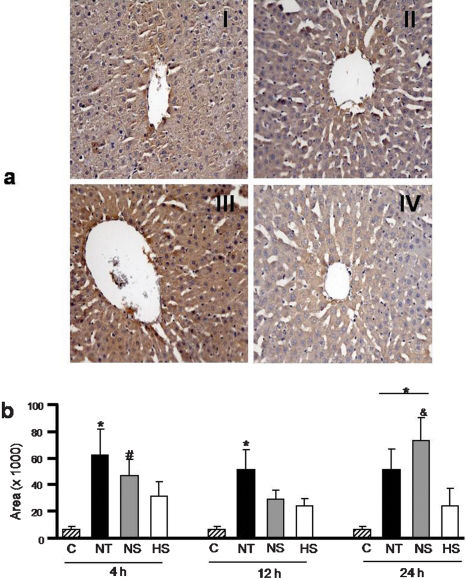
As shown in Figure 4a, cells that stained positive for nitrotyrosine were detected in liver samples collected from rats in the NT, NS, and HS groups. Nitrotyrosine staining was observed around the hepatic central vein. Figure 4b shows that intense expression of hepatic nitrotyrosine was observed in the NS and NT groups 4 h after the induction of pancreatitis (P<0.05 vs. control). Twelve hours after the induction of pancreatitis, nitrotyrosine levels remained high in the NT group, although they were slightly lower in the NS group (in comparison with the levels at 12 h). However, 24 h after the induction of pancreatitis, nitrotyrosine levels were again elevated in the NS group (P<0.05 vs. HS), although they were unchanged in the NT group (P<0.01 vs. control).
 from rats in the four groups: control (I) NT (II), NS (III) and HS (IV) at 24 h. Nitrotyrosine expression (b) in all groups. Data are the means ± SEM, n = 8 in each group. (Original magnification, ×200). *P<0.01 vs. C; #P<0.05 vs. C; &P<0.05 vs. HS.")
Nitrotyrosine expression in liver samples (a) from rats in the four groups: control (I) NT (II), NS (III) and HS (IV) at 24 h. Nitrotyrosine expression (b) in all groups. Data are the means ± SEM, n = 8 in each group. (Original magnification, ×200). *P<0.01 vs. C; #P<0.05 vs. C; &P<0.05 vs. HS.
Four hours after the induction of pancreatitis, we observed an increase in the plasma levels of ALT in the NT group (P<0.05 vs. control), which is shown in Figure 5. The same was found for the NS group 12 h after the induction of pancreatitis (P<0.05 vs. HS, P<0.05 vs. control). However, plasma ALT levels remained normal in the HS group throughout the 24-h study period.
Hsp70 expression, NS, HS, and NT groups. Data are means ± SEM, n = 6 in each group. *P<0.05 vs. C; #P<0.01 vs. C; &P<0.05 vs. HS.")
Figure 6 shows the hepatic expression of Hsp70. Four hours and twenty-four hours after the induction of pancreatitis, slight divergences were observed among the groups, although none of the differences were statistically significant. However, at 12 h, Hsp70 expression was significantly higher in the NS group than in the control group (P<0.05).
DISCUSSION; and representative film of Hsp70 protein expression (A). (B) β-actin was used as the loading control (100 μg protein/lane). Data are means ± SEM, n = 4 in each group. *P<0.05 vs. C.")
In inflammatory disorders, the liver filters inflammatory mediators from blood that has passed through other abdominal organs.21 Similar to the other organs, the liver is both a source and a target of oxidative stress. In the liver, multiple cell types, including hepatocytes, Kupffer cells, stellate cells, endothelial cells, and infiltrating leukocytes, have the capacity to generate NO, superoxides, and peroxynitrite.22 Therefore, the liver is a target of the oxidative stress that occurs in the early stages of pancreatitis-induced systemic inflammation.23,24 In the present study, we found that the infusion of a small volume of hypertonic saline had beneficial effects when used as the sole treatment for pancreatitis-associated hepatic injury.
Fluid resuscitation is a necessary therapeutic intervention in severe pancreatitis. Patients with pancreatitis present with volume extravasation to the peritoneum and retroperitoneum, and some have hemodynamic instability. However, the infusion of large volumes can induce pulmonary interstitial edema and can increase intra-abdominal pressure. Fluid accumulation in the lungs exacerbates respiratory failure and can make mechanical ventilation necessary. Increased abdominal pressure reduces the venous return to the heart, which decreases cardiac output and reduces perfusion of the kidney and gut and can provoke organ damage.25 These data demonstrate the relevance of studying the infusion of small volumes to improve cardiac output and organ perfusion.
Pancreatitis is known to cause an increase in superoxide production as determined by the degree of lipid peroxidation, which was measured in our study. Four and twelve hours after pancreatitis induction, no differences were observed between the NS and NT groups in terms of the degree of lipid peroxidation in the liver. However, hypertonic saline resuscitation was highly effective in reducing lipid peroxidation during pancreatitis and protected against the pancreatitis-induced increase in iNOS expression and the consequent elevation of plasma NO levels. The key players in the physiology of inflammatory processes are NO and its products.26 The literature is unclear as to whether the effects of NO are protective or harmful.10 Evidence suggests that NO promotes hepatocyte death under conditions of oxidative stress, indicating concurrent superoxide production. However, under resting conditions, NO is hepatoprotective.10,27 One interesting finding of the present study is that in the HS group, lipid peroxidation decreased between post-pancreatitis induction hours 4 and 12 and slightly increased at hour 24, even though it remained generally low over the 24-h study period. Hypertonic saline resuscitation produced an environment without superoxides and allowed a late-phase increase in NO; this process exerted a hepatoprotective effect. The effects of hypertonic saline have been shown to persist up to 18 h after administration;28 this therapeutic window may be extended to 24 h based on the results of the current study. In fact, it has been shown that the benefits of hypertonic saline are chronically sustained.29 Although the effects of hypertonic saline are rapid and transient, we can hypothesize that these initial effects on the inflammatory response can affect later pathological development, which has been previously suggested.27
In addition to the known, direct positive effect of hypertonic saline (protection via the modulation of NO, reduction of superoxide levels and modulation of interleukin activation30), there is an indirect benefit. In the present study, hypertonic saline treatment reduced the spontaneous formation of peroxynitrite from NO and superoxide. In the presence of high superoxide levels, peroxynitrite formation has been shown to become dependent on NO production.9 Peroxynitrite can exert its toxic effect through the nitration of macromolecules31 or as a selective oxidant,32 contributing to either necrosis or apoptosis.33 The formation of nitrotyrosine is a consequence of peroxynitrite activity, and increased nitrotyrosine levels have been detected in human diseases associated with oxidative stress.34 In the current study, a marked increase in nitrated proteins was observed in the liver 24 h after the induction of pancreatitis. Staining for nitrotyrosine was significantly greater in the NS and NT groups than in the control group, while it was significantly lower in the HS group than in the NS group. There are various ways in which peroxynitrite-induced impairment of endothelial function might contribute to the pathogenesis of organ failure due to circulatory shock: by exacerbating local vasospasm, increasing local neutrophil adhesion, and increasing neutrophil migration into inflamed tissues; by exacerbating platelet activation and aggregation; or by promoting hypoperfusion of certain parts of various organs. In our experiments, the hepatocytes around the central vein were apparently the most susceptible to peroxynitrite damage (most pronounced staining for nitrotyrosine). The superoxide anions and NO both reacted rapidly to form the toxic reaction product, i.e., the peroxynitrite anion.35 Pharmacological studies in a variety of experimental systems have demonstrated that peroxynitrite is more cytotoxic than NO or superoxide,36,37 and it is able to induce necrosis and apoptosis.38 Peroxynitrite (endogenous or exogenous) is a potent trigger of DNA single-strand breakage, but NO is not.38,8 Subsequently, DNA single-strand breakage can activate the nuclear enzyme poly(adenosine diphosphate-ribose) polymerase.39
The concentrations of NO and superoxide determine the formation of peroxynitrite.40 In our study, the production of NO in the NT group at 12 and 24 h after the induction of pancreatitis was similar to that seen in the control and HS groups at the same time points. This might be because peroxynitrite formation consumes NO, which was shown by the accumulation of nitrotyrosine 24 h after the induction of pancreatitis. Despite the increased NO production observed in the NS group 12 h after the induction of pancreatitis, nitrotyrosine accumulation did not occur until 24 h, which indicates that resuscitation with normal saline (NaCl 0.9%) retards the formation of nitrotyrosine. Although resuscitation with normal saline reduced superoxide production 12 h after the induction of pancreatitis, this effect was not sustained; elevated production of superoxides and NO, which favored peroxynitrite formation, was again observed at 24 h. However, in the HS group, no increase in nitrotyrosine levels was observed at any of the time points evaluated. The protective effect of hypertonic saline was related to the establishment of a superoxide-NO balance that was unfavorable to nitrotyrosine formation. When superoxide levels are low, the regular effects of NO, such as the control of blood flow and inhibition of leukocyte adhesion, predominate.41
We found high ALT levels, which indicate hepatic cellular damage, in the NS and NT groups, whereas ALT levels in the HS group were comparable to those observed in the control group. Any disturbance in membrane permeability, structure, or fluidity will cause a translocation of liver enzymes to the blood.42 In the current study, the release of cytosolic enzymes (ALT) into the serum was attributed to plasma membrane damage, which is primarily caused by lipid peroxidation. Although ALT levels in the NS and HS groups were not as high as those seen in the NT group, they were above normal, indicating hepatic injury. The damage caused by pancreatitis is self-limited and results in elevated plasma levels of ALT for a short time. However, TBARS and nitrotyrosine levels can remain elevated for longer periods depending on the rate of depuration of lipids and proteins from tissues. These conditions can preclude the establishment of a direct clinical time-to-time correlation of the events. In addition, we found that the expression of Hsp70 was elevated in the NS group, which also exhibited a peak in the inflammatory profile at 12 h. Machado et al.12 also demonstrated this peak at 12 h after the induction of pancreatitis and reported elevated plasma levels of cytokines and hepatic enzymes in animals treated with normal saline or receiving no treatment. Similar to our study, no such peak was observed in the animals treated with hypertonic saline in that study. Heat-shock proteins are produced in response to stress and are regulated by heat-shock factors, which are inactive under conditions without stress.43 Due to the hepatoprotection conferred by hypertonic saline administration in the early stages of pancreatitis, heat-shock protein expression is not initially activated. In animal models of pancreatitis, hypertonic saline resuscitation has been reported to protect the liver against the uncontrolled increase of protein expression and activity.14,15
Despite the benefits of reduced oxidative stress with administration of hypertonic compared to normal saline, we did not observe any difference among the C, NT, and HS groups with respect to nitrite and nitrate production, ALT levels, and HSP70 expression at 12 h. This finding can be explained by the sustained ability of the hepatic antioxidant defense to maintain the redox balance in the HS group.44,45 However, cell activation, induced by reactive oxygen species, can be reduced after intense damage46, which includes increased ALT, nitrite and nitrate levels as well as lipid peroxidation and was noted in the NT group during the first 4 h. Furthermore, reactive oxygen species may antagonize reactive nitrogen species during liver injury and inflammation. Although few studies have examined groups submitted to AP or hepatic damage without treatment, we hypothesized that the intense, rapid, and transient hepatic oxidative stress during pancreatitis leads to local inflammation and could increase morbidity and mortality. However, the effect of the hypertonicity in the intracellular signalling cascades can be explained by the anti-oxidative activity. Also, hypertonic saline diminishes the inflammation, and this could contribute to maintenance of the redox balance.
CONCLUSIONSIn summary, our findings suggest that hypertonic saline reduces oxidative stress by modulating NO production, lipid peroxidation, and peroxynitrite formation, thereby minimizing liver damage during pancreatitis. Our results support the use of hypertonic saline as a therapeutic approach to prevent distant organ injury in critical illness, such as acute pancreatitis.
The authors are grateful for the financial support provided by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, Foundation for the Support of Research in the State of São Paulo).