



Activating mutations in exon 3 of the β-catenin gene are involved in the pathogenesis of adamantinomatous craniopharyngiomas. Recently, the interaction between β-catenin and PROP1 has been shown to be responsible for pituitary cell lineage determination. We hypothesized that dysregulated PROP1 expression could also be involved in the pathogenesis of craniopharyngiomas.
OBJECTIVES:To determine whether dysregulated gene expression was responsible for tumor pathogenesis in adamantinomatous craniopharyngiomas, the β-catenin gene was screened for mutations, and the expression of the β-catenin gene and PROP1 was evaluated.
METHODS:The β-catenin gene was amplified and sequenced from 14 samples of adamantinomatous craniopharyngiomas. PROP1 and β-catenin gene expression was assessed by real-time RT-PCR from 12 samples, and β-catenin immunohistochemistry was performed on 11 samples.
RESULTS:Mutations in the β-catenin gene were identified in 64% of the adamantinomatous craniopharyngiomas samples. Evidence of β-catenin gene overexpression was found in 71% of the tumors with β-catenin mutations and in 40% of the tumors without mutations, and β-catenin immunohistochemistry revealed a nuclear staining pattern for each of the analyzed samples. PROP1 expression was undetectable in all of the tumor samples.
CONCLUSION:We found evidence of β-catenin gene overexpression in the majority of adamantinomatous craniopharyngiomas, and we also detected a nuclear β-catenin staining pattern regardless of the presence of a β-catenin gene mutation. These results suggest that WNT signaling activation plays an important role in the pathogenesis of adamantinomatous craniopharyngiomas. Additionally, this study was the first to evaluate PROP1 expression in adamantinomatous craniopharyngiomas, and the absence of PROP1 expression indicates that this gene is not involved in the pathogenesis of this tumor, at least in this cohort.
Craniopharyngiomas are rare intra/suprasellar neoplasms that account for 10% of all pediatric intracranial tumors. During childhood, they are the most common lesions involving the hypothalamo-pituitary regions.1 Craniopharyngiomas are histologically benign and can be categorized into two primary subtypes with different histopathological features: adamantinomatous and papillary. The former is thought to arise from squamous cell rests of the craniopharyngeal duct, which is a canal that connects the stomodeal ectoderm with Rathke's pouch,2,3 and the latter subtype results from metaplasia of the adenohypophyseal cells in the pituitary gland.4 The adamantinomatous craniopharyngioma is the most common histological type and occurs at all ages, but it predominantly affects individuals in the first two decades of life. Conversely, papillary craniopharyngiomas have been almost exclusively described in adults.5
The pathogenesis of craniopharyngiomas is poorly understood. To date, mutations in the β-catenin gene have been identified only in the adamantinomatous subtype and at a variable frequency.6–9 The β-catenin protein is encoded by the CTNNB1 gene. The association of β-catenin with cadherin adhesion molecules at the plasma membrane provides a link between cadherins and the actin cytoskeleton, which is essential for strong cell-cell adhesion.10 Furthermore, β-catenin is a key mediator of the canonical WNT signaling pathway, which is involved in the control of stem cell pluripotency, cell proliferation, differentiation and migration.11 β-catenin is regulated by a multiprotein complex that contains the tumor suppressor protein adenomatous polyposis coli (APC), the scaffold proteins Axin and Axin2 and the phosphokinases glycogen synthase kinase-3β (GSK-3β) and casein kinase 1. When WNT signaling is inactive, β-catenin is phosphorylated within the multiprotein complex; this phosphorylation results in its ubiquitination and degradation by the proteasome.12 However, WNT activation inhibits GSK-3β, and β-catenin is then able to avoid degradation and accumulate in the cytoplasm. It is then translocated into the nucleus where it binds to the transcription factor T cell factor (TCF)/lymphoid enhancer factor (LEF) and thereby regulates gene expression.13 The GSK-3β-binding domain of β-catenin corresponds to its degradation targeting box and is encoded by exon 3 of CTNNB1. Activating mutations within this region, which have been described for the adamantinomatous subtype but not the papillary subtype,6–9 promote β-catenin accumulation through the inhibition of its degradation and lead to WNT signaling activation.14
Recently, the interaction between β-catenin and PROP1 was described as a new mechanism for the β-catenin-dependent regulation of pituitary cell lineage determination in response to WNT signaling. According to this model, the PROP1/β-catenin protein complex would act as a binary switch to simultaneously repress the transcription factor HESX1 and activate the expression of the transcription factor PIT1.15 Interestingly, both factors are involved in pituitary cell proliferation and differentiation.
PROP1 is a paired-like homeodomain transcription factor that is essential for pituitary development and function due to its regulation of PIT1 expression,16 which is essential for the terminal differentiation and expansion of somatotropes, lactotropes and thyrotropes.17 PROP1 is encoded by PROP1, a homeobox gene with 3 exons. The homeodomain is characterized by three helices, the third of which is essential for target gene DNA recognition.18 PROP1 can bind to DNA via the carboxy-terminus to activate target genes or via the amino-terminus and the homeodomain to repress target genes. These results suggest that PROP1 can act as both a transcriptional activator and repressor.16,19
Patients with loss-of-function mutations in PROP1 present with combined pituitary hormonal deficiencies that are generally associated with pituitary enlargement.20–25 Magnetic resonance imaging of the sellar regions of these patients can demonstrate a T1 hyperintense signal that resembles craniopharyngioma-like tumors.
In αGSU-Prop1 transgenic mice, persistent Prop1 expression may result in increased cellular proliferation, which would enhance the probability of a tumor-initiating mutation. In fact, these mice exhibit defects consistent with the dysregulation of pituitary cell proliferation, such as adenomatous hyperplasia with the formation of Rathke's cleft cysts and tumors. The phenotype of these mice suggests that dysregulation of PROP1 expression in humans may contribute to the pathogenesis of pituitary tumors.26
Morbidity and mortality frequently result from the aggressive nature of craniopharyngiomas as well as their treatment, which lead to significant medical and social problems. Therefore, it is critically important to better elucidate the pathogenesis of this tumor. Thus, the aim of this study was to analyze PROP1 and CTNNB1 expression in adamantinomatous craniopharyngiomas and correlate these findings with the presence of somatic mutations in CTNNB1.
MATERIALS AND METHODSSubjectsThe study was approved by the ethics committee of the Hospital das Clínicas in São Paulo, Brazil, and informed written consent was obtained from all of the patients' parents. We analyzed 14 adamantinomatous craniopharyngiomas that were obtained during the surgical resections of children and adolescents (five males and nine females). The age of these patients ranged from 1.58 to 21.58 years (mean 10 years). The phenotype of these patients was very similar. Most patients presented with headaches, visual deficiencies, hypopituitarism and large tumors, and most patients required two or more surgical procedures for the relief of their neurological symptoms. All samples were used for cDNA sequencing. However, two of the 14 tumor samples were excluded from the expression analysis due to their poor RNA quality. β-catenin immunohistochemistry was performed on ten samples.
RNA extraction and reverse transcriptionTotal RNA was extracted from fresh-frozen tissues, which had been preserved in RNAlater® (Ambion, Austin, TX USA) at -20°C using the TRIzol® reagent (Invitrogen, Carlsbad, CA, USA). The elapsed time between tissue isolation and RNA extraction was approximately one to two weeks. The integrity of the extracted RNA was evaluated on a 1% agarose gel, and the concentrations were determined using a spectrophotometer (Eppendorf Biophotometer, Hamburg, Germany). The cDNA samples were obtained from a reverse transcription reaction performed on the same day as RNA extraction and were stored at -20°C. The cDNA was synthesized from 1.25 μg of total RNA in a final volume of 50 μL, and a final concentration of 25 ng/μL was obtained using the high-capacity cDNA archive kit (Applied Biosystems, Foster City, CA, USA). The reverse transcription reaction was performed at 25°C for 10 minutes and was followed by a cycle at 37°C for 120 minutes. Next, the remaining RNA was stored at -80°C. The molecular analyses were most often conducted within two weeks of cDNA synthesis.
PCRExon 3 of CTNNB1 (ENSG00000168036) was amplified using the primers sense 5′-GATTTGATGGAGTTGGACATGG-3′ and antisense 5′-TGTTCTTGAGTGAAGGACTGAG-3′. The PCR reaction setup was as follows. cDNA (25 ng) from each tumor sample was added to a 25-μL reaction mixture containing 5x PCR buffer (50 mM KCL, 1.5 mM MgCl2 and 10 mM Tris-HCL pH 9.0), 200 μmol of dNTPs, 20 pmol of each primer and 1.5 U of GoTaq® DNA Polymerase (Promega, Madison, WI, USA). The PCR reaction mixture was then denatured for 5 minutes at 94°C, cycled 35 times (94°C for 45 sec, 65°C for 45 sec and 72°C for 60 sec) and incubated at 72°C for 10 minutes for a final extension. The PCR reaction yielded a fragment of 218 bp. Each of the amplified fragments was examined using 1.5% agarose gel electrophoresis, stained by ethidium bromide and visualized under UV light. The PCR products were pretreated with an enzymatic combination of exonuclease I and shrimp alkaline phosphatase (United Stated Biochemical Corp., Cleveland, OH, USA) and were directly sequenced using the BigDye terminator cycle sequencing ready reaction kit (PE Applied Biosystems, Foster City, CA) and an ABI PRISM 3100 automatic sequencer (Perkin-Elmer Corp., Waltham, MA, USA).
Quantitative real-time RT-PCRThe PROP1, PIT1 and CTNNB1 mRNA levels were measured by quantitative real-time RT-PCR from 12 adamantinomatous craniopharyngiomas using the ABI Prism 7000 sequence detection system (Applied Biosystems, Foster City, CA, USA). TaqMan® gene expression assays were used to quantify gene expression (PROP1 - Hs00196604_m1; PIT1 - Hs00230821_m1; CTNNB1 - Hs00355045_m1). The amplification reactions were performed in triplicate using 12.5 μL of TaqMan® universal PCR master mix (Applied Biosystems), 2 μL of cDNA (25 ng/μL) and 1.25 μL of TaqMan® gene expression assay solution in a final volume of 25 μL. The geNorm software was used to identify the most adequate internal control gene for the normalization of target gene expression.27 The three most stably expressed housekeeping genes in adamantinomatous craniopharyngiomas were PGK1 (phosphoglyceratekinase 1 - 4326318E), PPIA (cyclophilin A - 4326316E) and HPRT1 (hypoxanthine phosphoribosyltransferase 1 - 4326321E). The geometric mean of the expression of these three genes was used as the internal control gene value. A pooled sample of normal pituitary tissue from 39 adult patients (Human Pituitary Gland Poly A+ RNA, Clontech, Palo Alto, CA, USA) was used as a reference for relative quantification using the 2-ΔΔCT method.28 A no template control (NTC) sample was used in each amplification plate to exclude the possibility of contamination. PIT1 expression was also evaluated to exclude the presence of normal pituitary gene expression in the craniopharyngioma samples. PIT1 is a transcription factor that is a member of the POU domain family, which is essential for the proliferation and differentiation of the somatotrope, lactotrope and thyrotrope pituitary lineages.29 Samples were considered as overexpressed when they demonstrated a two-fold or greater expression change over the normal pooled pituitary samples. Undetectable expression was defined as a CT value greater than 34 due to high variability and poor precision of the relative quantification from the tumor samples above this CT value.
ImmunohistochemistryImmunohistochemical analyses were performed on 3-μm paraffin sections that were deparaffinized in xylene and rehydrated through a series of ethanol treatments. Antigen retrieval was accomplished by incubating the sections in 10 mM sodium citrate pH 6.0 and heating them with a steamer. To block endogenous peroxidase activity, sections were immersed for 30 minutes in a 6% hydrogen peroxide solution diluted in methanol (v/v) and were then washed with phosphate buffer (10 mM phosphate buffered saline (PBS) pH 7.4). Proteins were blocked using CAS-Block (Zymed). For the detection of β-catenin, monoclonal antibody (clone 14, BD) was used at a 1:800 dilution, and the stained cells were incubated at 37°C for 30 minutes and then at 4°C overnight. The final visualization was carried out by chromogenic detection using a solution of PBS containing 0.1% diaminobenzidine, 0.06% H2O2 and 1% dimethyl sulfoxide. The sections were immersed in this solution for 5 minutes at 37°C. Counterstaining was performed using Harris hematoxylin. A known positive control was included, and sections that were incubated with PBS instead of the primary antibody served as negative controls.
Statistical analysisAll statistical analyses were performed with SPSS version 16.0 (SPSS, Chicago, IL). Categorical variables were compared using Fisher's exact test, and p values <0.05 were considered significant.
RESULTSHeterozygous missense mutations in exon 3 of CTNNB1 were found in 9 of the 14 adamantinomatous craniopharyngiomas within codons 32, 33, 37 and 41, which affect the target region of GSK-3β (Table 1). All but two of these mutations have been previously described in craniopharyngiomas.6–9 One of these novel mutations was the result of an amino acid substitution of aspartic acid, a hydrophilic amino acid, for valine, a hydrophobic amino acid, at codon 32. The other novel mutation was a serine to proline replacement at residue 37; both of these amino acids are classified as polar and uncharged.
Somatic mutations in exon 3 of CTNNB1 in patients with adamantinomatous craniopharyngiomas.
| Patients | Age (years) | Sex | Coding DNA sequence change | Amino acid change |
|---|---|---|---|---|
| 1 | 9.66 | M | c.122C>T | p.Thr41Ile |
| 2 | 4 | M | Not found | Not found |
| 3 | 1.58 | F | c.110C>T | p.Ser37Phe |
| 4 | 10.58 | F | Not found | Not found |
| 5 | 9.9 | M | c.109T>C | p.Ser37Pro1 |
| 6 | 21 | M | Not found | Not found |
| 7 | 2.41 | F | c.94G>A | p.Asp32Phe |
| 8 | 1.83 | M | c.122C>T | p.Thr41Ile |
| 9 | 13.58 | F | c.98C>T | p.Ser33Phe |
| 10 | 17.58 | F | c.94G>A | p.Asp32Phe |
| 11 | 14.5 | F | c.98C>G | p.Ser33Cys |
| 12 | 11.66 | F | Not found | Not found |
| 13 | 9.25 | F | Not found | Not found |
| 14 | 4 | F | c.95A>T | p.Asp32Val1 |
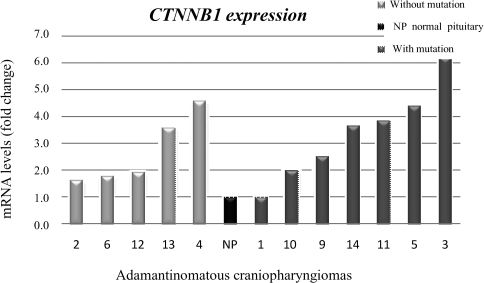
CTNNB1 overexpression was found in five of the seven samples with β-catenin mutations (tumor samples 3, 5, 9, 11, and 14), and this expression was found to be between 2.5- and 6.2-fold greater in these samples than in those from the normal pituitary. Of the five samples without β-catenin mutations, two were found to have upregulated CTNNB1 expression (fold change of 4.6 and 3.6 for samples 4 and 13, respectively) (Figure 1). However, this difference did not reach statistical significance (p = 0.558).

The PROP1 mRNA levels were undetectable in 10 of 12 samples. Two of these samples showed both PROP1 and PIT1 expression, which indicated contamination from normal pituitary tissue. Therefore, these samples were discarded from the analysis.
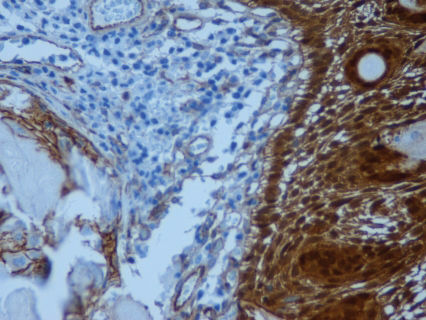
Immunohistochemical staining for β-catenin expression showed a nuclear staining pattern (Figure 2) in all the analyzed samples, and this same result was also observed for those samples without CTNNB1 mutations (Table 2).

Immunohistochemical staining for β-catenin expression, the mutational analysis and the expression of CTNNB1 in adamantinomatous craniopharyngiomas.
| Patients | Nuclear staining | CTNNB1 mutation | CTNNB1 expression (fold number) |
|---|---|---|---|
| 1 | ++ | Yes | 0.97 |
| 2 | Not performed | No | 1.6 |
| 3 | +++ | Yes | 6.2 |
| 4 | + | No | 4.5 |
| 5 | Not performed | Yes | 4.4 |
| 6 | Not performed | No | 1.7 |
| 7 | + | Yes | 3.3 |
| 8 | ++ | Yes | 2.7 |
| 9 | + | Yes | 2.5 |
| 10 | + | Yes | 1.9 |
| 11 | ++ | Yes | 3.8 |
| 12 | + | No | 1.9 |
| 13 | ++ | No | 3.5 |
| 14 | Not performed | Yes | 3.6 |
Nuclear staining was classified as follows: +, staining in a maximum of 10% of the cells; ++, staining in 10% to 50% of the cells; +++, staining in more than 50% of the cells. Expression is represented by the fold change compared to the normal pituitary pool.
Craniopharyngiomas are the most common lesions of the hypothalamo-pituitary region in the child population.1 Despite their benign histological appearance, the aggressive behavior and the invasiveness of these tumors into pituitary and parasellar structures, such as the optic nerve and the hypothalamus, can result in significant neurological sequelae and consequently may impair the quality of life.
Mutations in exon 3 of CTNNB, which encodes the GSK-3β binding domain of β-catenin, have been previously shown to occur in 16% to 100% of adamantinomatous craniopharyngiomas.6–9 In our cohort, CTNNB1 mutations were identified in 64% of the adamantinomatous craniopharyngiomas. These mutations were found at codon 32 (n = 3), which flanks the phosphorylation site of GSK-3β, as well as codons 33 (n = 1), 37 (n = 3) and 41 (n = 2), which affect the serine/threonine residues of β-catenin that are the target of GSK-3β.14 Recently, mutations in these codons were shown to be responsible for increased β-catenin target gene activation, and this increase was represented by elevated mRNA levels of Axin2 and BMP4 in craniopharyngioma cells with nuclear β-catenin accumulation.30Axin2 is a well-recognized inhibitor and target gene of β-catenin31, whereas BMP4 plays a crucial role in tooth development and enhances cell proliferation in tumors with β-catenin accumulation.32,33
Interestingly, 71% of the tumors (five out of seven) with β-catenin somatic mutations were shown to overexpress CTNNB1, and this result was also observed in 40% of the tumor samples without β-catenin mutations (two out of five). Each of the ten samples (seven with CTNNB1 mutations) submitted for immunohistochemical analysis demonstrated nuclear β-catenin staining (Table 2), whereas two out of the three non-mutated samples showed both CTNNB1 overexpression and nuclear β-catenin staining. These results suggested that CTNNB1 overexpression, which leads to WNT signaling activation, plays a key role in the tumorigenesis of adamantinomatous craniopharyngiomas regardless of the presence of CTNNB1 mutations.
The pathogenesis of adamantinomatous craniopharyngiomas that do not have mutations in or display the overexpression of CTNNB1 remains to be determined. To investigate other pathways that are likely involved in this process, we performed a PROP1 expression analysis. This gene was chosen based on the fact that β-catenin, which is the only protein shown thus far to be involved in this pathogenesis, interacts with PROP1 to control pituitary cell lineage determination15, as well as the fact that transgenic mice engineered to exhibit persistent expression of Prop1 demonstrate both defects in the control of cellular proliferation and an increased susceptibility to pituitary tumors.26 Moreover, patients with loss-of-function PROP1 mutations can present with pituitary enlargement.20–25 Thus, we investigated whether PROP1 dysregulation could contribute to the tumorigenesis of adamantinomatous craniopharyngiomas. However, PROP1 expression was undetectable in all the samples, which indicated that expression of the PROP1 gene most likely does not affect adamantinomatous craniopharyngioma pathogenesis.
In conclusion, CTNNB1 overexpression was found in the majority of the adamantinomatous craniopharyngiomas studied. Our findings also suggest that WNT signaling activation via β-catenin upregulation plays an important role in the pathogenesis of adamantinomatous craniopharyngiomas regardless of the presence of CTNNB1 mutations. Additionally, PROP1 dysregulation was not likely involved in the pathogenesis of adamantinomatous craniopharyngiomas in this cohort of patients. However, this study was the first to analyze PROP1 expression in this type of tumor, and further studies are necessary to confirm the lack of a role for PROP1 in this tumorigenesis.
No potential conflict of interest was reported.
Cani CMG is the first author who wrote the paper and was responsible for the collection of the craniopharyngioma samples, application of the methodology, and analysis of the results. Matushita H was the neurosurgeon responsible for collecting the adamantinomatous craniopharyngioma samples during the therapeutic surgery. Carvalho LRS, Brito LP, and Almeida MQ developed the methodology. Soares IC performed the immunohistochemistry. Mendonça BB supervised the study and instructed the group.