




Introducción
A diferencia de otros géneros de estreptococos, S. pyogenes es universalmente sensible a penicilina y mantiene una elevada tasa de sensibilidad frente a cefalosporinas 1.
En los últimos años la resistencia de S. pyogenes a macrólidos se ha incrementado de forma significativa 1,2. El principal mecanismo de resistencia se debe a la presencia de bombas de expulsión activa codificadas por el gen mefA. Éstas confieren resistencia a macrólidos de 14 y 15 átomos, mientras que permanecen sensibles los macrólidos de 16 átomos, las lincosamidas y la estreptogramina B (fenotipo M). Sin embargo, al igual que en otros estreptococos la resistencia a macrólidos puede deberse a la presencia de metilasas codificadas por los genes ermB o ermTR, que confieren resistencia a macrólidos, lincosamidas y estreptogramina B (fenotipo MLSB, constitutivo o inducible) 3.
El objetivo de este estudio es relacionar los resultados de sensibilidad antibiótica de las cepas pediátricas de S. pyogenes aisladas en nuestro hospital los años 2002 y 2003 con los fenotipos de resistencia implicados (M y MLSB) y analizar la evolución de las resistencias a claritromicina y clindamicina desde el año 1996, cuyos datos de sensibilidad fueron parcialmente publicados 4.
Métodos
Entre los años 1996 y 2003 se aislaron, en el Hospital Sant Joan de Déu de Barcelona, 480 cepas (137 en el período 2002-2003) de S. pyogenes procedentes de pacientes de entre 2 meses y 17 años de edad.
Un total de 247 cepas fueron obtenidas mediante frotis faríngeo en pacientes diagnosticados de faringoamigdalitis aguda y el resto de cepas procedían de diversas localizaciones no faríngeas (lesiones cutáneas, abscesos, muestras genitales, secreciones óticas y oculares, líquido articular, líquido pleural, sangre, etc.).
La identificación bacteriana se realizó mediante la determinación de la sensibilidad a bacitracina (BioMérieux, Marcy l'Etoile, Francia) en las cepas de estreptococos β -hemolíticas y posteriormente se confirmó mediante la determinación del serogrupo A (Phadebact® Strep A Test, Boule Diagnostics AB, Huddinge, Suecia). En los casos de cepas resistentes a bacitracina se completó la identificación con el sistema api® 20 STREP (BioMérieux, Marcy l'Etoile, Francia).
La determinación de la sensibilidad antibiótica se realizó por el método de E-test (AB Biodisk, Solna, Suecia). Se estudiaron los siguientes antibióticos: penicilina, claritromicina y clindamicina, y en los aislamientos de los años 2002-2003 se estudió también el linezolid. La sensibilidad antibiótica de linezolid se determinó también por el método de discodifusión (Oxoid, Oxoid Limited, Inglaterra). La interpretación de los resultados se realizó según los criterios del National Committee for Clinical Laboratory Standards (NCCLS).
Como medio de cultivo se utilizó agar Müeller-Hinton suplementado con el 5% de sangre de cordero y la incubación se realizó a 35-37 °C durante 20-24 h en atmósfera aerobia con 5% de CO2.
La caracterización fenotípica de la resistencia a macrólidos se hizo mediante la prueba del doble disco eritromicina (15 μg) y clindamicina (2 μg) 3 en placas de agar Müeller-Hinton con el 5% de sangre de cordero. Además, en las cepas de los años 2002 y 2003 se incluyó el disco de estreptogramina B (25 μg).
La detección de genes de resistencia a macrólidos se realizó mediante la técnica de reacción en cadena de la polimerasa (PCR) a todos los aislamientos resistentes a macrólidos de los años 2002 y 2003 (49 cepas). Para la detección de los genes mefA, ermB y ermTR se utilizaron los cebadores y condiciones descritas por Sutcliffe 5. Se incluyeron controles positivos y negativos en cada serie de amplificación. Los productos de PCR se separaron en geles de agarosa al 1% y tras tinción con bromuro de etidio se visualizaron bajo luz ultravioleta.
La comparación de variables cuantitativas se realizó mediante el test de la chi cuadrado.
Resultados
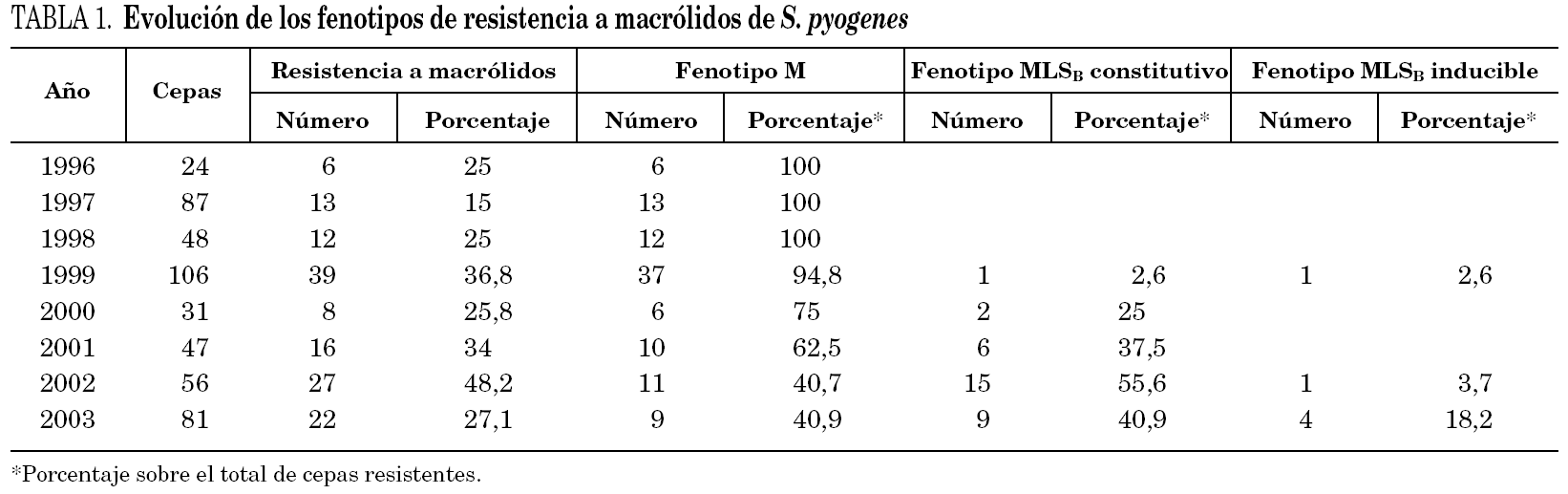
La tabla 1 muestra la evolución de la sensibilidad de S. pyogenes a macrólidos desde el año 1996 al 2003. Se observa una tendencia al aumento de la resistencia hasta alcanzar en el año 2002 un máximo del 48% de cepas resistentes. La resistencia media de estos años fue del 29,8% (27,4% entre 1996 y 2001 y 35,8% entre 2002 y 2003).
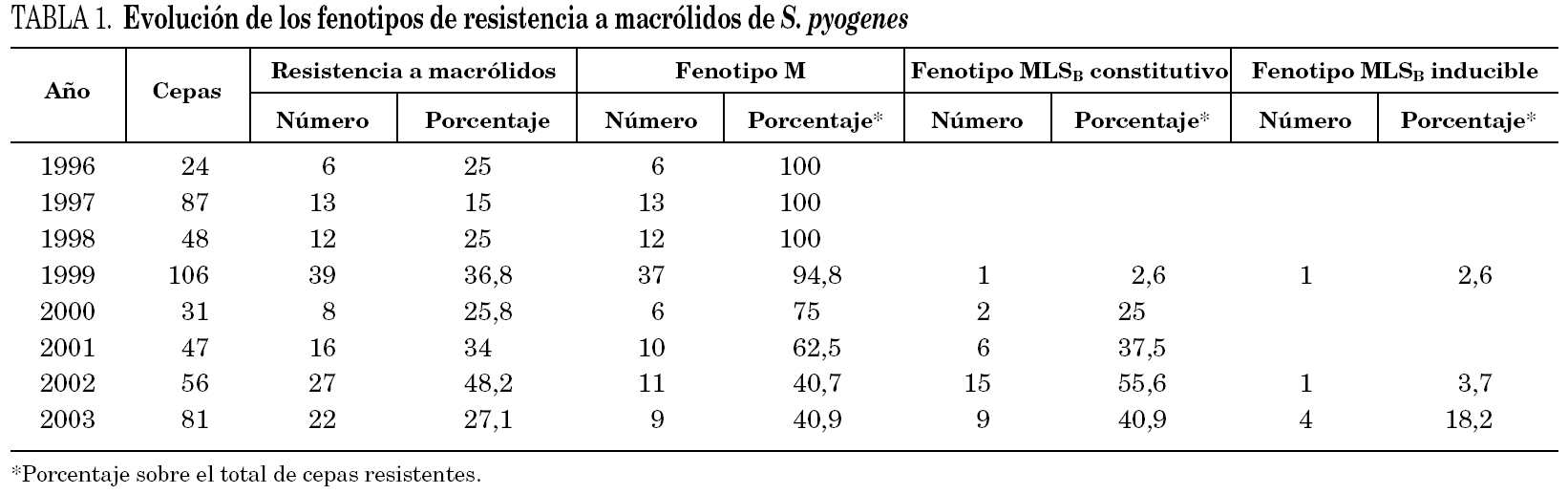
Durante el período de estudio, se ha observado un cambio en los fenotipos de resistencia a macrólidos. En el período 1996-1998 todas las cepas resistentes a claritromicina presentaron el fenotipo M. En 1999 se detectó la primera cepa con fenotipo MLSB constitutivo y en los años siguientes se produjo un importante aumento de cepas con este fenotipo de resistencia, hasta superar el 50% de los aislamientos resistentes a macrólidos en 2002. El aislamiento de cepas con fenotipo MLSB inducible fue anecdótico hasta 2003, en que representó el 18,2% de las cepas resistentes a claritromicina. El aumento del fenotipo MLSB desde el año 1999 es estadísticamente significativo (p < 0,001).
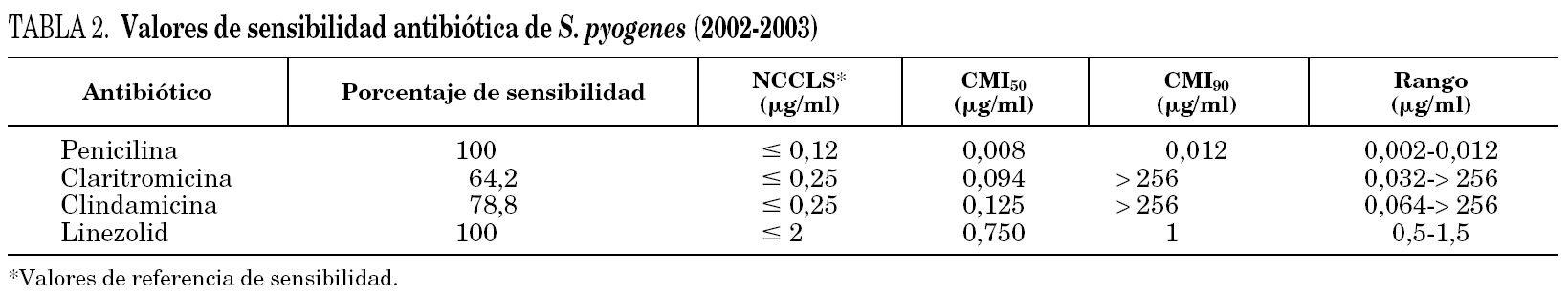
La tabla 2 muestra los valores de sensibilidad de los antibióticos estudiados en el período 2002-2003. La resistencia a claritromicina de las cepas del fenotipo M fue, como es habitual en este fenotipo, de bajo nivel, ya que en ningún caso la concentración inhibitoria mínima (CIM) superó los 32 μg/ml, mientras que clindamicina mantenía el mismo nivel de sensibilidad que las cepas sensibles a macrólidos. En todas las cepas con resistencia constitutiva la CIM de claritromicina y clindamicina fue superior a 256 μg/ml.
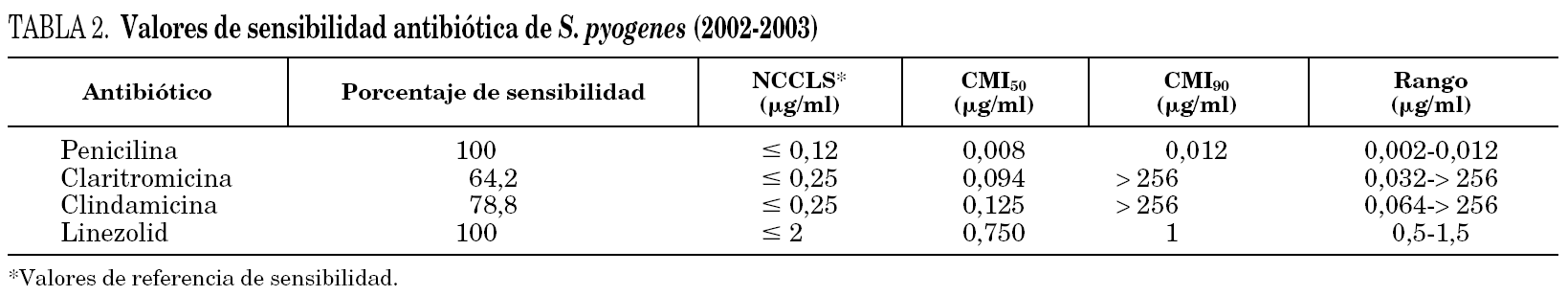
Todas las cepas fueron sensibles a penicilina y linezolid, sin diferencias de CIM entre las cepas sensibles y resistentes a macrólidos. El rango del halo de inhibición del disco de linezolid, obtenido mediante el método de disco difusión, fue de 26-34 mm.
De las cepas aisladas en el período 2002-2003, 8 fueron resistentes a bacitracina (diámetro de inhibición de 0 mm), siete pertenecientes al fenotipo MLSB constitutivo y uno al MLSB inducible.
No se observaron diferencias significativas en cuanto a sensibilidad antibiótica entre las cepas de origen faríngeo y no faríngeo.
En los 20 aislamientos con fenotipo M de resistencia a macrólidos se detectó la presencia del gen mefA y no se detectaron los genes ermB o ermTR. En los 24 aislamientos con fenotipo MLSB constitutivo se detectó la presencia del gen ermB y no se detectaron los genes mefA ni ermTR. De los cinco aislamientos con fenotipo de resistencia MLSB inducible, en dos se detectó la presencia del gen ermB, en uno los genes ermB y mefA y en otros dos el gen ermTR. Todas las cepas con fenotipo M y presencia del gen mefA mostraron halos de inhibición frente a la estreptogramina B superiores a 19 mm. Los aislamientos con la presencia del gen ermB mostraron halos de inhibición con el disco de estreptogramina B inferiores a 16 mm, con independencia del fenotipo (MLSB o MLSB inducible). Las dos cepas que presentaban un fenotipo inducible y en las que se detectó el gen ermTR mostraron halos de estreptogramina B superiores a 19 mm.
Discusión
En España, la resistencia de S. pyogenes a macrólidos se ha relacionado de manera casi exclusiva con cepas del fenotipo M, como resultado de la expresión de la bomba de expulsión activa MefA 1,6. Sin embargo, nuestros datos muestran, un aumento importante de cepas resistentes a macrólidos con fenotipo MLSB, a partir de 1999 mayoritariamente de tipo constitutivo, con presencia predominante del gen ermB, las cuales suponen el 21% del total de aislamientos y el 59% de las cepas resistentes de los últimos 2 años.
Estos resultados son equiparables a los observados en aislamientos pediátricos de S. pyogenes en Francia 7, donde el aumento de cepas con el gen ermB (69,4% de las cepas resistentes a macrólidos) se relaciona con la cepa epidémica emm28 T28. Esta cepa epidémica ha sido también detectada recientemente en algunas zonas de nuestro país, asociada a resistencia a bacitracina 8. Es probable que el incremento de cepas con fenotipo MLSB(ermB) que hemos encontrado se deba en parte a la diseminación de este clon epidémico en nuestra área geográfica.
El fenotipo MLSB inducible en S. pyogenes se ha asociado clásicamente al gen ermTR9. Sin embargo, en nuestro estudio el fenotipo inducible se relaciona indistintamente con los genes ermTR y ermB. Además, había relación entre el halo de inhibición del disco de estreptogramina B y el gen implicado en la resistencia de las cepas MLSB inducibles. Así, las cepas con el gen ermB presentaron halos de inhibición inferiores a 19 mm, mientras que las cepas con el gen ermTR tuvieron halos superiores.
La situación descrita supone un importante cambio en los parámetros habituales de resistencia de S. pyogenes a macrólidos y lincosamidas, al haberse convertido el fenotipo MLSB en predominante en los aislamientos pediátricos en nuestra zona geográfica durante los últimos 2 años estudiados. Un factor concomitante al aumento del fenotipo MLSB es la diseminación de cepas resistentes a bacitracina, que puede dificultar la identificación presuntiva de S. pyogenes.
Los macrólidos de 16 átomos y la clindamicina constituían hasta ahora una alternativa eficaz en el tratamiento de infecciones producidas por cepas resistentes en pacientes alérgicos o intolerantes a betalactámicos, al mantener la actividad frente a cepas del fenotipo M. Sin embargo, el aumento del fenotipo MLSB reduce drásticamente las posibilidades terapéuticas por vía oral.
En el actual contexto, linezolid podría ser una alternativa válida en el tratamiento por vía oral, debido a su buena actividad in vitro10.
Una característica destacable de las infecciones producidas por S. pyogenes, especialmente la faringoamigdalitis aguda, es que habitualmente el diagnóstico se realiza de forma clínica, sin cultivo bacteriano y, por tanto, el tratamiento es empírico. Esta circunstancia, unida a los cambios epidemiológicos descritos en los mecanismos de resistencia y la gran variabilidad geográfica de las tasas de resistencia a macrólidos, hace necesario incrementar la recogida de muestras para cultivo para conocer con mayor precisión la evolución de la sensibilidad antibiótica de este microorganismo y reducir la posibilidad de fracaso terapéutico. En nuestro caso será importante determinar si se mantiene la progresiva implantación del fenotipo MLSB y la disminución relativa del fenotipo M o, por el contrario, se trata de variaciones circunstanciales y geográficamente limitadas.
Correspondencia: Dr. Amadeu Gené.
Servicio de Microbiología. Hospital Sant Joan de Déu.
P.º Sant Joan de Déu, 2. 08950 Esplugues de Llobregat.
Barcelona. España.
Correo electrónico: agene@hsjdbcn.org
Manuscrito recibido el 8-2-2005; aceptado el 1-6-2005.

