




Las cepas verotoxigénicas de Escherichia coli (ECVT), especialmente las del serotipo O157:H7, constituyen un importante grupo de patógenos emergentes para los seres humanos1. Las cepas de ECVT O157:H7 pueden provocar desde una diarrea no sanguinolenta hasta una colitis hemorrágica (CH), con el síndrome urémico hemolítico (SUH) o la púrpura trombocitopénica trombótica como complicaciones más habituales, principalmente en niños, ancianos y pacientes inmunodeficientes2. Su patogenicidad se debe a la producción de una citotoxina conocida como verotoxina o toxina Shiga, de la que existen 2 variantes principales: Stx1 y Stx2. Desde su reconocimiento como patógeno en 1982, el serotipo O157:H7 se ha identificado como causante de innumerables brotes y casos esporádicos de CH y SUH, ampliamente descritos en países anglosajones y Japón. En España, las infecciones por ECVT son relativamente frecuentes, y se han descrito hasta la fecha 7 brotes de CH causados por cepas de ECVT O157:H7, según los datos del Laboratorio de Referencia de E. coli en Lugo3.
Entre los meses de junio de 2006 y diciembre de 2007, en colaboración con el Servicio de Microbiología del Hospital San Pedro de Alcántara de Cáceres, se analizaron para la detección de ECVT O157:H7 1.675 coprocultivos de pacientes internos y externos de todas las edades, aquejados de diarrea y otras alteraciones gastrointestinales. Para ello se llevó a cabo un enriquecimiento previo de las muestras en agua de peptona tamponada suplementada con antibióticos, seguido de separación inmunomagnética con Dynabeads® anti-E. coli O157 (Dynal, Noruega) y siembra del inmunoconcentrado resultante en el medio agar MacConkey sorbitol con telurito y cefixima, de acuerdo con el protocolo para la detección de ECVT en muestras fecales descrito por Rey et al4. A continuación se identificaron los genes que codifican las verotoxinas (stx1 y stx2) y la intimina tipo γ1 (variante eae-γ1, específica de O157:H7) mediante la reacción en cadena de la polimerasa (PCR) a partir de la zona de crecimiento confluente. En los casos positivos se seleccionaron 10 colonias sorbitol-negativas y se sembraron en agar MacConkey lactosa y agar coliformes Chromocult (Merck, Alemania), y se seleccionaron tras la incubación únicamente las colonias lactosa-positivas y beta-glucuronidasa-negativas (colonias de color rosa-rojo en el medio agar coliformes Chromocult). Estas colonias se confirmaron bioquímicamente como E. coli empleando el sistema API 20E (bioMérieux, Francia) y se analizaron individualmente mediante PCR con cebadores específicos para los genes que codifican el antígeno somático O157 (O157 rfbE) y el antígeno flagelar H7 (fliCh7), la intimina (gen eae general y su variante eae-γ1), las verotoxinas y la enterohemolisina (ehxA)5.
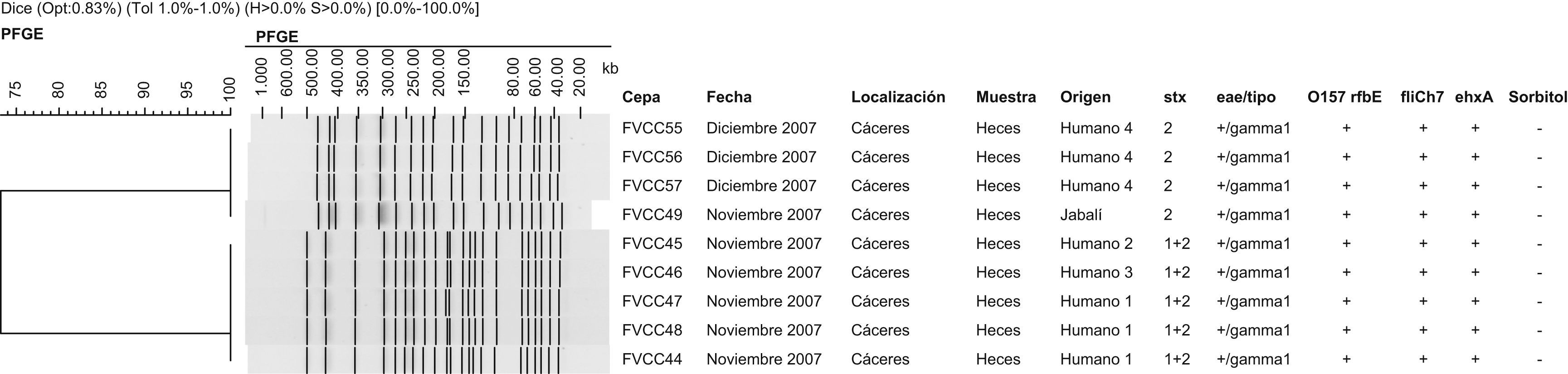
En total se detectaron y se aislaron cepas de ECVT O157:H7 en 8 coprocultivos (0,5%) pertenecientes a 4 pacientes aquejados de procesos gastroentéricos de diversa consideración, concretamente en los meses de noviembre y diciembre de 2007. Todas las cepas mostraron un fenotipo típico (sorbitol-negativas y beta-glucuronidasa-negativas), en todas ellas se identificaron los genes eae (su variante eae-γ1) y ehxA, y se confirmaron los genes O157 rfbE y fliCh7. En 5 cepas se identificaron los genes stx1 y stx2, mientras que en las 3 cepas restantes únicamente se detectó el gen stx2 (fig. 1). En cuanto al patrón de sensibilidad a los antibióticos, las 5 cepas de ECVT O157:H7 stx1 stx2 fueron resistentes a estreptomicina y a sulfamidas, mientras que las 3 cepas de ECVT O157:H7 stx2 fueron sensibles a los 26 antibióticos ensayados. Paralelamente, se obtuvieron los perfiles de electroforesis en campo pulsado (PFGE) de todos los aislados, de acuerdo con el protocolo descrito por Sánchez et al6. Las 5 cepas de ECVT O157:H7 stx1 stx2, aisladas en el mes de noviembre a partir de 3 pacientes pertenecientes a la misma área de salud en un intervalo de aproximadamente 15 días, mostraron perfiles indistinguibles. Las 3 cepas de ECVT O157:H7 stx2, aisladas a partir de un cuarto paciente en el mes de diciembre, mostraron perfiles indistinguibles entre sí, aunque distintos del patrón observado en las 5 cepas anteriores. Al comparar estos perfiles de PFGE obtenidos en cepas de ECVT O157:H7 de origen humano con los de cepas aisladas de animales y de alimentos en estudios previos realizados por nuestro grupo de investigación, el patrón observado en las 3 cepas de ECVT O157:H7 stx2 resultó indistinguible del observado en una cepa de ECVT O157:H7 con el mismo perfil toxigénico aislado a partir de las heces de un jabalí abatido en el mes de noviembre en la misma área de salud (fig. 1).
.")
Aunque no fue posible establecer la fuente de infección en ninguno de los casos, estos resultados ponen de manifiesto el primer brote de infección por ECVT O157:H7 ocurrido en Extremadura y el octavo caso ocurrido en España, y apuntan a un posible papel del jabalí en los casos humanos de infección por ECVT O157:H7 que tienen lugar en la región. Por otro lado, la gravedad de las complicaciones que suelen acompañar a este tipo de infecciones pone en evidencia la necesidad de establecer una vigilancia activa mediante la implantación de protocolos de detección de ECVT O157:H7 en los laboratorios de microbiología clínica de áreas con alta prevalencia del patógeno. En áreas de baja prevalencia, la vigilancia activa debería limitarse al estudio de casos en los que se observe presencia de sangre en heces, brotes de intoxicación alimentaria y casos de pacientes con SUH.
FinanciaciónEste estudio se ha financiado con fondos del III Plan Regional de Investigación, Desarrollo e Innovación de la Junta de Extremadura y FEDER (Ref. Proyecto 3PR05A009).
Nuestro agradecimiento a la Dra. Pilar Teno (Servicio de Microbiología, Hospital San Pedro de Alcántara, Cáceres) por su disponibilidad y su colaboración, y a Raquel Rubio por su inestimable asistencia técnica.

