





Editado por: Andre R Brunoni, Marie-Anne Vanderhasselt, Leigh Chavert
Más datosCompassion motivation is associated with increased heart rate variability (HRV), reflecting a calm and self-soothing physiological state. Recent work, however, suggests that this association is dynamic for the specific components of compassion.
ObjectivesThe present study adopted anodal transcranial direct current stimulation (tDCS) targeting the right insula to see whether this would modulate the sensitivity to suffering and the commitment to engage in helpful actions (i.e., the components of compassion motivation).
MethodNinety-seven healthy individuals underwent 15-min anodal or sham tDCS over the frontotemporal lobe, while watching a video inducing empathic sensitivity and performing a Redistribution Game. Tonic and phasic HRV, dispositional traits, and momentary affects were assessed.
ResultsCompared to sham condition, anodal stimulation favored significant i) HRV reductions during the video and HRV increases during the Redistribution Game; ii) decreases in self-reported levels of negative affect and increases in positive affect during task when the latter was preceded by the video, without influencing altruistic behavior.
ConclusionsAnodal tDCS over the right insula may modulate the engagement phase of compassion by intensifying the psychophysiological sensitivity to signals of distress and protecting from being subjectively overwhelmed by it.
The putative benefits of compassion-based interventions on health have driven a growing interest in the psychophysiological signatures of compassion, defined as “the sensitivity to suffering in self and others, with a commitment to try to alleviate and prevent it” (Gilbert et al., 2017). Within this perspective, recent meta-analytic evidence has found compassion to be positively associated with the fluctuation of instantaneous periodicity of the heart over time, namely heart rate variability (HRV), which is considered a proxy of how top-down appraisal shapes the autonomic response in the body (Di Bello et al., 2020). In the context of interpersonal motivations, higher tonic HRV has been positively associated with mind reading and empathy (e.g., Lischke et al., 2017; 2018), theory of mind (Zammuto et al., 2021), social engagement (e.g., Shahrestani et al., 2015), social connectedness (e.g., Kok & Fredrickson, 2010), prosociality (e.g., Kogan et al., 2014), and cooperative behavior (e.g., Beffara et al., 2016).
Compassion is rooted in social connection and engagement, encompassing both a sensitivity to distress signals of ourselves and others and affiliation-based responsiveness to others (Gilbert, 2019). This approach-related process is dependent upon the dynamic interplay between sympathetic and parasympathetic nervous systems, thus HRV is crucial for compassionate processes such as offering of support, soothing, and comfort (e.g., Kogan et al., 2014). Concomitantly, during states of compassion motivation, prefrontal inhibition of subcortical activation contributes to greater parasympathetic nervous system dominance that, in turn, exerts inhibitory control over sympathetically driven responses at the level of the heart through the vagus nerve, enabling calmness, soothing, and safeness (Porges, 2007; 2017).
Nonetheless, despite the overall positive correlation found between compassion and HRV, recent preliminary evidence suggests that such association is nonlinear. Compassion would be at first related to reduced vagally-mediated HRV, reflecting empathic engagement with suffering, and only at a second stage with increased HRV, when the appropriate helpful action is performed (Di Bello et al., 2021). The present study further examined the putative role of the insular cortex in determining such a physiological and motivational shift.
Non-invasive brain stimulation techniques have been widely used in the field of social interactions with different brain targets (Penton et al., 2022); however, their application to the understanding of compassion is limited. The few existing studies targeted the dorsolateral prefrontal cortex to treat compassion fatigue (Stanton et al., 2015) or to improve the effectiveness of a compassion-based intervention in depressed individuals (Asadinoghabi & Gharibzadeh, 2020). Evidence exists, however, suggesting that the anterior insular cortex (AI) may be the optimal target to stimulate when studying compassion and -even more- its autonomic signatures (e.g., Lutz et al., 2009). The AI is involved in the integration of bodily (interoceptive) states, emotion, and cognition, and it is thought to play a role in the experience of the associated motivational tendencies, rather than in the pure perception of emotional stimuli (e.g., Deng et al., 2021; Wager, 2017). As a critical hub of the salience network, the AI is specifically sensitive to salient environmental stimuli and is responsible for switching between internally (default mode network) and externally focused (central executive network) networks (Menon & Uddin, 2010). By integrating external sensory information with internal emotional and interoceptive information the insular cortex facilitates access to attention and segregates among internal and external personally relevant stimuli to enable the appropriate behavioral response and to modulate homeostatic states. In addition, while the left insula appears to be associated with cognitive-evaluative forms of empathy, the right AI (rAI) may be a more appropriate target as apparently more involved in affective-perceptual forms of empathy and distress (e.g., Li et al., 2020; Uddin et al., 2017).
Overall, these functions of rAI overlap with the empathic sensitivity component of compassion motivation, suggesting a plausible involvement of rAI in the engagement processes. By detecting distress signals and mitigating escape or avoidance action repertoires, the rAI would determine an interoceptive allostatic state. In turn, this facilitates the allocation of attention toward behaviorally important cues (distress), ultimately contributing to the intention (to alleviate it); then, by switching to an executive cognitive state, it would foster the evaluation and monitoring of intentional action decisions and outcomes (helpful actions) (e.g., Brass et al., 2010; Menon & Uddin, 2010; Wager, 2017). According to the evolutionary framework of caring motives, the rAI might be at the core of the care-based algorithm that characterizes compassion: "if there is a signal of distress, then engage in trying to alleviate it" (Gilbert, 2020).
The paucity of studies targeting the rAI with non-invasive brain stimulation techniques is likely due to difficulties in reaching such a deep brain region; however, secondary activations within the insular cortex have been demonstrated by combining brain stimulation and resting-state functional magnetic resonance imaging (Li et al., 2017). It also appeared possible to indirectly increase cortical excitability in the insular cortex by the use of transcranial direct current stimulation (tDCS) (e.g., Petrocchi et al., 2017; Salvo et al., 2021), possibly modulating interoception (Sagliano et al., 2019). TDCS functions through subthreshold modulation of the membrane potentials of neurons, by altering cortical excitability (Nitsche & Paulus, 2000), ultimately changing neuronal synaptic plasticity (Brunoni et al., 2012). This change in neuronal synaptic plasticity is thought to produce Long Term Potentation-like ‘learning’ in the neurons that are modulated. The common assumption is that the anode electrode causes an enhancement of cortical excitability during stimulation, while the cathode electrode generates the opposite effect, even if a more complex interaction of polarity and brain activity than a rigid dichotomy has been suggested (Jacobson et al., 2012).
In light of preliminary evidence suggesting a significant role of emotional saliency in the empathic sensitivity component of compassion (Di Bello et al., 2021; Lutz et al., 2009) and the consistent activation of the salience network in this process (Kim et al., 2020), the present study aimed to determine whether a single session of tDCS over the right frontotemporal lobe to target the insular cortex would be capable of modulating engagement with suffering and, in turn, the aptitude to engage in helpful actions. Given the role of the right insula in integrating interoceptive states, emotions and cognition to generate motivated responses (Menon & Uddin, 2010), we hypothesized that anodal (compared to sham) tDCS over the right insula would 1) enhance attentional sensitivity to others’ distress via phasic changes in psychophysiological reactivity (i.e., reduced HRV) and increased negative affect; and 2) facilitate social engagement with that distress, influencing the response to others’ suffering with increased altruistic behavior, accompanied by increased HRV and reduced negative affect.
MethodParticipantsNinety-seven healthy right-handed individuals aged between 18 and 65 years old, recruited through online announcements, gave written informed consent to participate (Prot. CE/PROG.941). Exclusion criteria were current or past psychiatric, cardiovascular, or neurological disorders; medication intake; pregnancy; body mass index > 30 kg/m2. Four participants were excluded from the analysis due to the bad quality of the physiological recording, leaving a sample of 93 participants (23.98 ± 8.13 years; 76 females).
ProcedureParticipants were asked to abstain from alcohol and heavy physical exercise in the 24 h and not to eat, smoke, or take any stimulants in the 2 h before the session.
Demographic and dispositional information was first assessed, then ECG electrodes were attached. As depicted in Fig. 1, self-reported levels of state affect were given at the beginning of the session and after the beginning of stimulation, video, and task. After filling out the questionnaires, participants sat for 15 min (habituation), during which they colored a mandala to avoid the focus on physical sensations (Daudén Roquet et al., 2021). After the habituation period, participants received the 15-minute active tDCS or sham stimulation, and 2.30 min after the beginning of the stimulation, they were exposed to i) a 2.30-min video representing an “empathic sensitivity” triggering condition (i.e., individuals with a chronic or terminal illness); ii) a recovery period of 2.30 min; iii) the Redistribution Game (Weng et al., 2013) for 10 min; and iv) a second recovery phase of 2.30 min. To minimize any potential carryover effects, the video and the task were randomly presented.
Self-report measures
Sex, age, height, and weight for body mass index calculation, physical activity habits, medical background, and currently prescribed medications were assessed.
The Compassionate Engagement and Action Scales (CEAS; Gilbert et al., 2017) are three measures of compassion competencies assessing the compassion: i) experienced for others; ii) from others; and iii) toward self. Each scale provides the “engagement” (8 items) and “action” (5 items) subscales, and a total score (sum of items). Cronbach's alphas for the present study ranged between 0.75 and 0.93.
The Fears of Compassion Scale (FCS; Gilbert, McEwan, Matos & Rivis, 2011) assesses fears of i) feeling or expressing compassion for others (10 items); ii) receiving compassion from others (13 items); and iii) compassion for self (15 items). Cronbach's alphas ranged between 0.76 and 0.91.
The Interpersonal Reactivity Index (IRI; Albiero et al., 2006; Davies, 1980) is a tool for the multidimensional assessment of empathy. It consists of 28 items divided into four subscales (7 items each): Empathic Concerns, Perspective Taking, Fantasy, and Personal Distress. Cronbach's alpha was 0.90 for the total score.
The Self-Awareness Questionnaire (SAQ; Hughes et al., 2019; Longarzo et al., 2015) consists of 35 items assessing how and how frequently participants feel signals from their own bodies. Cronbach's alpha was 0.91.
The Marlowe-Crowne Social Desirability scale (MC-SDS; Crowne & Marlowe, 1960) is a measure of social desirability bias, defined as the need to respond in culturally sanctioned ways. In the present study, the Italian adaptation (Manganelli Rattazzi et al., 2000) of the short 9‐item version of the MC-SDS was used, with a Cronbach's alpha of 0.95.
As to self-reported levels of state affect, participants were asked to rate how much they felt happy, calm, strong, content, relieved, angry, sad, anxious, self-critical, and weak on a visual analog scale (VAS) from 0 to 10.
Physiological assessmentInter-beat intervals were recorded by using the Bodyguard 2 (Firstbeat) with a standard electrode configuration. HR, time-domain (root mean square of successive differences between normal heartbeats; rMSSD), and frequency-domain (high frequency; HF-HRV) HRV measures, as well as outlier and artifact detection, were analyzed via Kubios software (Tarvainen et al., 2014). Overall, 0.62% of beats were corrected.
Transcranial direct current stimulation protocolA constant current of 2 mA was delivered for 15 min (30 s ramp-up and 30 s ramp-down) by a battery-driven (9v) constant current stimulator (BrainSTIM, EMS s.r.l.), with a maximal output of 10 mA. Conductive rubber electrodes (7 × 5 = 35 cm2) covered by sponges soaked in isotonic saline solution (0.9% w/v of NaCl) were used and held in place by an elastic bandage material. The appropriate amount of saline (6 mL) was obtained using a syringe to avoid the oversaturation of the sponge. An electroconductive gel was applied under the sponges before the montage to reduce skin impedance (< 5 kohms; DaSilva et al., 2011). Differences in skull dimension were dealt with head measurements over participants’ scalps according to the 10–20 System (Klem et al., 1999).
Montage and stimulation parameters were chosen to induce the highest and most widespread electric field amplitude distribution (Nitsche & Paulus, 2000). In both anodal and sham conditions, the anode electrode was placed over the middle point between F8 and T4 and the cathode electrode over the contralateral frontal pole (Fp1). In the sham condition, the current was ramped down after 30 s to ensure that participants felt the same itching sensation (Gandiga et al., 2006). Participants were asked to indicate the condition to which they believed they were assigned to. No adverse effects aside from itching sensations were reported.
VideoParticipants were instructed to watch a 2:30-min video clip in which scenes of chronic or terminally ill patients experiencing suffering were presented wherein they are depicted i) dreading their appointment, ii) too shocked to comprehend treatment options, iii) waiting for a heart donor for transplant surgery, iv) meeting their families for the last time; as in Di Bello et al., 2021).
Redistribution gameThe redistribution game is a decision-making task aimed at assessing altruistic behavior by simulating the unfair treatment of a victim and the costly redistribution of funds to the victim (Weng et al., 2013). Participants observed a programmed dictator (endowed with 90 points) transferring an unfair quantity of points (10 points) to a receiver (endowed with 0 points) through an anonymous (mock) online interaction. Participants (endowed with 50 points) could choose to redistribute any amount (x) of their endowment, and each point redistributed by the participant would result in twice the amount (2x) of points taken from the dictator and transferred to the receiver. Participants were informed that they were playing the game with live participants over the internet. At the end of the experiment, participants were debriefed and asked: “Did you believe you were playing with actual people?” (Yes vs No; Believer Status). Reaction times (RT) and the number of points redistributed by the participants were taken as outcomes.
Data analysisA single-blind, randomized, controlled between-subjects design was used. The analyses were performed using IBM SPSS (v. 27). Normality assessment was performed as a prerequisite for general linear model (GLM). A series of t- and chi-square tests were performed to search for any baseline difference between groups. The variables that emerged to be significantly different between the two groups were included as covariates in the subsequent analyses. Due to the unequal sex distribution in the present sample and given the documented differences between males and females in both rMSSD (Koenig & Thayer, 2016) and compassion tendencies (Yarnell et al., 2019), sex was included as a covariate in all analyses.
To evaluate the effects of tDCS on rMSSD, a GLM with Stimulation type (anodal vs sham) as between-subject and Time (baseline, video, recovery from video, task, recovery from task) as a within-subject factor was specified.
Self-reported levels of state affect were subjected to Principal Component Analysis (PCA) and Varimax orthogonal rotation to extract factors (as in Di Bello et al., 2021). Cattell's (1966) scree test was used as decision rule for identifying the number of factors to retain. Next, the effects of tDCS on affective responses were tested by a series of GLMs with Stimulation type (anodal vs sham) as between-subject factor and Time (Baseline, Video, Task) as a within-subjects on the factor(s) that emerged from the PCA.
Then, the effects of stimulation on altruistic behavior were tested by two univariate GLMs with Stimulation type (anodal vs sham) as a between-subjects factor and the redistribution response value and RTs as outcomes, respectively, controlling for believer status.
For exploratory purposes, the analyses were repeated including scores on the dispositional measures (SAQ, FCS, IRI) as covariates.
ResultsTable 1 shows pre-existing differences between the two groups.
Pre-existing differences between the active and sham groups.
Note. BMI = Body Mass Index; rMSSD = root mean square successive difference; HR = Heart Rate; CEAS = Compassionate Engagement and Action Scales; FCS = Fears of Compassion Scale; SAQ = Self-Awareness Questionnaire; MC-SDS = Marlowe-Crowne Social Desirability scale; IRI = Interpersonal Reactivity Index.
Independent t-tests on baseline state and dispositional measures showed significant differences between groups in Compassion to Other-Engagement subscale (p = .033), with higher scores in the control compared to the experimental group (Table 1); thus, this variable has been included as covariate in the analyses. No other pre-existing differences between groups emerged.
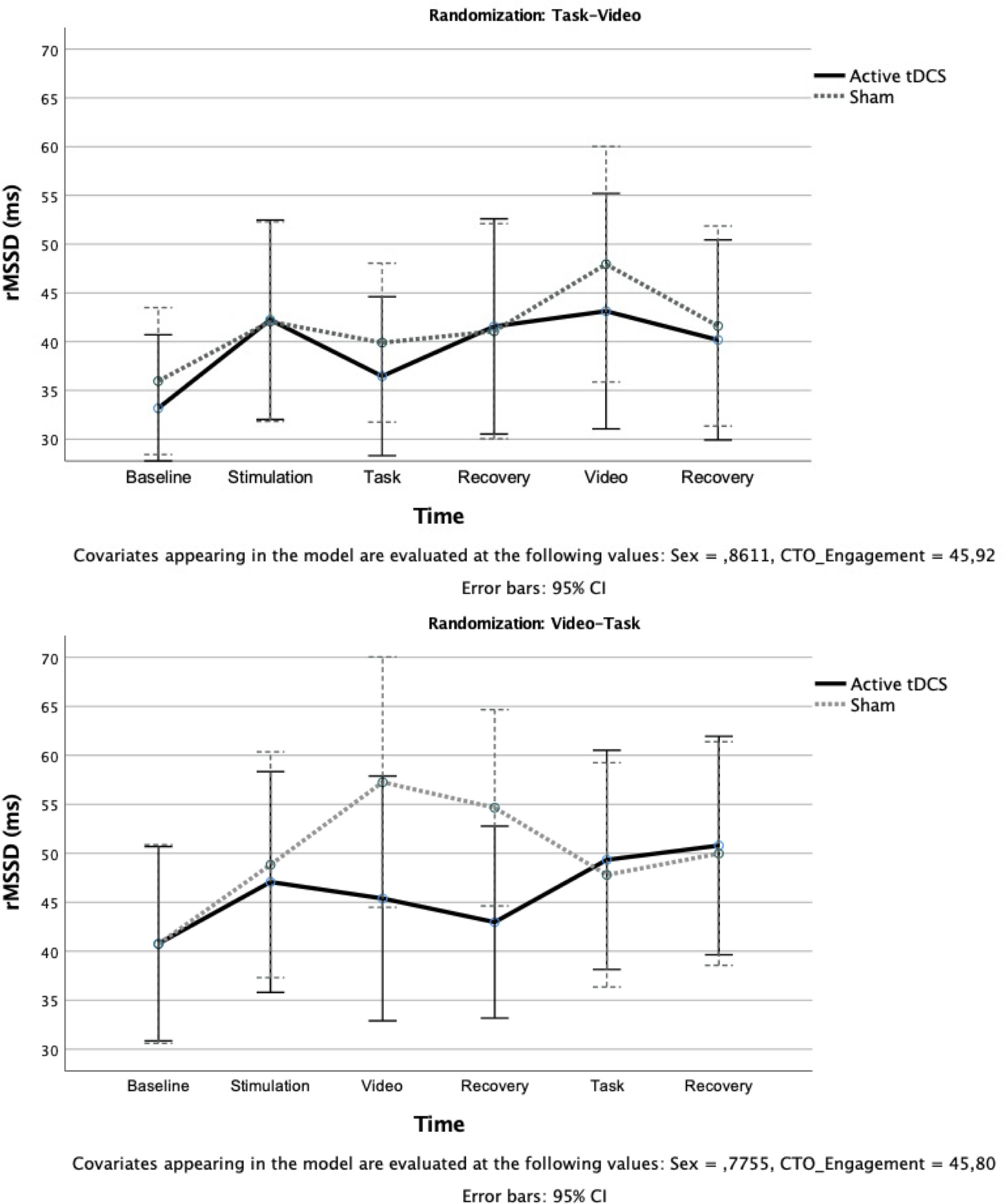
The model having rMSSD as outcome yielded significant Stimulation Type X Time, F[4316] = 5.02; p = .03; ηp2 = 0.06 and Stimulation Type X Time X Randomization, F[4316] = 3.24; p = .045; ηp2 = 0.04 interactions. In the task-video randomization, pairwise comparisons revealed increased rMSSD during stimulation, task, video, and recovery phases compared to baseline (ps < .02) and during video compared to all the other phases (ps < .01) except for stimulation (Fig. 2, upper panel). In the video-task randomization, rMSSD increased during stimulation in both groups (ps < .04), followed by a further increase during the video in the sham group (ps = .03), and a decrease in the active group (p = .045); then, the opposite pattern emerged, with a significant increase from recovery from the video to task in the active group (ps = .04) and a marginally significant decrease in the sham group (ps < .056) (Fig. 2, lower panel).
 in the different phases of the protocol for the active and sham groups the active transcranial direct current stimulation (tDCS) and sham groups in the Video first (lower panel) and Redistribution Game (upper panel) first conditions.")
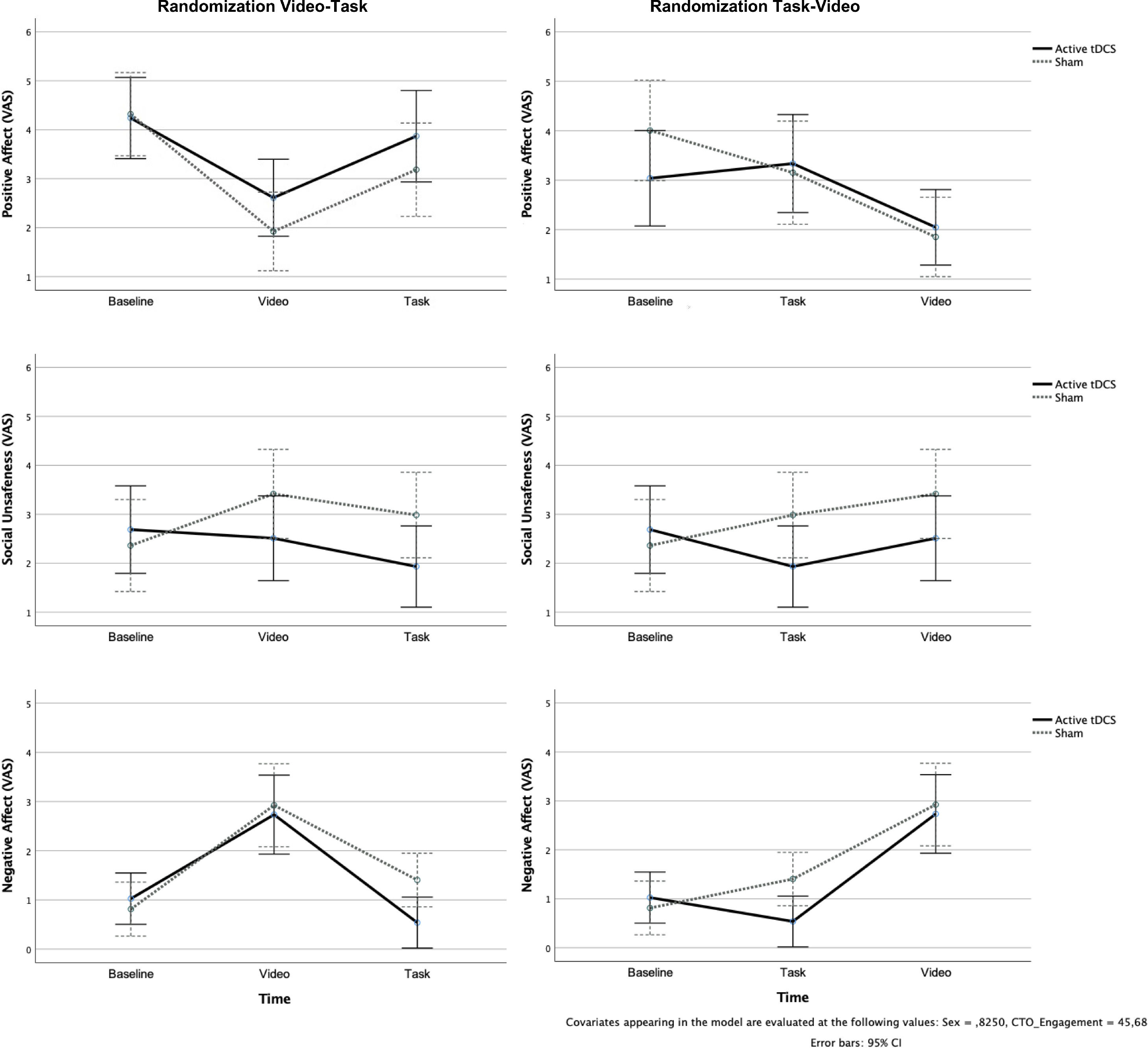
The PCA on self-reported momentary affect yielded the following three-factor model, explaining 67% of the variance and with loading values > 0.5: Positive Affect (happy, strong, content, relieved); (b) Social unsafeness (weak, self-critical, anxious); and (c) Negative Affect (sad, angry).
The GLM having Positive Affect as outcome yielded significant Time X Stimulation Type interaction, F[2170] = 4.17; p = .04; ηp2 = 0.10. Post hoc comparisons showed a stronger decrease in Positive Affect from baseline to Video in the sham compared to active conditions (p = .04) (Fig. 3, upper panel).
, Social unsafeness (middle panel), and Negative Affect (lower panel) assessed by visual-analog scale (VAS) in the different phases of the protocol for the active transcranial direct current stimulation (tDCS) and sham groups in the Video first (left column) and Redistribution Game (task) first (right column) conditions.")
Momentary Positive Affect (upper panel), Social unsafeness (middle panel), and Negative Affect (lower panel) assessed by visual-analog scale (VAS) in the different phases of the protocol for the active transcranial direct current stimulation (tDCS) and sham groups in the Video first (left column) and Redistribution Game (task) first (right column) conditions.
The GLM having Social unsafeness as outcome, yielded significant Time X Stimulation Type interaction, F[2170] = 7.37; p = .008; ηp2 = 0.080. Pairwise comparisons showed higher levels of Social unsafeness during Video and Task compared to baseline in the sham versus active conditions (ps < .003; Fig. 3 middle panel).
As to Negative Affect, significant Time X Stimulation Type interaction emerged, F[2186] = 54.91; p < .001; ηp2 = 0.371. In both stimulation conditions, Negative Affect was higher during Video compared to baseline and Task (ps < .001) and remained higher during Task compared to baseline in the sham condition only (p = .02) (Fig. 3, lower panel).
As to the Redistribution Game, only a main effect of Sex emerged F[1,83] = 7.61; p = .01; ηp2 = 0.08, with males redistributing more points compared to females (p = .03). No other significant effects emerged neither for RTs nor for number of points redistributed.
The exploratory analyses controlling for scores on the dispositional measures as covariates yielded Fear of Compassion to Self as a significant covariate in the analysis having RTs as the dependent variable (F[1,73] = 7.99; p = .006; ηp2 = 0.099), with higher scores on the FCS associated with longer RTs. No other significant main effects of or interactions with dispositional variables emerged.
DiscussionThe present study aimed to investigate whether increased cortical excitability of the right insular cortex, obtained by means of anodal tDCS over the frontotemporal lobe, would modulate engagement with suffering, indexed by HRV, and enhance the capacity to perform helpful actions.
In line with previous studies, anodal tDCS stimulation increased HRV, boosting physiological reactivity to the pain of others (i.e., reducing HRV during the video), while at the same time protecting from being subjectively overwhelmed by it (contrasting the reduction in positive affect and the increase in social unsafeness that characterized the sham group). However, in contrast to our hypothesis, the psychological and physiological changes were not mirrored by significant effects at a behavioral level, i.e., increased altruistic behavior at the redistribution game in the experimental group.
Only when the video preceded the task, HRV decreased during engagement with suffering and increased again during the task (motivation to engage in helpful actions), replicating and extending previous findings on the non-linear association between HRV and the subcomponents of compassion (Di Bello et al., 2021). Importantly, this was true for the active condition only, supporting the role of the right insula in enhancing the saliency of emotional sensory information to motivate behavior (e.g., Menon & Uddin, 2010). Interestingly, when the task preceded the video, the opposite pattern emerged: seeing the others’ suffering after having had the opportunity to behave altruistically, significantly increased HRV irrespective of the type of stimulation (active or sham). The last pattern of results also fits well with previous evidence linking high parasympathetic nervous system activation with prosocial behavior (e.g., Gilbert, 2020; Petrocchi et al., 2017) and with the predictions of the Polyvagal Theory, according to which increased vagal tone is a pre-requisite for social engagement (Porges, 2007).
At a subjective level, current results supported Gilbert's tripartite model of affect regulation (Gilbert et al., 2008), as our self-reported measures co-segregated in three clusters indicating negative affect (sad, angry), positive affect (happy, strong, content, relieved), and lack of social safeness (weak, self-critical, anxious). Anodal stimulation of the insular cortex was effective in mitigating the decrease in positive affect triggered by the video in the sham group; importantly, this was accompanied by a stronger increase in overall levels of happiness, strength, contentment, and relief in response to the Redistribution Game in the active versus control groups. Similarly, increasing cortical excitability of the insular cortex (compared to sham) decreased social unsafeness-related affect in response to both video and task and negative affect in response to the task only. These results did not fully replicate those obtained by Petrocchi et al. (2017), who found that anodal tDCS over the insular cortex increased soothing positive emotions without affecting negative affect. This inconsistency may be explained by the fact that the authors assessed subjective affectivity at rest (Petrocchi et al., 2017), while the present study examined the change of affective experience in response to conditions potentially threatening social safeness, such as a video depicting people in need and a task involving others receiving an unfair behavior.
Several limitations of the current study need to be mentioned as they may explain these unexpected findings. First, we relied on visual analog scales instead of validated questionnaires to assess changes in momentary affect. We did so to assess Gilbert's tripartite model of affect regulation and because a comparative review supported the reliability of this method to capture rapid state-dependent variations in affect (Rossi & Pourtois, 2012). Second, the current version of the Redistribution Game was based on points rather than money; this may have limited the ecological validity of the task and made it more susceptible of cultural factors. Third, the sample was predominantly composed by females and male participants appeared to behave more altruistically than females. A recent meta-analysis has shown that women are generally more generous on the Dictator Game than men, but also that this effect changes based on moderators as social distance and cultural background (Doñate-Buendía et al., 2022). In terms of cultural influences, it should be noted that the only available study testing sex differences on the Dictator Game, in Southern Italy, found similar results as the present study, thus highlighting the role of matrilineal culture in social negotiations (O'Higgins et al., 2015). We cannot exclude that the virtual version of the task employed in the current study may have made women more reluctant to give, simply due to the fact that they are generally less familiar with video-gaming compared to men (Leonhardt & Overå, 2021). Additionally, given the exploratory result on the association between fear of compassion to self and higher RTs in making altruistic choices, it is possible that personality characteristics explain current behavioral results. It is indeed well reported that fear of compassion for self correlates strongly with fear of compassion to others (e.g., Gilbert et al., 2011). In order to clarify whether the lack of effects on altruistic behavior were due to the inadequacy of the Redistribution Game to detect the desired effect, replications would be needed i) adopting more than a single task, ii) testing prosocial behavior in more than one occasion, and iii) including behaviors that are less “artificial”, such as for example the intention or attitude toward donating (Bo O'Connor et al., 2022). It is also possible that the absence of behavioral effects is due to the limited duration of the stimulation protocol (i.e., a single 15-min tDCS session). Indeed, longer stimulation protocols have been associated with enhanced behavioral effects in several cognitive tasks, as highlighted by meta-analytic findings (e.g., Dedoncker et al., 2016; Hill et al., 2016). However, stimulations of longer durations have been shown to lead to prolonged after-effects up to a limit, after which excitation may switch to inhibition (e.g., Monte-Silva et al., 2013).
The main limitation inherent to the present study is that tDCS typically delivers diffuse stimulation, which is sensitive but not specific to the targeted region. Hence, future studies combining tDCS with neuroimaging techniques are warranted to elucidate whether the observed results are specific to the insular cortex. Relatedly, to exclude that the effects were due to the location of the cathode close to the frontal pole (rather than to insular stimulation), a third cathodal condition would be desirable in future replication studies.
ConclusionsLimitations notwithstanding, current results replicate the previously reported non-linear association between compassion and parasympathetic modulation, wherein the empathic sensitivity component of the compassionate experience is characterized by reduction in HRV and the action component by increase in HRV (Di Bello et al., 2021). The present study also provides preliminarily support for the putative role of right insular cortex in “the sensitivity to suffering in self and others” phase of compassion but not in the “commitment to try to alleviate and prevent it” (Gilbert et al., 2017). Given the recognized role of the insular cortex in interoception and promising evidence that it is possible to enhance interoceptive awareness by means of non-invasive stimulation of the insular cortex (Sagliano et al., 2019), we may speculate that the empathic sensitivity phase of compassion may be enhanced by increased awareness of internal bodily states. If current findings are replicated and the above-mentioned limitations overcome, insular cortex stimulation may be combined with Compassion Focused Therapy to boost the engagement component of (self) compassion in individuals with psychopathological conditions characterized by deficits in social/emotional engagement and higher levels of self-criticism.
FundingThis research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Data availabilityThe data that support the findings of this study are available from the corresponding author, [MDB], upon reasonable request.
