

Uno de los análisis complementarios para el diagnóstico de la infertilidad masculina es el estudio de la fragmentación del ADN espermático. Una de las causas que puede explicar el daño en la molécula del ADN es el estrés oxidativo. Este viene provocado por radicales libres de oxígeno, entre los cuales se encuentra el anión superóxido (O2.−).
ObjetivoCaracterizar el exceso de estrés oxidativo en muestras recién eyaculadas y su incidencia en la fragmentación del ADN espermático.
Material y métodosSe evaluaron un total de 78 muestras de pacientes y donantes. Cada una de ellas fue clasificada en uno de los 4 niveles (EO1, EO2, EO3 o EO4) en función del nivel de estrés oxidativo hallado en el test del estrés oxidativo NBT-t (detección del anión superóxido). De cada una de las muestras se calculó la concentración de espermatozoides y el índice de fragmentación (IF) analizado a t0, t2, t6 y t24h de incubación a 37°C utilizando la técnica sperm chromatin dispersion test (SCD test).
ResultadosLos resultados obtenidos muestran que no existen diferencias significativas en el IF basal (t0h) en función del nivel de estrés oxidativo. Al estudiar la dinámica de fragmentación se observa un aumento del IF a lo largo del tiempo (siendo estable en las 2 primeras horas). El comportamiento del IF es diferente según el grado de estrés oxidativo: a medida que aumenta el estrés, la fragmentación pierde antes la estabilidad. A partir de un determinado momento, la velocidad se iguala para todos los grupos.
DiscusiónNo hemos encontrado una correlación significativa entre el nivel de estrés oxidativo y el IF basal. En lo que se refiere a la dinámica de fragmentación, las muestras tienen un comportamiento distinto en lo referente a su variación del IF en función del nivel de estrés oxidativo.
ConclusionesLos resultados insisten en la importancia de valorar la fragmentación del ADN espermático desde un punto de vista dinámico y no tan solo en la comparación estática del IF basal. Las muestras con un elevado estrés oxidativo presentan un rápido incremento de su fragmentación en las primeras horas tras el eyaculado frente a la estabilidad del resto de muestras.
One of the additional tests for the diagnosis of male infertility is the study of sperm DNA fragmentation. One reason that may explain the damage of the DNA molecule is oxidative stress. This is caused by free oxygen radicals, among which is the superoxide anion (O2.−).
ObjectiveTo measure the oxidative stress excess in semen samples and the impact on sperm DNA fragmentation.
Material and methodsA total of 78 samples from patients and donors were evaluated. Each one was classified into one of four levels (EO1, EO2, EO3 or EO4) according to the level of oxidative stress found using the NBT-test (detection of superoxide anion). Sperm concentration and DNA fragmentation index (at t0, t2, t6, and t24h during incubation at 37°C) were calculated for each sample using the sperm chromatin dispersion test (SCD).
ResultsThere were no significant differences between results of the four groups in the baseline DNA fragmentation index (t0h). When the dynamic process was studied, an increase in the fragmentation index could be observed (the first two hours remained stable). This increase varied depending on the levels of oxidative stress. The DNA fragmentation index (DFI) remained stable for less time as the levels of oxidative stress increased. There is a particular time where the velocity becomes the same in all the groups.
DiscussionNo significant correlation was found between the levels of oxidative stress and the baseline DFI. However, the behaviour of the fragmentation dynamics of the samples varied depending on the levels of oxidative stress.
ConclusionsThese results enhance the importance of understanding DNA fragmentation from a dynamic point of view rather than a static process of the baseline DFI. Samples with higher levels of oxidative stress showed a rapid increase in their DFI during the first hours after ejaculation compared with the stability shown in the rest of the samples.
Según la Organización Mundial de la Salud (OMS) una pareja infértil es aquella que después de tener relaciones sexuales frecuentes durante 12 meses, sin ninguna protección, no ha logrado un embarazo. La infertilidad afecta aproximadamente a un 15% de las parejas que tratan de concebir, y se cree que el factor masculino contribuye en aproximadamente la mitad de estos casos (Oehninger, 2001).
Para comprobar si el factor masculino influye en la fertilidad, se realiza el análisis de algunas características de muestras seminales del paciente. El análisis básico del semen consiste en medir el volumen seminal, la concentración de espermatozoides, movilidad y morfología (WHO, 2010). Cada parámetro tiene unos límites inferiores de referencia determinados por la OMS. Sin embargo, en un 15% de los casos, los parámetros medidos son superiores a dichos límites inferiores de referencia, es decir, presentan un seminograma normal pero el individuo tiene problemas de fertilidad (Guzick et al., 1998). En estos casos se debe analizar la muestra en mayor profundidad. Uno de los estudios complementarios es el de la fragmentación del ADN espermático. El término fragmentación de ADN hace referencia al ADN espermático desnaturalizado o dañado que no puede ser reparado. La integridad del ADN es importante para que tenga lugar un desarrollo normal del embrión (Practice Committee, 2015).
En la actualidad se han desarrollado muchos test que detectan de forma directa o indirecta la proporción de espermatozoides con ADN fragmentado. A pesar de su aparente funcionalidad, no se recomienda realizar este test como una prueba de rutina (Practice Committee, 2013), y varios estudios ponen en duda su utilidad clínica. El resultado de varios metaanálisis pone de manifiesto que la principal indicación del test de fragmentación del ADN espermático es en parejas que sufren aborto recurrente (Robinson et al., 2012), no obstante, otros trabajos ponen en duda esta utilidad (Coughlan et al., 2015).
El test de fragmentación del ADN espermático y su utilidad clínica son por tanto un tema controvertido, sin embargo son necesarios trabajos que investiguen la fisiopatología de la fragmentación del ADN espermático en busca de dicha utilidad clínica y su aplicación en el diagnóstico de la infertilidad masculina (Barratt et al., 2010).
Una de las posibles causas de la fragmentación del ADN espermático es el estrés oxidativo. Hay evidencias que sugieren que los radicales libres de oxígeno (reactive oxygen species [ROS]) contribuyen en el daño del semen en un 30-80% de los casos de infertilidad asociada al varón (Agarwal et al., 2006). Los ROS son productos que se obtienen durante el metabolismo celular normal. La mayor parte de la energía del cuerpo se obtiene en reacciones enzimáticas controladas de reducción del oxígeno con el hidrógeno en el proceso de fosforilación oxidativa que tiene lugar en la mitocondria durante el metabolismo oxidativo. Los radicales libres se forman durante esa reducción enzimática del oxígeno (Valko et al., 2007). Un radical libre se define como una molécula de oxígeno que contiene uno o más electrones desapareados. Los radicales libres participan en reacciones químicas en las que ceden su electrón desapareado oxidando lípidos de membrana, aminoácidos de proteínas e hidratos de carbono como los del ADN (Ochsendorf, 1999). Son capaces de causar infertilidad principalmente por 2 mecanismos: en el primero, los ROS peroxidan los ácidos grasos insaturados dañando las membranas de los espermatozoides, lo que reduce su movilidad y su habilidad para fusionarse con el oocito (De Lamirande et al., 1998). En el segundo mecanismo, los ROS dañan directamente al ADN del espermatozoide. En general, el ADN está tan compactado por las protaminas que está protegido del ataque de los radicales libres, sin embargo, los hombres infértiles suelen presentar una protaminación deficiente, lo que hace que su ADN espermático sea más vulnerable (Oliva, 2006).
En el semen hay principalmente 2 fuentes de producción de radicales libres: leucocitos y espermatozoides. La mayor parte de las muestras de semen contienen leucocitos, siendo los neutrófilos el tipo de leucocito predominante (Aitken et al., 1995). El mecanismo principal de los neutrófilos para destruir patógenos es producir ROS, lo que explica su capacidad para provocar estrés oxidativo. En el caso de los espermatozoides, la capacidad de producir ROS está inversamente correlacionada con su estado de maduración. Los espermatozoides inmaduros teratozoospérmicos tienen un exceso de citoplasma en la pieza media. Este residuo es rico en la enzima glucosa-6-fosfato deshidrogenasa, que entre otras cosas controla la producción intracelular de la nicotinamida-adenina-dinucleótido-fosfato (NADPH). La NADPH es una fuente de generación de ROS vía NADPH oxidasa, que está localizada en la membrana de los espermatozoides (Said et al., 2005).
El estrés oxidativo tiene lugar cuando la producción de ROS sobrepasa la capacidad antioxidante de los mecanismos de defensa del organismo, lo que lleva al daño celular. Los antioxidantes del plasma seminal ayudan a minimizar el estrés oxidativo de los espermatozoides eyaculados, pero no pueden actuar a nivel de la rete-testis y epidídimo (Tremellen, 2008).
Dado que el estrés oxidativo parece tener un efecto negativo directo sobre diferentes estructuras del espermatozoide y sobre el ADN, el objetivo del presente estudio es caracterizar el exceso de estrés oxidativo relacionado con la presencia de un exceso de ion superóxido (SOx) en muestras de semen recién eyaculadas, y su incidencia en la fragmentación del ADN espermático que presenta el paciente en el momento de la eyaculación y tras la incubación in vitro durante un periodo de 24h.
Material y métodosLa determinación de los niveles de estrés oxidativo y del daño en el ADN del espermatozoide se realizó en 78 muestras obtenidas en 3 clínicas diferentes: 25 muestras son de pacientes o donantes de la clínica Tambre (Madrid), 36 de la clínica Gutenberg (Málaga) y 17 de la Universidad Autónoma de Barcelona. Al tratarse de un estudio prospectivo no se establecieron criterios de selección sobre el tipo de paciente.
El procesamiento de las muestras según el centro fue el siguiente: las muestras de la clínica Tambre fueron enviadas en fresco a la Universidad Autónoma de Madrid (UAM) para realizar la dinámica (incubación de las muestras) y el estudio de la fragmentación, junto con una alícuota congelada en t0 y el resultado del estrés oxidativo (realizado en el propio centro). En el caso de la clínica Gutenberg, el estrés oxidativo y la dinámica de fragmentación (incubación y congelación en los distintos tiempos) se llevaron a cabo en el mismo centro, y las alícuotas congeladas se enviaron a la UAM para analizar la fragmentación de ADN. Las muestras de la Universidad Autónoma de Barcelona se analizaron por completo en su laboratorio.
Análisis del estrés oxidativoLos niveles de ion SOx se estudiaron utilizando el test OxiSperm (Halotech DNA, Madrid, España). La base de este test es la reacción que genera la molécula nitro blue tetrazolium (NBT) con el anión superóxido para formar diformazán, un cristal azul insoluble en agua. La cantidad de diformazán presente en el medio se relaciona con un desequilibrio de SOx en el medio, probablemente no contrarrestado por la presencia de SOD (Esfandiari et al., 2003).
Las muestras se procesaron de acuerdo con el protocolo especificado que básicamente consiste en licuar el gel reactivo que contiene el NBT y mezclarlo con una cantidad variable de microlitros de la muestra seminal, de acuerdo con la concentración espermática de la muestra. La mezcla de gel reactivo y espermatozoides con plasma seminal se incuba a 37°C durante 45min en una baño termostatizado y se evalúa el nivel de color obtenido. En función de la intensidad de la reacción, y por tanto del color, se asigna de forma visual un valor del 1 al 4 (EO1, EO2, EO3 o EO4) teniendo en cuenta la menor o mayor intensidad de color percibida.
Análisis de la fragmentación de ADNPara evaluar el daño del ADN espermático se utilizó la técnica sperm chromatin dispersion test (SCD test) en su versión comercial HaloSperm (Halotech SL, Madrid, España). Básicamente, el método consiste en someter una muestra de semen a un tratamiento ácido inicial que desnaturaliza el ADN y posteriormente a una desproteinización controlada que retira la mayor parte de las proteínas nucleares. En los espermatozoides sin una fragmentación masiva del ADN se producen nucleoides con grandes halos formados por bucles de ADN emergiendo de un core central, mientras que los nucleoides de espermatozoides con ADN fragmentado no muestran halo de dispersión o este es mínimo (Fernández et al., 2003). Dichos halos se valoran utilizando microscopia de campo claro o de fluorescencia. En función del tamaño y del aspecto del halo, cada espermatozoide se clasifica en una de las 3 categorías: halo grande/halo mediano, halo pequeño/sin halo (fragmentado) o degradado (espermatozoide altamente fragmentado).
En nuestro caso, las muestras de semen se diluyeron en PBS hasta alcanzar una concentración de 5-10 millones de espermatozoides por mililitro. En muestras poco concentradas no es necesario diluir el semen. Una vez procesadas las muestras de acuerdo con las recomendaciones expresadas en el protocolo y tras la deshidratación final de la muestra, se deja secar el portaobjetos a temperatura ambiente. Una vez seca, la muestra procesada se puede guardar a temperatura ambiente y en oscuridad hasta su tinción. Las muestras se tiñeron con SYBR Green (Molecular Probes, Inc. Willow Creek Road, Eugene) a una concentración final 10× en agua destilada. Para su utilización final, el SYBR Green 10× se diluye en proporción 1:1 con el Vectashield.
Dinámica de fragmentaciónPara el estudio de la dinámica de fragmentación, las distintas muestras seminales se incubaron en una estufa a 37°C. Durante el proceso de incubación, se procedió a un remuestreo seriado donde se obtuvieron muestras a t0 (índice de fragmentación [IF] basal), transcurridas 2h (t2), 6h (t6) y 24h (t24). Cada una de las alícuotas se procesaron mediante la técnica SCD test anteriormente detallada.
Análisis estadísticosPara analizar de un modo dinámico la fragmentación del ADN se empleó la t de Student en una muestra para valorar el incremento temporal de la fragmentación entre 2 ítems temporales y la regresión lineal simple para el conjunto del periodo analizado.
La comparación de la media de velocidad de fragmentación entre 2 grupos se realizó mediante la t de Student para muestras independientes.
ResultadosDesde un punto de vista estrictamente estático, no encontramos diferencias en el IF inmediatamente tras el eyaculado en función del nivel de estrés oxidativo (EO) de la muestra, siendo la media del IF en nuestra muestra de 21,97% (EO1: 21,50%, EO2: 23,26%, EO3: 22,19% y EO4: 19,82% ANOVA p=0,844).
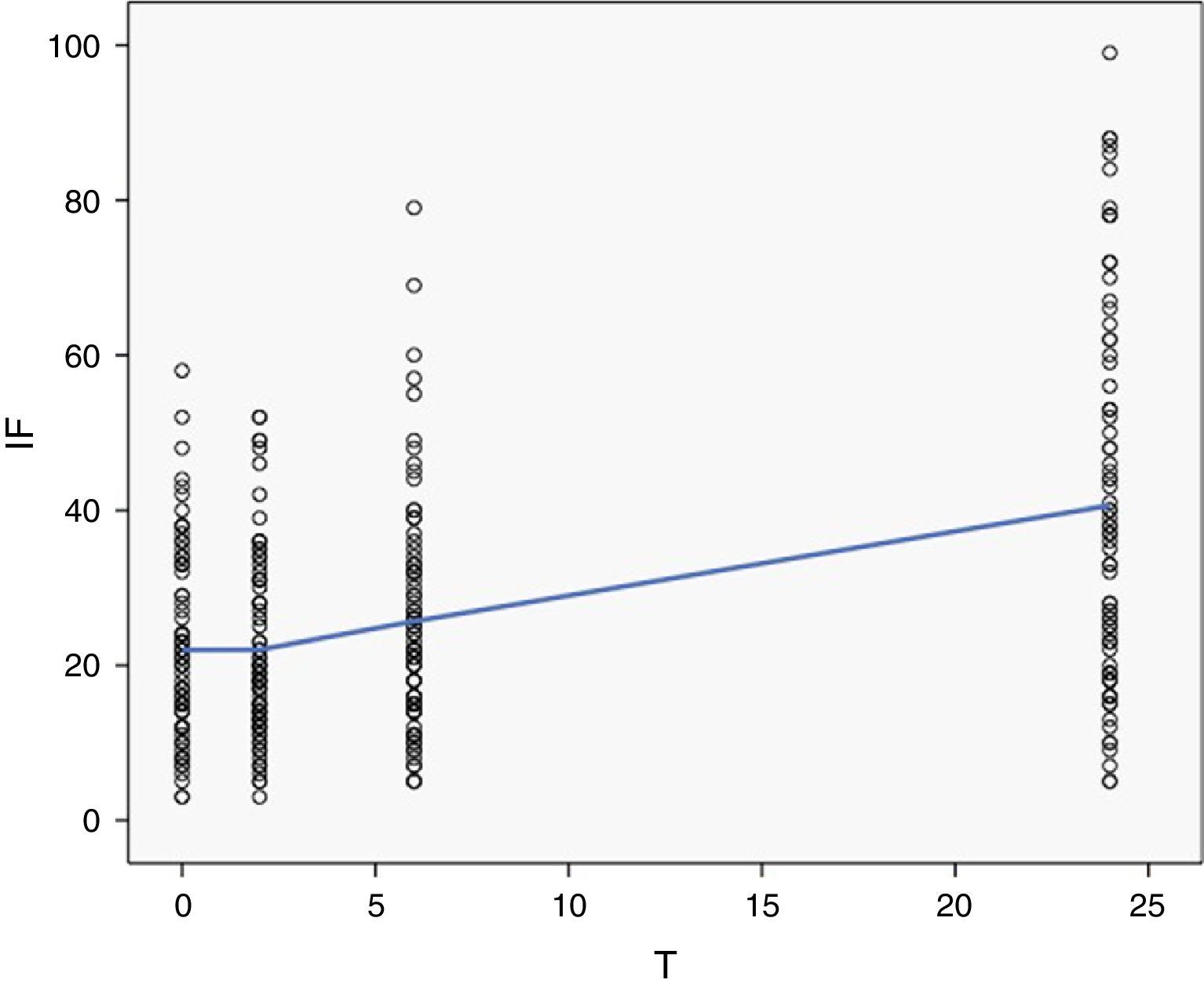
Para analizar de manera dinámica la fragmentación del ADN espermático, lo primero que hacemos es un modelo de regresión lineal univariante que nos confirma el hecho de que la evolución en el tiempo del IF se ajusta a un modelo lineal (p<0,001) con un IF basal de 21,098% y un incremento de 0,807% por cada hora transcurrida desde el eyaculado.
Analizado por tramos, podemos ver como en las primeras 2h tras el eyaculado no se produce un incremento del IF, encontrando una velocidad de aumento de fragmentación (vF) no significativamente diferente de 0 (t Student para una muestra vF: –0,038/h, p=0,945).
Sí que confirmamos ese incremento progresivo de la fragmentación a partir de la segunda hora tras el eyaculado, con una vF de 0,914/h entre las 2 y las 6h (p=0,003) y de 0,834/h a partir de la sexta hora (p<0,001). Un comportamiento que no es incompatible con el modelo lineal, dado que incluso una comparación de medias de las velocidades entre estos 3 tramos mediante una t de Student para muestras pareadas no encuentra diferencias significativas entre ellos (p=0,125 entre el primer y el segundo tramo y p=0,801 entre el segundo y el tercero) (fig. 1).

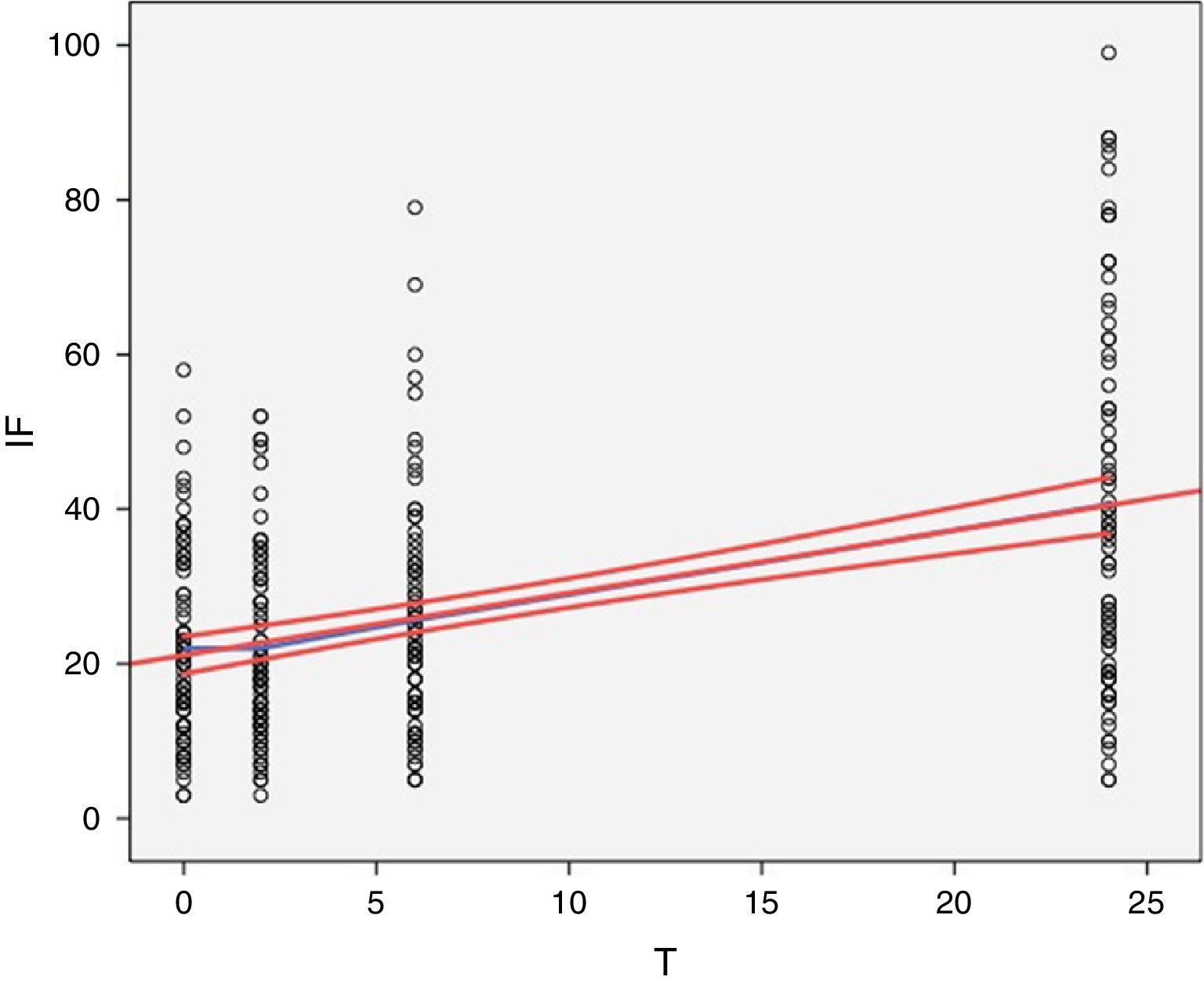
La perfecta superposición de ambos modelos se aprecia perfectamente en la representación gráfica que se muestra en la figura 2.

Superposición del modelo empírico de la figura 1 y de un modelo de regresión lineal simple del total de la muestra, representando la media de la pendiente y los valores del IC al 95% de la media.
El uso de la t de Student para una muestra es una buena herramienta para comprobar la existencia de un incremento significativo de la fragmentación con el tiempo en los distintos subgrupos. Con esta estrategia observamos un comportamiento claramente diferente según el grado de estrés oxidativo de la muestra:
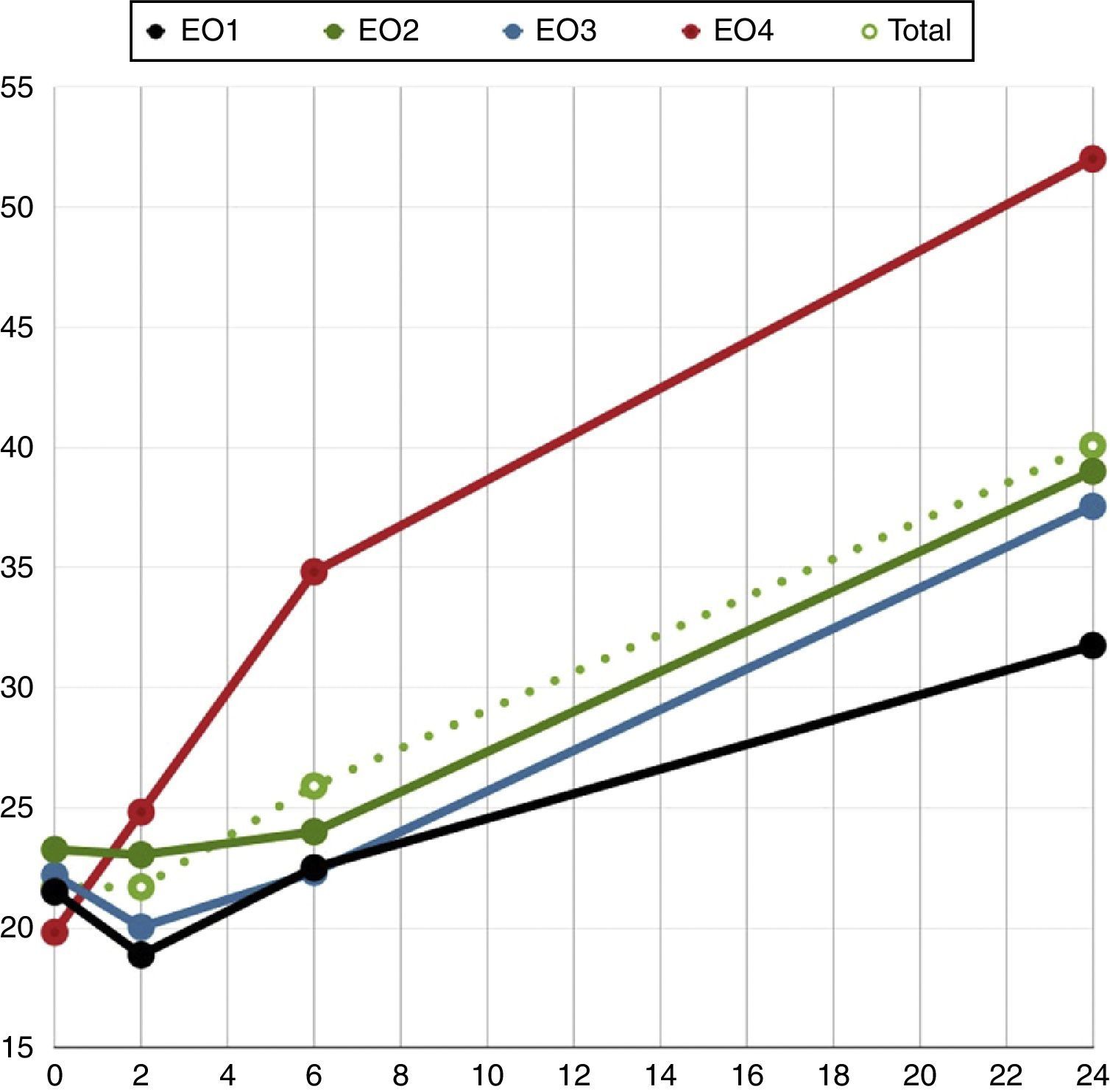
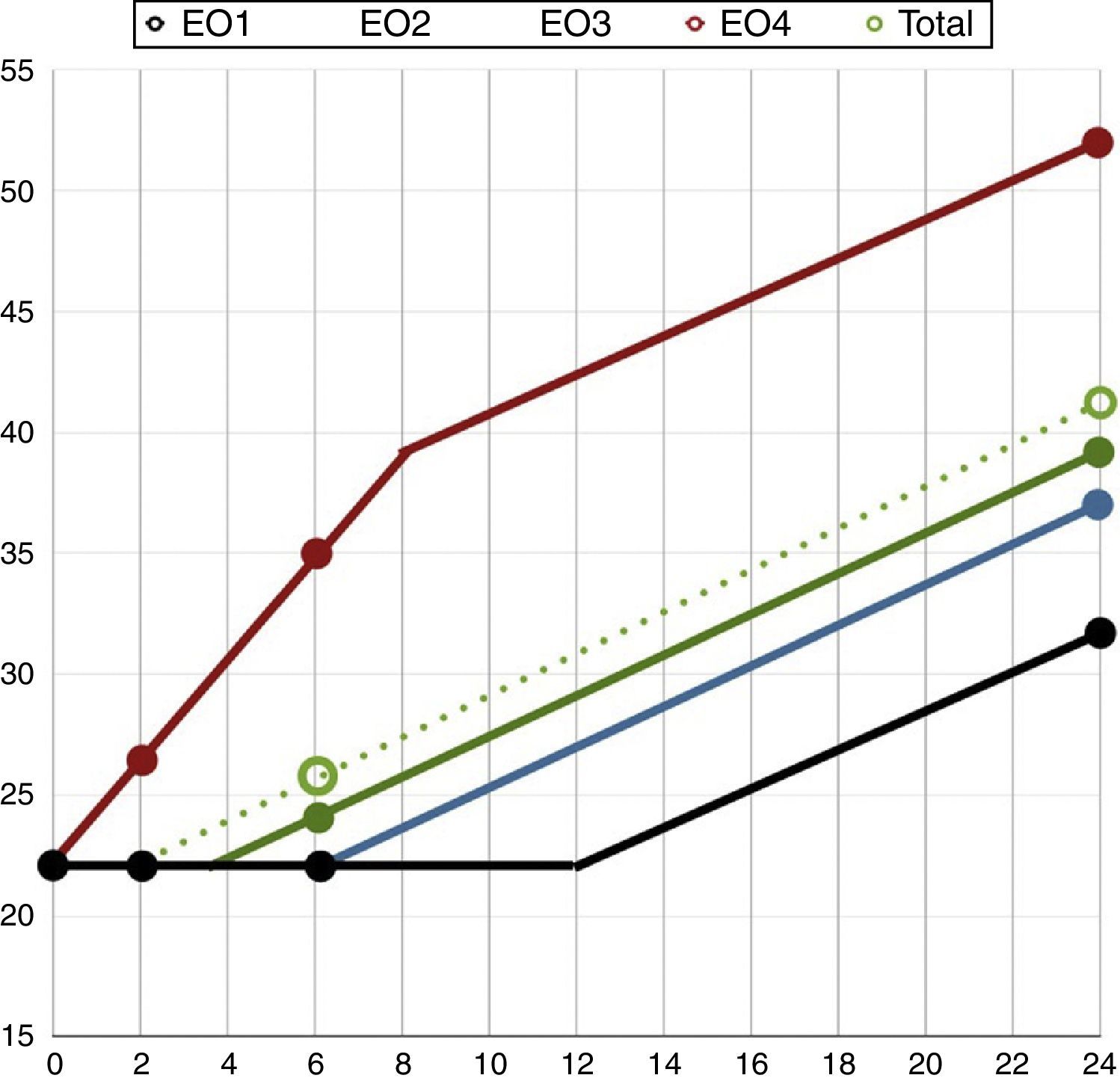
Así en la muestra con nivel EO1 encontramos que la variación temporal de la fragmentación alcanza niveles levemente superiores a la significación estadística solo a partir de la sexta hora tras el eyaculado (1.er tramo vF:–1,321/h, p=0,493; 2.o tramo vF: 0,907/h, p=0,516; 3.er tramo vF: 2,536/h, p=0,039).
En el grupo con nivel EO2 la vF es mayor, pero tampoco se alcanza antes de las 6h tras el eyaculado (1.er tramo vF: –0,157/h, p=0,833; 2.o tramo vF: 0,232, p=0,660; 3.er tramo vF: 0,837/h, p<0,001).
En el grupo con nivel EO3 el ascenso significativo del IF aparece ya a partir de la segunda hora (1.er tramo vF:–1,146/h, p=0,340; 2.o tramo vF: 0,578/h, p=0,029; 3.er tramo vF: 0,853/h, p<0,001).
Y en el grupo con nivel EO4 el incremento de la fragmentación con una vF significativamente superior a cero se produce ya desde el momento del eyaculado y en los 3 tramos de análisis (1.er tramo vF: 2,450/h, p=0,013; 2° tramo vF: 2,515/h, p=0,014; 3.er tramo vF: 0,950/h, p=0,004).
En las figuras 3 y 4 observamos el comportamiento empírico y el correspondiente a los modelos matemáticos descritos que explican las diferencias en el comportamiento de las distintas muestras con distinto nivel de estrés oxidativo.


Con los datos anteriores podemos comprender como una comparación de medias de velocidades mediante una t de Student de muestras independientes nos ofrece diferencias significativas entre la vF de la muestra con un alto estrés oxidativo (EO4) desde el mismo momento del eyaculado frente a la estabilidad de las muestras con menor estrés oxidativo en las primeras horas (2,45/h vs. –0,73/h en el 1.er tramo, p=0,016 y 2,52/h vs. 0,47/h en el 2.o tramo, p=0,044). Igualmente, a partir de determinado momento esa velocidad se iguala para todos los grupos, no siendo mayor en el grupo con mayor estrés oxidativo que en el resto (0,95/h vs. 0,80/h, p=0,0588).
DiscusiónLa velocidad de fragmentación del ADN espermático difiere entre especies y entre individuos de la misma especie (Gosálvez et al., 2009; López-Fernández et al., 2010), sin embargo, las causas de estas variaciones no se han estudiado sistemáticamente. Este estudio plantea uno de los primeros análisis que se han realizado para ver la posible correlación entre las variaciones de esa velocidad y la presencia de estrés oxidativo.
El test de fragmentación del ADN espermático nos da información del estado del material genético del espermatozoide. A la hora de interpretar esta información, podemos entender la fragmentación como un proceso estático si nos centramos en el IF basal (tras la eyaculación), o como un proceso dinámico si analizamos la evolución de la fragmentación a lo largo del tiempo. En este estudio no hemos observado que haya diferencias en el IF basal en función del nivel de estrés oxidativo, sin embargo, analizando los datos de la fragmentación a lo largo de las 24h posteyaculación, sí hemos encontrado distintos comportamientos en las muestras según el nivel de estrés oxidativo.
El IF aumenta a lo largo del tiempo independientemente del nivel de estrés oxidativo. Si fraccionamos las 24h en distintos tramos, vemos que durante las 2 primeras horas tras el eyaculado no se produce un incremento del IF, sin embargo, sí encontramos un aumento significativo en la velocidad de fragmentación a partir de esa segunda hora.
En este estudio hemos tratado de ver si el estrés oxidativo ejercido por el ion SOx tenía alguna correlación con el IF del ADN espermático y si influía en su dinámica. Tal y como hemos explicado, el IF es estable en las primeras horas. Según el nivel de estrés oxidativo las muestras pierden dicha estabilidad en un momento diferente, variando a velocidades de fragmentación también diferentes. Las muestras con un nivel de estrés oxidativo bajo (EO1) tienen índices de fragmentación estables a lo largo de las primeras horas (hasta la sexta), mientras que las muestras con niveles de estrés oxidativo superiores (EO4) aumentan la velocidad de fragmentación desde que son eyaculadas. Al cabo de un tiempo determinado, todas las muestras alcanzan una velocidad constante y similar de aumento de la fragmentación.
En la actualidad se debate la idea de que los niveles de ROS tengan significación a nivel clínico. Algunos autores han visto que esta información es de poca ayuda para dar un diagnóstico tanto en reproducción in vivo como in vitro (Tomlinson et al., 1993), mientras que otros sí piensan que los niveles de ROS pueden tener un valor predictivo (Krausz et al., 1994). En nuestro caso hemos visto que el NBT-t nos da información del comportamiento de la muestra seminal en lo referente a la velocidad de fragmentación según el nivel de estrés oxidativo. Esta información debería considerarse en la logística diaria de manejo de las muestras de semen: en aquellas muestras donde existe un estrés oxidativo elevado se recomendaría una mayor rapidez en el manejo de esas muestras, para evitar en la medida de lo posible una sobreexposición a este daño.
Por otra parte, y como opción de trabajo futuro, hay que tener en cuenta que el exceso de estrés oxidativo no se circunscribe a los singletes de oxigeno libres que existen en el plasma seminal. Los radicales hidroxilo (OH), formados a partir de la descomposición de peroxinitrito, extremadamente reactivo y que ataca la mayoría de los componentes celulares, los hidroperóxidos orgánicos (ROOH), formados por reacciones de radicales con componentes celulares como lípidos muy representados en las membranas del espermatozoide, el ácido hipocloroso, formado a partir de H2O2 por la mieloperoxidasa, soluble en lípidos y altamente reactivo y que rápidamente oxida constituyentes proteicos, incluyendo grupos tiol, grupos amino y metioninas, y por último los peroxinitritos (OONO) (Rice-Evans y Gopinathan, 1995), pueden estar jugando papeles similares a los de los singletes de oxígeno y estar asociados a cuadros clínicos diferentes. Por lo tanto, la medida del estrés oxidativo dependiente de los singletes de oxígeno supone una parte de una realidad mucho más compleja que habrá que diseccionar con estudios similares para poder entender parte de la realidad que subyace en el complejo mundo del daño celular asociado al estrés oxidativo y a la infertilidad.
ConclusionesLos resultados insisten en la importancia de valorar la fragmentación del ADN espermático desde un punto de vista dinámico y no tan solo en la comparación estática de los valores de fragmentación inmediatamente tras el eyaculado.
Las muestras con un elevado estrés oxidativo se caracterizan por un rápido incremento de su fragmentación en las primeras horas tras el eyaculado, de en torno a un 2,5% cada hora transcurrida, frente a la estabilidad del resto de muestras.
Transcurrido un tiempo determinado todas las muestras alcanzan, independientemente de su estrés oxidativo, una vF constante y similar de en torno a un 0,9% por cada hora transcurrida.
El periodo de estabilidad de la fragmentación nuclear en las muestras sin un excesivo estrés oxidativo abarca un tiempo variable en función de dichos niveles de estrés oxidativo que, aunque nuestros datos no permiten demostrar o calcular con exactitud, apuntan a que es de unas 12h en las muestras con bajo nivel oxidativo (EO1) y de 6 y 4h en los niveles de estrés oxidativo medio (EO2 y EO3 respectivamente).
Responsabilidades éticasProtección de personas y animalesLos autores declaran que para esta investigación no se han realizado experimentos en seres humanos ni en animales.
Confidencialidad de los datosLos autores declaran que han seguido los protocolos de su centro de trabajo sobre la publicación de datos de pacientes.
Derecho a la privacidad y consentimiento informadoLos autores declaran que en este artículo no aparecen datos de pacientes.
FinanciaciónLos autores declaran no haber recibido ningún tipo de financiación económica para la realización del estudio.
AutoríaLos autores firmantes cumplen los requisitos de autoría y declaran haber colaborado en la redacción del texto y en las posibles revisiones del mismo.
Conflicto de interesesLos autores declaran que no existe ningún conflicto de intereses en la elaboración del manuscrito.
