



Sarcocystis aucheniae are apicomplexan protozoa that infect South American camelids (SACs), giving rise to macroscopic cysts similar to rice grains in skeletal muscles. Visual detection of macrocysts in slaughtered animals hampers commercialization of SAC meat, a highly relevant economic exploitation for Andean rural families. Importantly, the consumption of undercooked S. aucheniae-infested meat causes gastroenteritis. A carnivore definitive host, possibly the dog, acquires the parasite when feeding on infected SAC meat, and later eliminates infective oocysts in its feces. The parasite cycle is completed when SACs ingest contaminated water or pastures. We hypothesized that parasite DNA can be detected in SAC blood using molecular methods. In order to test this hypothesis, a seminested PCR format was specifically designed to target the hypervariable 18S rRNA gene region of S. aucheniae. PCR conditions were optimized using genomic DNA extracted from macrocyst bradyzoites. A detection limit of up to 1 parasite in 10μl of llama blood was established based on DNA samples extracted from aliquots of S. aucheniae bradyzoite-spiked non-infected llama blood. The seminested PCR allowed to detect natural infections of S. aucheniae in llama blood samples originating in the Andean flatlands of Argentina. Specific amplification of S. aucheniae DNA was corroborated by amplicon sequencing. This is the first report of S. aucheniae detection in llama blood, which provides a valuable diagnostic tool for epidemiological studies and for the evaluation of the efficacy of control measures for this parasitosis.
Sarcocystis aucheniae es un protozoo apicomplexa que infecta a camélidos sudamericanos (CS), dando lugar a la formación de quistes macroscópicos similares a granos de arroz en los músculos esqueléticos. La detección visual de macroquistes en animales faenados dificulta la comercialización de la carne de CS, una explotación de gran relevancia para la economía de las familias rurales andinas. Es importante destacar que el consumo de carne infectada con S. aucheniae no suficientemente cocida causa gastroenteritis. Un hospedador definitivo carnívoro, posiblemente el perro, adquiere el parásito cuando se alimenta de carne de CS infectada y luego elimina ooquistes infectivos en las heces. El ciclo del parásito se completa cuando un CS ingiere agua o pasturas contaminadas. Hemos hipotetizado que es posible detectar ADN del parásito en la sangre de CS usando métodos moleculares. Para poner a prueba esta hipótesis, se diseñó una PCR semianidada que utiliza como blanco una región del gen 18S ARNr específica para S. aucheniae. Se optimizaron las condiciones de la PCR usando ADN genómico extraído de bradizoítos presentes en macroquistes. Se estableció un límite de detección de un parásito en 10μl de sangre de llama, basado en muestras de ADN extraído de alícuotas de sangre de llama no infectada a las que se agregaron cantidades conocidas de bradizoítos de S. aucheniae. Más aún, la PCR semianidada permitió la detección de infecciones naturales por este parásito en muestras de sangre de llama de la Puna argentina. La amplificación específica de ADN de S. aucheniae fue corroborada por secuenciación de los productos de amplificación. Este es el primer reporte de la detección de S. aucheniae en sangre de llama. Además, este estudio contribuye una herramienta diagnóstica valiosa para estudios epidemiológicos y para la evaluación de la efectividad de medidas de control para esta parasitosis.
The domestic South American camelids (SACs), llama (Lama glama) and alpaca (Lama paco), are raised mainly in Andean regions of Argentina, Chile, Bolivia and Peru. They constitute the main type of cattle in these areas due to their adaptability to harsh environmental conditions, such as altitude, lack of water, excessive slopes, and frost. South America has a population of over 7 million llamas and alpacas5,6. They are essential to the livelihood strategy of rural families, being their meat one of the most important products resulting from their exploitation. SAC meat, while similar to that of other herbivores in protein content, has reduced cholesterol amounts compared to bovine meat14. Therefore, besides being a major source of animal protein for the Andean families, it is particularly attractive for national and international gourmet cuisine8, constituting an important source of income for small and medium producers.
In general, SAC breeding is carried out under traditional management procedures and often poor sanitary conditions. One of the constraints regarding the production and marketing of SACs meat is the frequent presence of macroscopic cysts (1–5mm long) resembling rice seeds in skeletal muscle, caused by a parasitic disease called sarcocystosis. Consumption of infected raw or undercooked meat can produce gastroenteritis with nausea, diarrhea, cramps, and chills8.
Sarcocystis spp. are protozoans that belong to the Apicomplexa phylum, and as other members of this taxon, they are obligate intracellular parasites. They display an indirect life cycle, i.e. involving two hosts. In the intermediary host, the parasite develops its asexual stage and forms cysts in the musculature, and sexual reproduction takes place10 in the definitive host - a carnivore which acquires the parasite when feeding on infected meat.
In the case of SAC sarcocystosis, an analysis of full length 18S rRNA gene sequences of parasite DNA isolated from cysts of Argentine llamas has recently shown that the parasite species that produces sarcocystosis in this region is Sarcocystis aucheniae1. Dogs are accepted as the main definitive host of SAC sarcocystosis3,9,10. However, the studies supporting this notion are based on microscopic observations, which cannot differentiate between oocysts of different Sarcocystis species12. Molecular techniques are thus required to clearly establish whether dogs act as S. aucheniae definitive host and if there are other carnivores that can also fulfill this role.
Thousands of sporocysts are excreted into the environment in the feces of the definitive host. In turn, the intermediary host acquires the infection when feeding on contaminated water and/or pastures. Sporozoites are released into the small intestine and invade the blood vessels. Merogony events followed by merozoite release into the circulation take place in the endothelia of mesenteric arterioles, lymph nodes and/or the capillaries of different organs. Finally, merozoites invade muscle cells, where they form cysts containing high numbers of infective bradyzoites4. In the case of S. aucheniae, an average cyst contains approximately 20 million bradyzoites2.
Infection of SACs with S. aucheniae is usually subclinical, although experimental inoculations have shown that the parasites can be highly pathogenic or even lethal when inoculated in high doses, because of the massive destruction of the vascular endothelium of vital organs9.
During its blood passage, the parasite could generate a humoral response, and accordingly, IgG antibodies against Sarcocystis spp. were found with high prevalence in a serological survey of llamas in Jujuy, Argentina11.
In this study we hypothesized that S. aucheniae DNA can be detected in the blood of infected llamas. To test this hypothesis we developed a S. aucheniae-specific seminested PCR, established its sensitivity and tested it in blood samples of llamas from Buenos Aires and Jujuy suspected of having low and high prevalence of S. aucheniae infections, respectively.
Materials and methodsSamples and DNA extractionMacroscopic S. aucheniae cysts (4–5mm) were obtained from the intercostal and cervical muscles of a slaughtered llama (Llama lama) from the NW province of Jujuy, Argentina. Cysts (n=6) were separated from surrounding tissues and excised with a scalpel. Their contents, consisting of a suspension of S. aucheniae bradyzoites2 were pooled in an Eppendorf tube and diluted to 150μl with phosphate buffer saline, pH 7.4 (PBS). An aliquot of 100μl was reserved for DNA extraction, while the rest was diluted 1:100 with PBS and bradyzoites were counted in a Neubauer hematocytometer. The measured density of this parasite suspension was consistent with previous reports that showed that one average sized S. aucheniae cyst (∼4mm long) contains 2.2±0.4×107 bradyzoites2. Blood samples were obtained from adult llamas of Abra Pampa, Province of Jujuy (L_1022, L_1034, L_863, L_961) and Castelar, Province of Buenos Aires (26, 34, 137), Argentina. Blood was aseptically withdrawn from the jugular vein into Vacutainer tubes containing 3.8% sodium citrate (Becton Dickinson), transferred to plastic tubes and kept frozen at −20°C until use. Blood from animal 137, which was determined to be negative for S. aucheniae infection by seminested PCR as described below, was used to determine the sensitivity of the detection technique. To this end, 950μl blood aliquots were spiked with 50μl of serial (1:10) dilutions of the bradyzoite suspension, resulting in final parasitemias of 1 to 105 parasites per ml of blood. DNA from macrocysts, field blood samples and spiked blood aliquots was extracted using the DNeasy blood and tissue kit (Qiagen), following the manufacturer's instructions. In the case of blood samples, 100μl blood were applied to the kit and in all cases, elution was effected in 100μl. The concentration of the DNA sample extracted from the macrocyst bradyzoite suspension was spectrophotometrically determined (Nanodrop ND-1000, program ND-1000 3.3) to be 583μg/μl. This sample was used to set up the seminested PCR optimal conditions. All DNA samples were kept at −20°C until further use.
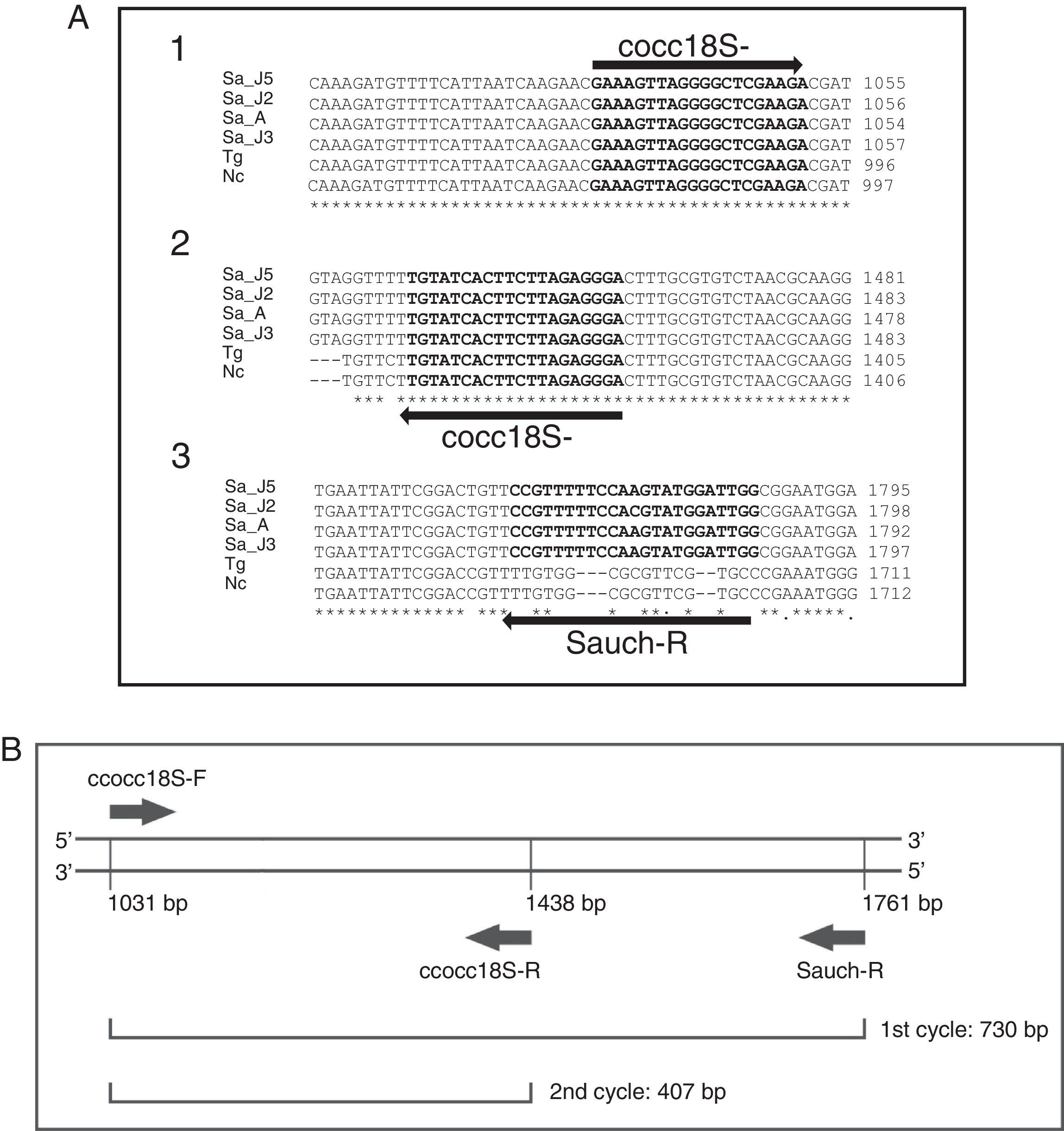
Design of primers for seminested PCRThe 18S rRNA gene hypervariable region sequences of S. aucheniae, Toxoplasma gondii and Neospora caninum (GenBank accession numbers AF017123.1, L24381.1 and L24380.1, respectively) were aligned by ClustalW (www.ebi.ac.uk/Tools/msa/clustalw2/). Two primers were designed in DNA regions common to the three coccidian protozoa (cocc18S-F: 5′-GAAAGTTAGGGGCTCGAAGA-3′ and cocc18S-R: 5′-CCCTCTAAGAAGTGATACA-3′), while primer Sauch-R (5′-CCAATCCATACTTGGAAAAACGG-3′) hybridized with a S. aucheniae-specific region (Fig. 1). The specificity of this primer was checked using Primer Blast (http://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi?LINK_LOC=BlastHome), and the lack of loop formation by all primers was verified using the Oligoanalyzer software (www.idtdna.com/analyzer/applications/oligoanalyzer/).
 ClustalW alignments of 18S rRNA gene partial sequences of S. aucheniae isolates from Argentina (Sa_J2, Sa_J3, Sa_J5) and Australia (Sa_A), and the corresponding sequences of Neospora caninum (Nc) and Toxoplasma gondii (Tg) (GenBank accession numbers: KF383266, KF383267, KF383268, AF017123, U03069 and U03070, respectively). Hybridization regions of primers cocc18S-F, cocc18S-R and Sauch-R are shown in bold. Nucleotide positions are shown to the right. Note that primer Sauch-R hybridizes with a sequence region that is not conserved in N. caninum or T. gondii. (B) General scheme of the seminested PCR.")
S. aucheniae seminested PCR design. (A) ClustalW alignments of 18S rRNA gene partial sequences of S. aucheniae isolates from Argentina (Sa_J2, Sa_J3, Sa_J5) and Australia (Sa_A), and the corresponding sequences of Neospora caninum (Nc) and Toxoplasma gondii (Tg) (GenBank accession numbers: KF383266, KF383267, KF383268, AF017123, U03069 and U03070, respectively). Hybridization regions of primers cocc18S-F, cocc18S-R and Sauch-R are shown in bold. Nucleotide positions are shown to the right. Note that primer Sauch-R hybridizes with a sequence region that is not conserved in N. caninum or T. gondii. (B) General scheme of the seminested PCR.
The amplification of a hypervariable region of the 18S rRNA gene was performed using two consecutive PCR rounds. Both reactions were carried out in a final volume of 25μl, containing 0.5μM of each primer, 0.2mM of each dNTP (Thermo Fisher Scientific), 0.003 U/μl GoTaq polymerase and its corresponding buffer (Promega, Madison, USA), and 1μl DNA from the above described samples for the first round, or from the product of the first round of amplification for the second. The primers used were: cocc18S-F and Sauch-R for the first, and cocc18S-F and cocc18S-R, for the second round, and cycling conditions were as follows: denaturing at 95°C for 10min, followed by 30 cycles of 95°C for 45s, 59°C or 55°C for 45s, for the first and second round, respectively, 72°C for 45s, and a final extension period of 5min at 72°C. Negative controls containing no template were run in parallel in both rounds to rule out amplification due to contamination. Amplification products were separated by horizontal electrophoresis (80V, 40min) in ethidium bromide-containing 0.8% agarose gels, in TAE buffer (40mM Tris-acetate/1mM EDTA, pH 8). A DNA ladder (1 Kb Plus, Life Technologies) was included. Amplification products were observed in a UV transilluminator, and images were documented using a UVItec gel documentation equipment.
SequencingPCR products obtained from field llama blood DNA samples were purified from the agarose gels using GFX PCR DNA and gel band purification kit (GE Healthcare), following the manufacturer's instructions, quantified by Nanodrop spectrophotometric measurements and sequenced using an ABI3500xl equipment (Applied Biosystems), at the Sequencing Service of the CICVyA, INTA. Sequences were cleaned using Sequence Massager (www.attotron.com/cybertory/analysis/seqMassager.htm), analyzed by BLASTn (http://blast.ncbi.nlm.nih.gov/Blast.cgi), and aligned by ClustalW with the corresponding 18S rRNA gene regions of S. aucheniae Argentine (KF383266; KF383267; and KF383268) and Australian (AF017123) isolates. Percentage identities were calculated using MATGAT1.
Results and discussionThis work focused on testing whether S. aucheniae DNA could be detected in blood of SACs. For this purpose, a seminested PCR was developed targeting the only gene sequence available so far for this parasite, the 18S rRNA gene2. The hypervariable region of this gene is polymorphic, allowing the development of species-specific detection methods2,7,13. Importantly, primers were designed to differentiate DNA of S. aucheniae from that of N. caninum and T. gondii, two coccidian parasites also infecting SACs9,11. Additionally, it was ensured that primers would hybridize with all S. aucheniae 18S rRNA sequences available in the Genbank, which correspond to an Australian and three Argentine isolates (Fig. 1A).
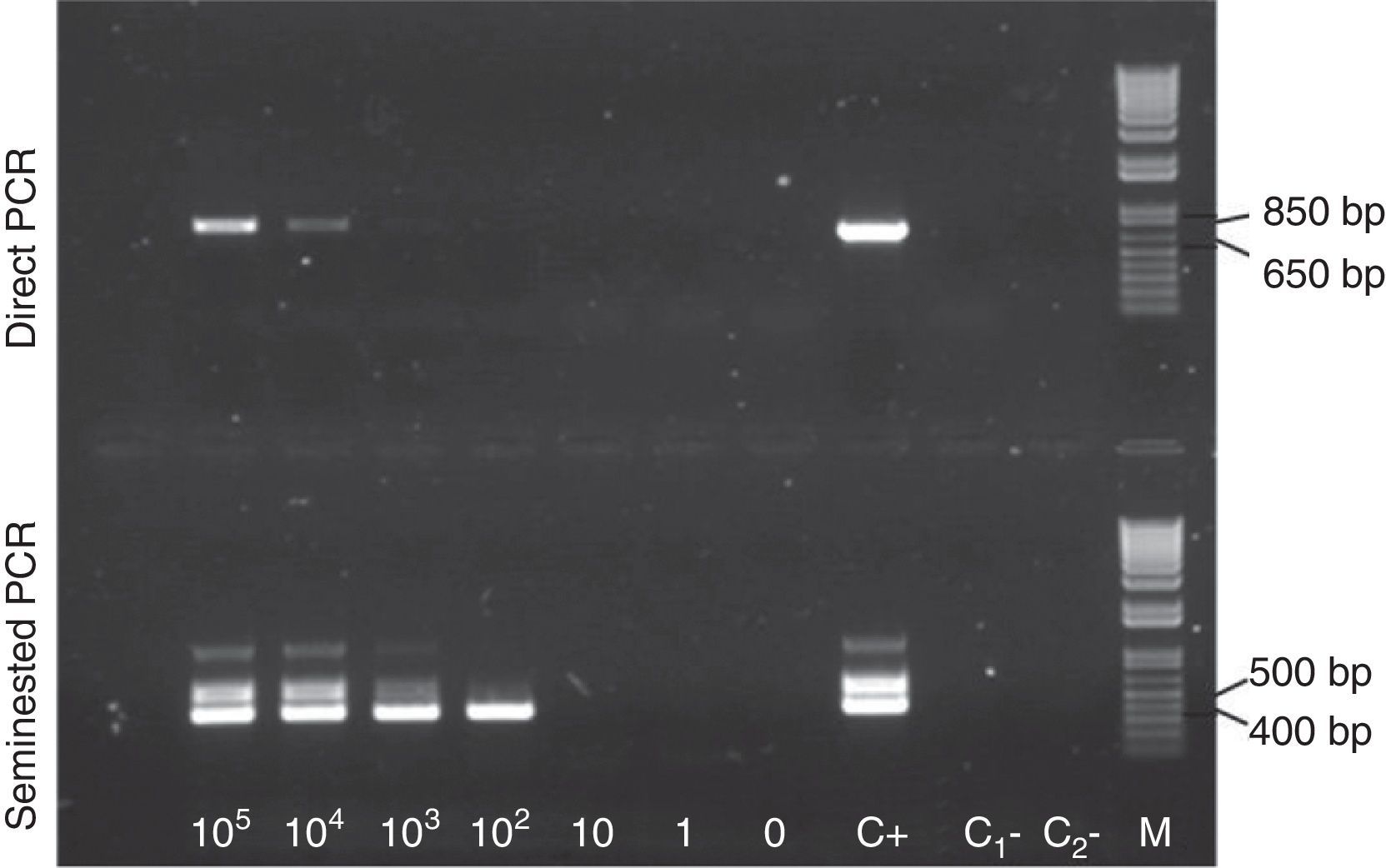
The seminested PCR consisted of a first round using cocc18S-F and Sauch-R primers, and a second round using cocc18S-F and cocc18S-R primers, with products of an expected size of 730 and 407bp, respectively (Fig. 1B). The assay was first tested using a DNA sample extracted from S. aucheniae cysts, and amplification products, analyzed by horizontal electrophoresis, were consistent with the expected sizes. Occasionally, two bands of a larger size were also observed among the amplification products of the second round (Fig. 2).
 and second (seminested PCR) amplification rounds were observed under UV light. C+: positive control where DNA extracted from a S. aucheniae macrocyst was used as template. C1− and C2−: negative controls of the first and second rounds, respectively. M: 1kb Plus DNA marker. A ∼400bp band can be observed in the second round of amplification until a parasitemia of 102 bradyzoites/ml blood.")
Sensitivity of S. aucheniae seminested PCR. Aliquots of blood from a S. aucheniae-negative llama were spiked with serial dilutions of a bradyzoite suspension, resulting in final parasitemias of 0 to 105 bradyzoites/ml. DNA was separately extracted from each aliquot and amplified by seminested PCR. The figure shows an ethidium bromide-stained agarose gel where the products of the first (direct PCR) and second (seminested PCR) amplification rounds were observed under UV light. C+: positive control where DNA extracted from a S. aucheniae macrocyst was used as template. C1− and C2−: negative controls of the first and second rounds, respectively. M: 1kb Plus DNA marker. A ∼400bp band can be observed in the second round of amplification until a parasitemia of 102 bradyzoites/ml blood.
To evaluate the parasitemia detection limit of the method, a bradyzoite suspension was obtained from S. aucheniae cysts and its cell density was microscopically evaluated. Known amounts of parasites were added to aliquots of a llama blood sample (animal No. 137) that had been determined to be negative for S. aucheniae DNA, using the seminested PCR described in this work. DNA was extracted from each sample and used as template in seminested PCR determinations. As shown in Figure 2, clear detection in the first PCR round could be achieved in samples containing down to 104 parasites per ml blood, while the second PCR round significantly extended the detection limit to 102 parasites per ml blood.
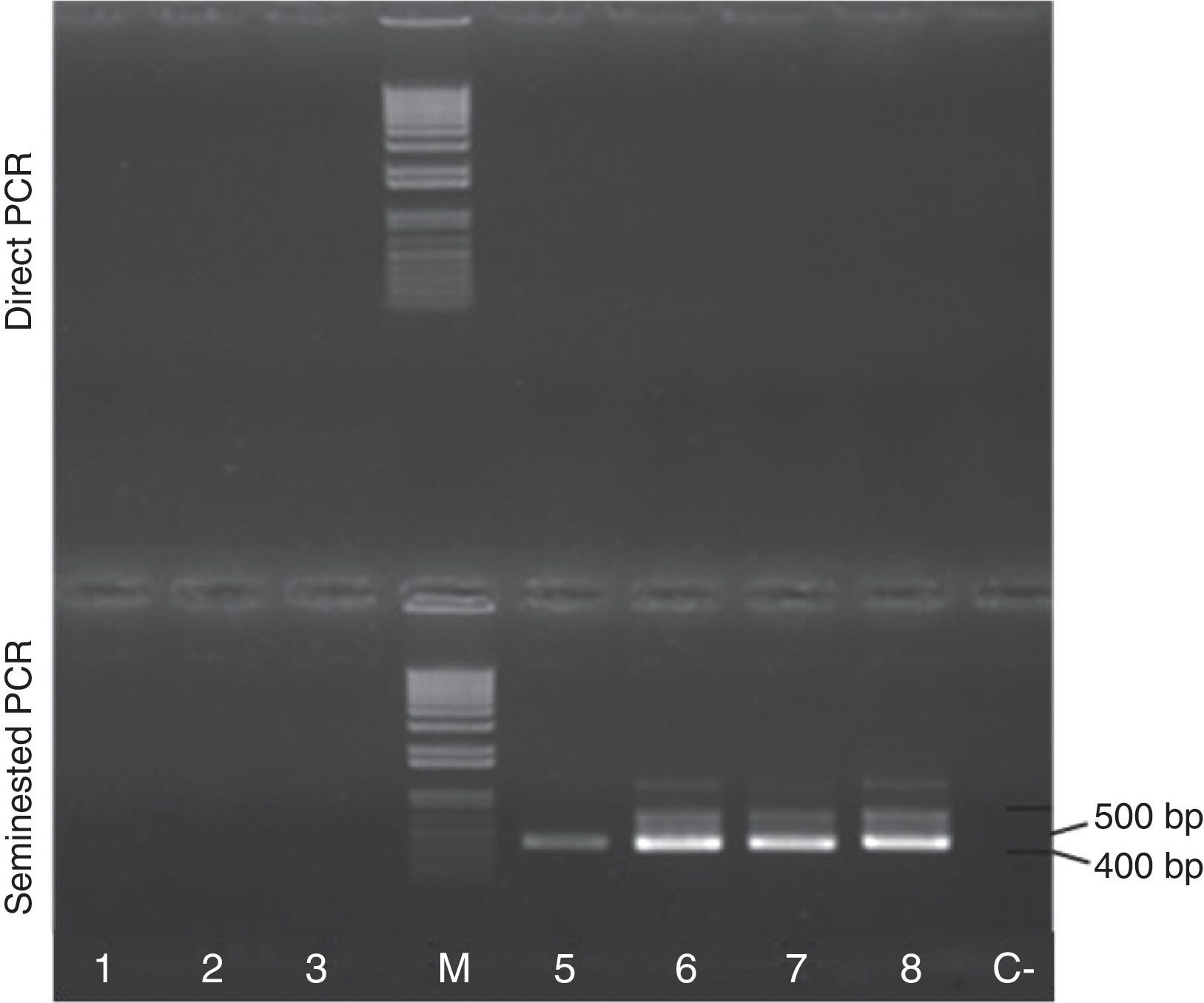
Finally, the seminested PCR was tested in DNA samples extracted from the blood of llamas raised in two regions of Argentina, Buenos Aires Pampa (n=3) and Jujuy Puna (n=4), under different management and agro-ecological conditions. While the samples from Buenos Aires corresponded to llamas raised in a pen under controlled feeding conditions, the samples of Jujuy Puna corresponded to animals allowed to graze freely. Additionally, both regions differ in humidity, temperature amplitude and altitude. Herds of llamas from Puna have been reported to present a high prevalence of anti-Sarcocystis sp. antibodies11. Noteworthy, all samples of llamas from Jujuy, and none from Buenos Aires, yielded positive amplification in the second round of the seminested PCR. The first PCR round showed no amplification in any of the samples (Fig. 3). The second round amplicons of two samples (L_1034 and L_991) were further analyzed. To this end, the three resulting bands were separately cut out of the agarose gels, purified and sequenced. In all cases, the sequences were identical between both samples, and yielded maximal BLAST hits (evalue 0) with 18S rRNA genes of S. aucheniae isolates from Argentina (J2, J3, J5) and Australia. The percent identities between the 407bp amplicons and the corresponding 18S rRNA gene fragments of S. aucheniae isolates, as calculated by MATGAT, were 99.5, 98.7, 98.4 and 97.4% for isolates J5, J2, Australia and J3, respectively.
 and Jujuy (lanes 5–8: animals No. L_1022, L_1034, L_863, L_961, respectively) was amplified by seminested PCR and analyzed by gel electrophoresis, as in Figure 2. M: 1kb Plus DNA marker; C−; negative control. Note the presence of a main ∼400bp band in the four samples from Jujuy.")
Detection of S. aucheniae DNA in the blood of llamas from Jujuy, Argentina. DNA extracted from blood aliquots of llamas from the Province of Buenos Aires (lanes 1–3: animals No. 26, 34 and 137, respectively) and Jujuy (lanes 5–8: animals No. L_1022, L_1034, L_863, L_961, respectively) was amplified by seminested PCR and analyzed by gel electrophoresis, as in Figure 2. M: 1kb Plus DNA marker; C−; negative control. Note the presence of a main ∼400bp band in the four samples from Jujuy.
Subsequent BLAST hits corresponded to 18S rRNA gene sequences from Sarcocytis species infecting other intermediary hosts, such as S. sinensis (bovine), S. capreolicanis (roe deer), S. grueneri (reindeer), S. suihominis (pig), S. hominis (bovine) and S. capracanis (goat).
Notably, a second species of Sarcocystis, S. lamacanis, has been detected in SACs and reported to be the causative agent of small cysts in the cardiac muscle12. In-depth molecular analysis of these parasites is still pending and only some fragments of the 18S rRNA gene have so far been deposited in the GenBank (accession numbers DQ100056, AY840990, EF640652, EF640651). Only a 60 nucleotide segment is shared between L_1034 and L_991 amplicons and one of these S. lamacanis 18S rRNA gene fragments with relatively low nucleotide identity (65%), thus indicating that the amplification products of the seminested PCR here described is not due to S. lamacanis infection.
Furthermore, the amplicons showed only 91.6% identity with corresponding 18S rRNA gene fragments of N. caninum (U03069) and T. gondii (TGU03070), showing that the sequence of the amplified fragment can be easily distinguished from one that would result from infection with one of these coccidian parasites.
These results confirm that the observed amplification bands correspond to S. aucheniae DNA.
In conclusion, the hypothesis that S. aucheniae infection can be detected in the blood of SACs has to be accepted. Furthermore, these results indicate that (i) the developed seminested PCR could be suitable for the diagnosis of S. aucheniae in blood samples, (ii) a second amplification step is necessary for parasite detection in field llama samples due to low parasitemia levels; (iii) S. aucheniae parasitemia levels in the tested llamas from Jujuy, Argentina, ranged from 102 to 104 parasites/ml, since detection was possible in the second PCR round, but not in the first. Additionally, the observation that primers hybridize both with S. aucheniae 18S rRNA gene sequences obtained from llama isolates from Argentina and also from an alpaca isolate from Australia (Fig. 1), suggests that the method could be useful for the diagnosis of S. aucheniae infections of both types of SAC from different geographic regions. As a drawback, the primer design employed could amplify DNA from other coccidia, such as T. gondii, N. caninum or other Sarcocystis spp. in high parasitemia scenarios. Thus, only sequencing of the amplicon undoubtedly ensures a correct diagnosis. However, high coccidian parasitemias seem to be rare among llamas. Supporting this notion, DNA samples extracted from ten S. aucheniae-positive and twenty negative llama blood samples from Argentina, in accordance with the seminested PCR described here, were amplified by direct PCR using the primers of the second round (cocc18S-F and cocc18S-R), with negative results in all cases (results not shown).
The availability of a sensitive and specific tool for S. aucheniae molecular diagnosis opens the possibility to deepen our understanding of SAC sarcocystosis, including its geographical distribution, the existence of correlations between infection rates and different parameters such as age and breeding conditions, the development of parasitemia in experimentally infected animals and the correlation between molecular detection of the parasite and the presence of antibodies against it.
In addition, a specific molecular method is of paramount importance to determine S. aucheniae definitive hosts, since the studies carried out so far in dogs are based on microscopic observations that are inadequate to differentiate S. aucheniae oocysts from those of other Sarcocystis spp.3. Once dogs are unequivocally confirmed as definitive hosts for S. aucheniae, the effectiveness of anti-parasitic treatments in the shedding of infective oocysts into the feces could also be evaluated.
Current research from our laboratory is focused on improving the molecular detection of S. aucheniae using other primer sets that avoid the need of diagnosis confirmation by sequencing, applying these methodologies to statistically valid numbers of SACs of different geographic regions and raised under different conditions, and checking whether a correlation exists between the presence of cysts in the muscles and parasite DNA in blood. If the latter is the case, a molecular tool could be applied for the diagnosis of sarcocystosis in living SACs that would allow making rational decisions about slaughtering. As previously stated, SAC meat production constitutes an important family-based agricultural exploitation in Andean regions, which has high growth potential. However, until SAC sarcocystosis is better understood and controlled, this exploitation will remain precarious and the growth of this regional economy will be postponed.
Ethical disclosuresProtection of human and animal subjectsThe authors declare that the procedures followed were in accordance with the regulations of the relevant clinical research ethics committee and with those of the Code of Ethics of the World Medical Association (Declaration of Helsinki).
Confidentiality of dataThe authors declare that no patient data appear in this article.
Right to privacy and informed consentThe authors declare that no patient data appear in this article.
Conflict of interestThe authors declare that they have no conflicts of interest.
This work was financially supported by grants from CONICET (PIP 114 20080100653), ANPCyT (PICT 2014-3747) and INTA (SALJU-1232204). The authors gratefully acknowledge Dr. Gisela Marcoppido, Instituto de Patobiología, CICVyA, for the provision of llama blood samples from INTA-Castelar.
