




It has been established that a decrease in the population of Gluconacetobacter diazotrophicus associated with sugarcane occurs after nitrogen fertilization. This fact could be due to a direct influence of NH4NO3 on bacterial cells or to changes in plant physiology after fertilizer addition, affecting bacterial establishment. In this work, we observed that survival of G. diazotrophicus was directly influenced when 44.8mM of NH4NO3 (640mgN/plant) was used for in vitro experiments. Furthermore, micropropagated sugarcane plantlets were inoculated with G. diazotrophicus and used for split root experiments, in which both ends of the system were fertilized with a basal level of NH4NO3 (0.35mM; 10mgN/plant). Twenty days post inoculation (dpi) one half of the plants were fertilized with a high dose of NH4NO3 (6.3mM; 180mgN/plant) on one end of the system. This nitrogen level was lower than that directly affecting G. diazotrophicus cells; however, it caused a decrease in the bacterial population in comparison with control plants fertilized with basal nitrogen levels. The decrease in the population of G. diazotrophicus was higher in pots fertilized with a basal nitrogen level when compared with the corresponding end supplied with high levels of NH4NO3 (100dpi; 80 days post fertilization) of the same plant system. These observations suggest that the high nitrogen level added to the plants induce systemic physiological changes that affect the establishment of G. diazotrophicus.
La población de Gluconacetobacter diazotrophicus asociada a la caña de azúcar disminuye después de la fertilización nitrogenada, lo cual podría ocurrir por la influencia directa del NH4NO3 sobre la supervivencia bacteriana, o por cambios en la fisiología de las plantas, que impiden el establecimiento bacteriano. En el presente trabajo se observó que en experimentos in vitro la supervivencia de G. diazotrophicus fue influenciada por 44,8mM de NH4NO3 (640mgN/plant). Además, G. diazotrophicus fue inoculado en plántulas micropropagadas de caña de azúcar, que fueron usadas para realizar experimentos de raíz dividida, en las que ambos extremos de los sistemas se fertilizaron con un nivel basal de NH4NO3 (0,35mM; 10mgN/planta). A los 20 días posteriores a la inoculación (dpi), la mitad de plantas fueron fertilizadas en uno de sus extremos con una dosis elevada de NH4NO3 (6,3mM; 180mg of N/plant). Este nivel fue menor al que afectó directamente a las células de G. diazotrophicus; sin embargo, provocó una disminución de la población bacteriana en comparación con plantas testigo fertilizadas con niveles basales de nitrógeno. La disminución de la población fue mayor para raíces fertilizadas con un nivel basal de nitrógeno en comparación con las raíces fertilizadas con altos niveles del mismo sistema de plantas (100dpi; 80días posfertilización). Estas observaciones indican que el alto nivel de nitrógeno añadido a las plantas inducen cambios fisiológicos sistémicos que afectan el establecimiento de G. diazotrophicus.
Gluconacetobacter diazotrophicus is a gram-negative bacterium, initially isolated as endophyte from Brazilian sugarcane plants8 and subsequently from sugarcane plants in other countries6,34. In addition, G. diazotrophicus has also been isolated from the inner tissue of diverse hosts13, such as Ipomoea batatas, Pennisetum purpureum12, Saccharococcus sachari3,31, Eleusine coracana18, pineapple37, and also from the rhizosphere of Coffea arabica17. More recently this bacterium was isolated from wetland rice27, carrot, raddish and beetroot19 and was related to a dominant phylotype detected as endophyte from needles of Pinus flexilis and Picea engelmannii using 16S rRNA pyrosequencing9.
G. diazotrophicus is a nitrogen fixing bacterium that produces phytohormones, such as indol acetic acid15,32,33 and gibberellins4. This bacterial species is able to stimulate the growth of sugarcane after its inoculation25,35,36,40. The principal mechanism for stimulating plant growth occurs through the auxinic via32,35 and depends on the sugarcane variety and the genotype of G. diazotrophicus25.
Isolation of G. diazotrophicus from sugarcane plants depends on the amount of nitrogen fertilization applied to the crops: the higher the level of nitrogen applied to the crops, the lower the probability to isolate G. diazotrophicus14,28,31. In addition, seven genotypes of G. diazotrophicus associated with sugarcane plants fertilized with low levels of nitrogen were identified in Brazilian fields and the diversity between them seemed to be affected by the high levels of nitrogen applied to sugarcane crops7 while only one genotype was detected in sugarcane plants fertilized with high levels of nitrogen in Mexican fields. Moreover, G. diazotrophicus colonization is reduced in plants fertilized with high doses of nitrogen14,22,25.
The decrease in the population of G. diazotrophicus associated with sugarcane plants could be due to pleomorphic changes that occur while culturing bacteria in the presence of high nitrogen concentrations29. Additionally, it has been proposed that the decrease in the population of G. diazotrophicus associated with sugarcane could be due to physiological changes that the plant suffers in the presence of high nitrogen fertilization7,14.
Split root experiments have been developed to evaluate the systemic effect of a specific substance on plants, when this substance is supplied only on one end of the plant, while the other end could be used as control42, but also to evaluate the systemic effect on plant pathogens due to the action of the induced systemic resistance produced by rhizobacteria1.
In this work we show a statistical analysis of the behavior of the population of G. diazotrophicus present inside the roots and in the rhizosphere using split root experiments, both at high or low nitrogen levels in the form of NH4NO3. In accordance with our results, the negative effect of nitrogen on the population of G. diazotrophicus is influenced by the plant.
Materials and methodsBacterial strains used for in vitro studies were G. diazotrophicus PAl 5T, PAl 3 and UAP 5560, each one corresponding to a different genotype. PAl 5T represents the predominant genotype isolated from different Brazilian sugarcane varieties (ET 3), PAl 3 corresponds to a rare genotype not frequently isolated (ET 5) and UAP 5560 is the predominant genotype isolated from different Mexican sugarcane varieties (ET1)7.
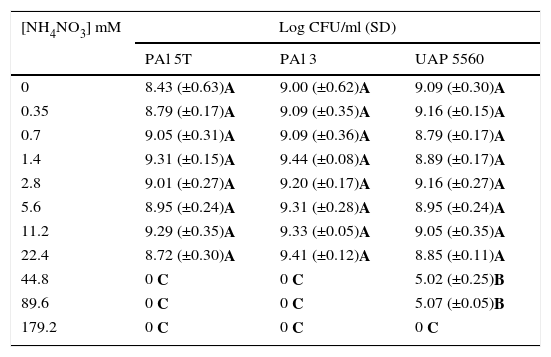
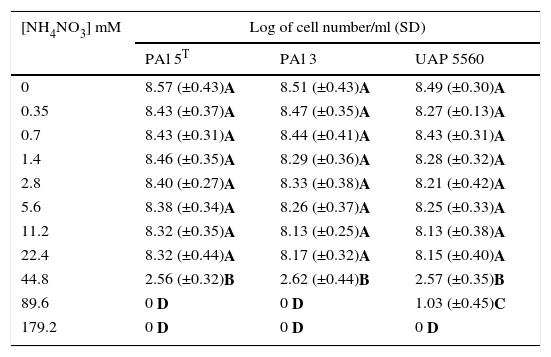
Experiment 1. In vitro assaysThe ability of G. diazotrophicus strains to grow at different nitrogen concentrations was explored in two in vitro conditions: one using LGI solid Plates8 supplemented with different concentrations of NH4NO3 and the other using semisolid LGI medium8 supplemented with different concentrations of NH4NO3 (Sigma–Aldrich A3795). In the first case, bacteria were grown until stationary phase (five independent growth tubes by strain) in MESMA liquid medium14 for 48h at 30°C and 200rpm. Cells were washed twice by centrifugation and resuspended in MgSO4 10mM (Sigma–Aldrich M7506). Each bacterial suspension was serially diluted (factor 1:10) and dilutions were placed in plates at different NH4NO3 concentrations. Bacterial population was quantified by the DPSM method10. For the second condition, bacterial strains were grown until stationary phase (five independent growth tubes by strain), in 150ml of MESMA liquid medium, for 48h at 30°C and 200rpm. Each washed bacterial suspension was serially diluted (factor 1:10) and 100μl of each dilution were placed in series of semisolid LGI tubes in triplicate25 containing the amount of NH4NO3 assessed (data observed in Tables 1 and 2). Quantification was carried out by the most probable number method (MPN) using a McCrady table with three replicate vials for each dilution.
Bacterial number of three strains of G. diazotrophicus grown in solid LGI media supplemented with different NH4NO3 concentrations
| [NH4NO3] mM | Log CFU/ml (SD) | ||
|---|---|---|---|
| PAl 5T | PAl 3 | UAP 5560 | |
| 0 | 8.43 (±0.63)A | 9.00 (±0.62)A | 9.09 (±0.30)A |
| 0.35 | 8.79 (±0.17)A | 9.09 (±0.35)A | 9.16 (±0.15)A |
| 0.7 | 9.05 (±0.31)A | 9.09 (±0.36)A | 8.79 (±0.17)A |
| 1.4 | 9.31 (±0.15)A | 9.44 (±0.08)A | 8.89 (±0.17)A |
| 2.8 | 9.01 (±0.27)A | 9.20 (±0.17)A | 9.16 (±0.27)A |
| 5.6 | 8.95 (±0.24)A | 9.31 (±0.28)A | 8.95 (±0.24)A |
| 11.2 | 9.29 (±0.35)A | 9.33 (±0.05)A | 9.05 (±0.35)A |
| 22.4 | 8.72 (±0.30)A | 9.41 (±0.12)A | 8.85 (±0.11)A |
| 44.8 | 0 C | 0 C | 5.02 (±0.25)B |
| 89.6 | 0 C | 0 C | 5.07 (±0.05)B |
| 179.2 | 0 C | 0 C | 0 C |
Values correspond to the media of five independent samples determined by the DPSM method. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. SD: standard deviation.
Bacterial number of three strains of G. diazotrophicus grown in semisolid LGI media supplemented with different NH4NO3 concentrations
| [NH4NO3] mM | Log of cell number/ml (SD) | ||
|---|---|---|---|
| PAl 5T | PAl 3 | UAP 5560 | |
| 0 | 8.57 (±0.43)A | 8.51 (±0.43)A | 8.49 (±0.30)A |
| 0.35 | 8.43 (±0.37)A | 8.47 (±0.35)A | 8.27 (±0.13)A |
| 0.7 | 8.43 (±0.31)A | 8.44 (±0.41)A | 8.43 (±0.31)A |
| 1.4 | 8.46 (±0.35)A | 8.29 (±0.36)A | 8.28 (±0.32)A |
| 2.8 | 8.40 (±0.27)A | 8.33 (±0.38)A | 8.21 (±0.42)A |
| 5.6 | 8.38 (±0.34)A | 8.26 (±0.37)A | 8.25 (±0.33)A |
| 11.2 | 8.32 (±0.35)A | 8.13 (±0.25)A | 8.13 (±0.38)A |
| 22.4 | 8.32 (±0.44)A | 8.17 (±0.32)A | 8.15 (±0.40)A |
| 44.8 | 2.56 (±0.32)B | 2.62 (±0.44)B | 2.57 (±0.35)B |
| 89.6 | 0 D | 0 D | 1.03 (±0.45)C |
| 179.2 | 0 D | 0 D | 0 D |
Values correspond to the media of five independent samples determined by the MPN method using a McCrady table with three replicate vials for all dilutions of each sample. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. SD: standard deviation.
The effect of NH4NO3 on bacterial association with sugarcane was assessed with the use of split root experiments. For this purpose, sugarcane plantlets variety MEX 57-473 were obtained by micropropagation as described previously25. Micropropagated plantlets were free from bacteria. Forty plantlets were inoculated with G. diazotrophicus PAl 5T strain by immersion of roots for 1h in the bacterial suspension (5×108UFC/ml). Forty plantlets were used as non-inoculated controls and were only immersed in distilled sterile water. To obtain the bacterial suspension, G. diazotrophicus PAl 5T was grown in 10 flasks containing 35ml of TESMA medium until stationary phase, cells were washed by centrifugation twice; after, the pellet was resuspended in 35ml of MgSO4 (10mM) and mixed to obtain 350ml of bacterial suspension. Ten milliliters of this suspension were dispensed in tubes of 25cm×2.5cm and a single tube was used to inoculate each sugarcane plantlet.
After inoculation, both inoculated and non-inoculated plantlets were placed in sterile split root systems (Fig. 1), each consisting of two pots joined by the upper part. Each pot had 500ml capacity and contained sterile vermiculite. For each plant, half of the roots were placed in pot 1 and the other half in pot 2, approximately 3 roots in each pot. The split root systems were watered in both pots with enough water and low doses of NH4NO3 (10mgN/plant equivalent to 0.35mM) and mineral salts, according to Muñoz-Rojas and Caballero-Mellado25. The pots were covered with aluminum foil, and the zone where the plants emerged was protected with sterile cotton. The plantlets were maintained under greenhouse conditions with controlled temperature (26–30°C) with a light/dark photoperiod of 16/8h. Twenty days after inoculation (dpi) the bacterial number was determined for five plants both in their rhizospheres and inside the roots. At 20dpi, 15 plants of each treatment (inoculated and non-inoculated) were supplemented with a high dose of NH4NO3 in one of the pots (180mg of N/plant equivalent to 6.3mM) (Fig. 1) under sterile conditions. Plants were again maintained under greenhouse conditions and watered periodically with distilled sterile water. Rhizospheric and endophytic bacteria were recovered from the two pots for each plant system and the population was determined at 35, 55 and 100dpi, equivalent to 15, 35 and 80 days post fertilization (dpf). Bacterial number was determined as described previously by the most probable number method using a McCrady table with three replicate vials for each dilution25. For this purpose, five independent plants or systems for each treatment (inoculated and non-inoculated) and treatments at different nitrogen levels were analyzed in each time. The plants were carefully removed from the vermiculite, each side of the root system was placed in an independent sterile container, and the root was shaken to discard vermiculite that was not adhered. The resultant root-vermiculite was submerged in enough sterile water (covering the root) and vortexed to maximal velocity for 40s: the suspension was used to perform rhizospheric bacterial quantification. Vermiculite weight was obtained by drying samples without the roots. Furthermore, for endophytic bacteria quantification, each root was placed in a sterile bottle, washed to discard vermiculite and disinfected with 70% ethanol for 30s. Then the roots were rinsed with distilled water and the surface was sterilized with a 1.5% sodium hypochlorite solution (Sigma–Aldrich 425044) for 20min. Later, the roots were rinsed six times with sterile distilled water under sterile conditions. Fresh roots were macerated in water in 1:10 (w/v) proportion. Each sample used for bacterial quantification was diluted (factor 1:10) until dilution 1:1,000,000 and after, 100μl of each dilution were placed in a tube containing semisolid LGI medium (without nitrogen for the growth of diazotrophic bacteria), three tubes were dispensed. After bacterial growth, positive and negative tubes were registered; the presence of a yellow pellicle at the top of the LGI semisolid medium, was recorded as a positive tube8,25,27. In addition, acidification of the media was observed with color changes from green to yellow, which is characteristic of G. diazotrophic growth; furthermore, after 7 days the yellow color was absorbed by bacteria8. The estimation of the bacterial number of each sample was carried out by the most probable number method using a McCrady table with three replicate vials for each dilution (with a confidence limit of 95%). For rhizospheric bacteria quantification, the value obtained from the McCrady table was multiplied per 10 and the dilution factor was considered in order to obtain the number of bacterial cells/ml of liquid suspension (sample). This value was multiplied per the initial water volume where the root was vortexed and divided by the amount in grams (g) of vermiculite (V) present in the suspension (considered as the adhered soil to the roots)25. The final quantified values obtained were stated as the number of cells/gV. To assess endophytic bacteria quantification, the value obtained from the McCrady table, was multiplied by 10 and the dilution factor was considered in order to obtain the number of bacterial cells/ml of liquid suspension (sample); later this value was multiplied by 10 due to initial dilution (w/v) of fresh roots for each side of the root systems. Each bacterial number value obtained was transformed to logarithmic form for statistical purposes. All treatments explored in the present work had five bacterial number values which were used to calculate the standard deviation and the statistical analysis. To ensure that quantified bacteria corresponded to G. diazotrophicus PAl 5T, its ability to inhibit a sensitive strain (PAl 3) was checked and electrophoretic mobility patterns of 12 metabolic enzymes were compared with a reference strain6,7,17. To achieve this goal, some positive tubes of semisolid LGI media with characteristic growth of G. diazortrophicus, were used to streak the bacteria pellicle on solid plates of LGI media and selected colonies were assessed for their ability to inhibit a sensitive strain by the double agar layer method20,26. All selected isolates were able to inhibit the growth of G. diazotrophicus PAl 3 (an antagonistic characteristic of strain PAl 5T), and they also had the same pattern of electrophoretic mobility of the metabolic enzymes explored, as the reference strain G. diazotrophicus PAl 5T (data not shown).
. After G. diazotrophicus inoculation, the root of each plant was divided and placed in 2 pots. At 20dpi half of the plants were supplemented with high doses of NH4NO3 on one side of the system. Images from plants in the split root experiment (B), in the greenhouse (C) and one plant at 35dpi before the bacterial count (D).")
Scheme representing the system used for the split root experiments (A). After G. diazotrophicus inoculation, the root of each plant was divided and placed in 2 pots. At 20dpi half of the plants were supplemented with high doses of NH4NO3 on one side of the system. Images from plants in the split root experiment (B), in the greenhouse (C) and one plant at 35dpi before the bacterial count (D).
Data corresponding to each treatment for the different experiments were statistically compared in pairs with the t-Student test, using Sigma Plot of the Jandel Scientific Software. Results of comparison were used to generate a matrix of differences and similarities between treatments for assignment of letters (data not shown).
ResultsEffect of NH4NO3 on G. diazotrophicus strains in vitroThe three strains of G. diazotrophicus explored (PAl 5T, PAl 3 and UAP 5560) were able to grow in solid media until 22.4mM of NH4NO3 (Table 1), which is a high nitrogen level and corresponds to 640mg of N/plant. The statistical analysis showed no differences between bacteria grown in the presence of 22.4mM of NH4NO3 in comparison with normal LGI. G. diazotrophicus UAP 5560 tolerated a concentration of 89.6mM in solid LGI, but the bacterial number was reduced from 108 to 105CFU/ml (Table 1). In semisolid LGI medium it was also observed that the three strains of G. diazotrophicus explored were able to grow until 22.4mM of NH4NO3 (Table 2); the growth of the strains was affected at 44.8mM in the order of 102cells/ml and the growth of UAP 5560 also tolerated better than others the presence of high levels of NH4NO3 (89.6mM); however, under this condition bacterial numbers diminished in the order of 101cells/ml.
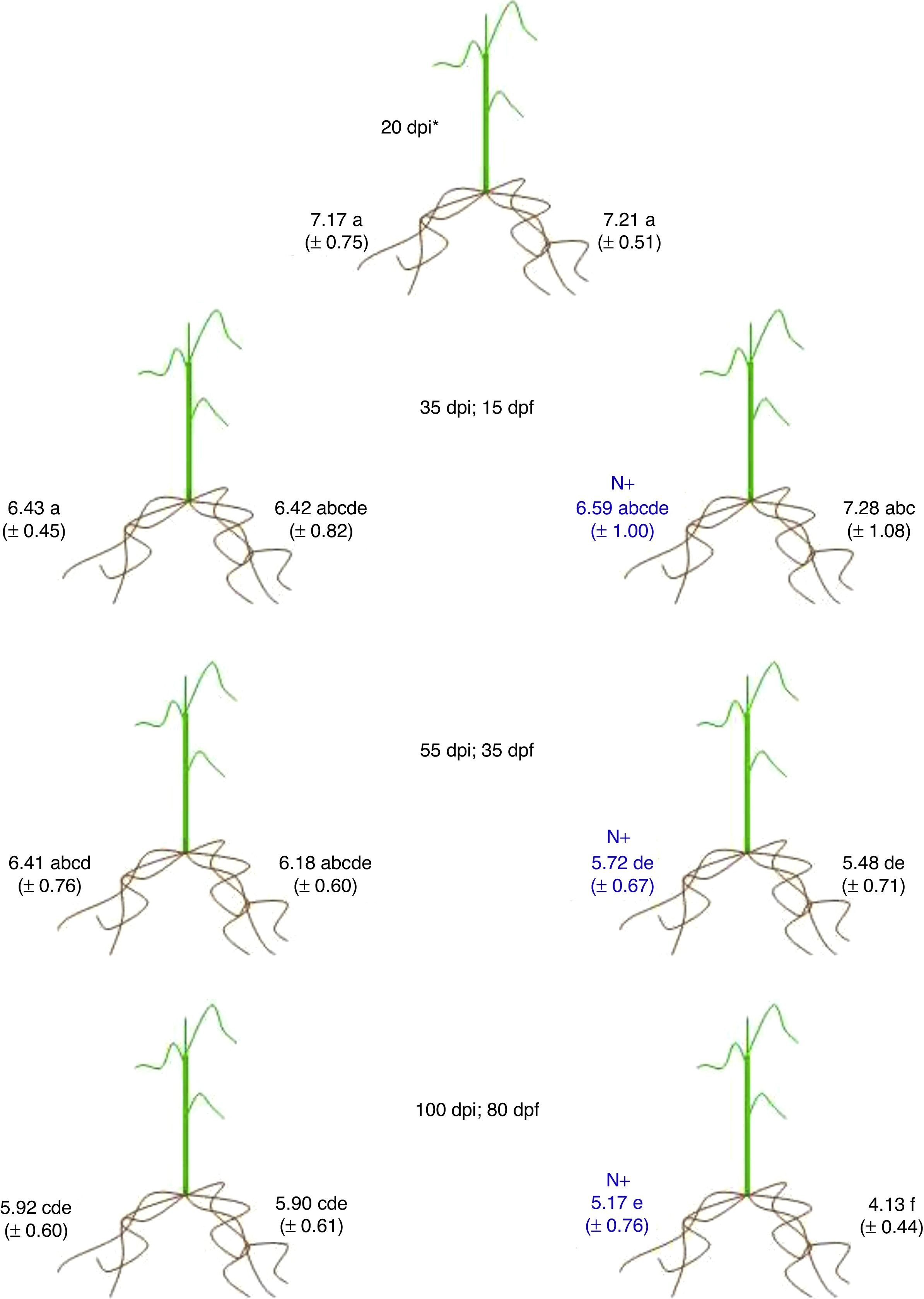
Effect of NH4NO3 on the colonization of sugarcane testing G. diazotrophicus PAl 5T in split root experimentsThe analysis of the population of G. diazotrophicus strain PAl 5T was carried out using the sugarcane variety MEX-57473 with split root experiments both in rhizospheres as endophytically. Bacteria were not detected in non-inoculated control plants. Rhizospheric population was similar in both sides of the root systems at 20dpi, about 1×107cells/g vermiculite (V) when basal levels of NH4NO3 were present. No statistical differences were observed at 35dpi (15dpf) in the rhizospheric population between both sides of the system, neither between treatments fertilized with high levels of NH4NO3 in comparison to those fertilized with basal levels. Furthermore, there were no differences when comparing the same treatments 20dpi (Fig. 2). In accordance with our data, bacterial rhizospheric population decreased at 55dpi (35dpf) in plants fertilized with high levels of NH4NO3 when compared to the initial population observed in plants at 20dpi. However, no differences were observed between the bacterial numbers recovered from each side of the split root systems neither for the plants fertilized with high levels of NH4NO3 nor for the plants treated with basal levels. Interestingly, at 100dpi (80dpf), fertilized plants with high levels of nitrogen had differences in rhizospheric population, detecting low bacterial numbers (around 1×104cells/gV) in the pot fertilized with basal nitrogen in comparison with the bacterial numbers detected in the other pot of the system and also when compared to the bacterial numbers of both pots from plants fertilized with low levels of nitrogen, in the order of 105cells/gV (Fig. 2).
 with the respective standard deviation. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. dpi: days post inoculation; dpf: days post fertilization; N+: addition of 180mg of nitrogen/plant.")
Rhizospheric bacterial populations in split root experiments. Each value represents the media of data for five independent plants (Log of cell number/gV) with the respective standard deviation. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. dpi: days post inoculation; dpf: days post fertilization; N+: addition of 180mg of nitrogen/plant.
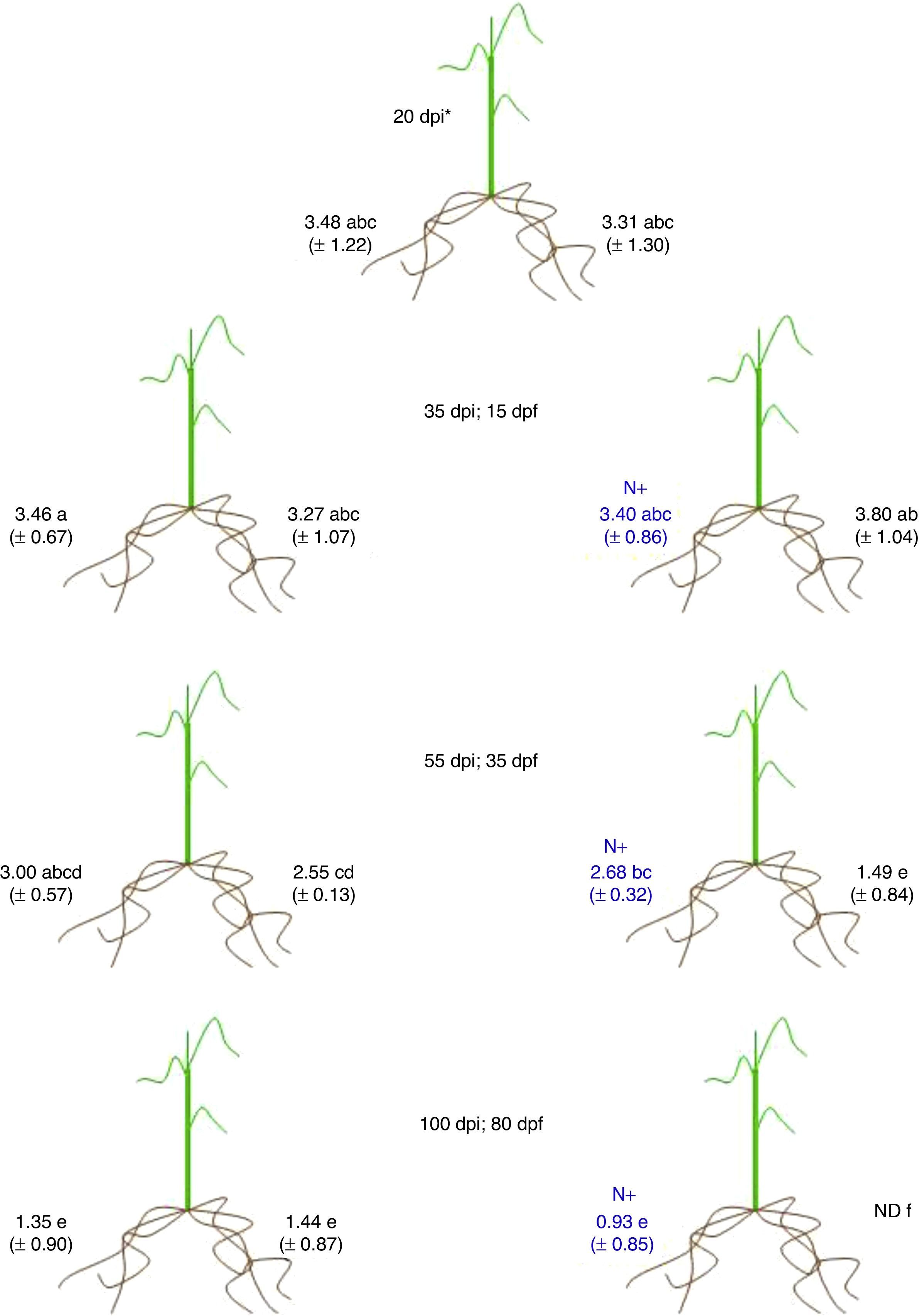
The effect of nitrogen on the endophytic population of G. diazotrophicus PAl 5T was more evident (Fig. 3). Inside the roots, the population of G. diazotrophicus was detected in the order of 103cells/g of root at 20dpi. After the addition of high levels of NH4NO3 in one pot of the split root systems corresponding to the treated plants containing high levels of nitrogen, no changes were observed in the bacterial population measured 35dpi (15dpf) when comparing pots of the same system or systems with basal nitrogen. However, 55dpi (35dpf), the bacterial population of roots from pots with basal nitrogen showed a decrease (around 50cells/g of root) in comparison with the bacterial population of roots from pots added with high level of NH4NO3 (around 6×102cells/g of root) of the same plant system, and also when compared to the bacterial numbers observed in roots from plants fertilized with low levels of nitrogen (Fig. 3). Similar results were observed 100dpi (80dpf) in plants fertilized with high levels of nitrogen. In this case, the population of G. diazotrophicus was not detected inside the roots from pots fertilized with basal levels of NH4NO3, but bacteria were detected in roots from pots fertilized with high levels of nitrogen (around 10cells/g of root) of the same split root system. These fertilized pots are statistically similar to the bacterial numbers detected in pots from plants fertilized with basal levels of nitrogen (around 40cells/g of root) (Fig. 3).
 with the respective standard deviation. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. dpi: days post inoculation; dpf: days post fertilization; ND: not detected; N+: addition of 180mg of nitrogen/plant.")
Bacterial population inside roots in split root experiments. Each value represents the media of data for five independent plants (Log of cell number/g root) with the respective standard deviation. Mean values with equal letters are not statistically different at p≤0.05, using the t-Student test. dpi: days post inoculation; dpf: days post fertilization; ND: not detected; N+: addition of 180mg of nitrogen/plant.
Plants require nitrogen for their development. The addition of this component produces key molecules that increase plant growth or the accumulation of metabolic compounds2,5,11. Rhizospheric bacteria population could be modified in response to nitrogen fertilization. It has been shown that ammonium nutrition increased root colonization by Pseudomonas fluorescens 2-79RLI at the root tip and in the lateral root zone when the pH of the nutrient solution was allowed to change according to the nitrogen form provided21. In contrast, the population of G. diazotrophicus associated with sugar cane diminishes after nitrogen fertilization14,25, regardless of the form of nitrogen supplemented22. This decrease could be explained by pleomorphic changes observed in G. diazotrophic cells when they grow in the presence of high levels of NH4NO3 (25mM)29. However, in accordance with this study, bacterial cells survive under this nitrogen concentration in culture media. On the other hand, it has also been suggested that the reduction in the number of bacterial cells associated with sugarcane occurs by stimulating changes in plant physiology after nitrogen fertilization14,31; however, this hypothesis has not been confirmed yet. In this work, the survival of G. diazotrophicus was influenced by 44.8mM of NH4NO3 (equivalent to 640mgN/plant) irrespective of the genotype of G. diazotrophicus used (in vitro experiments). However, strain UAP5560 was more tolerant to nitrogen concentrations, showing survival with a decreased number of cells. Nitrogen concentration affecting bacterial survival (44.8mM) was higher than the level of nitrogen that affects the population associated with sugarcane (6.3mM equivalents to 180mgN/plant). Based on the results presented in this work and data previously published25, it was reasonable to propose that nitrogen fertilization induces changes in the physiology of plants that prevent G. diazotrophicus colonization. To verify this hypothesis, we carried out split root experiments to evaluate the effect of high levels of NH4NO3 when applied on one side of system, while the population of G. diazotrophicus was measured on the other side. For this experiment, strain UAP5560 was applied to the sugarcane variety MEX 57-473, given that this interaction is very stable according to previous data25. As we had expected, G. diazotrophicus population diminished in the plants fertilized with high levels of NH4NO3. This decrease was more evident inside the roots exposed to basal levels of nitrogen, in comparison with the roots fertilized with high nitrogen levels, both in the same plant system. Moreover, this decrease in bacterial population was also evident in plant systems fertilized with low levels in both pots. These observations suggest that the effect of NH4NO3 on the population of G. diazotrophicus occurs through systemic changes in the plant, affecting the establishment of G. diazotrophicus in the roots on the other side. This observation was more evident 100dpi or 80dpf, when a decrease in the population of G. diazotrophicus was observed, due to the age of the plant25. Supplementing high levels of NH4NO3 to sugarcane plants induce a decrease in the sucrose content in stalks in early growth24 and sucrose has been proposed as the principal carbon source to G. diazotrophicus12. Furthermore, NH4NO3 produces changes in the components of the apoplastic sap of sugarcane plants, including aminoacids, proteins, and sugars38. Those changes could increase during plant growth and could be related to bacterial diminution. Moreover, exopolysaccharide production is required for biofilm formation and plant colonization by G. diazotrophicus23, and the changes occurring in the plant could inhibit biofilm formation and bacteria establishment. Finally, some chemical compounds are responsible for inducing a resistance to disease in plants30, but could also induce a systemic resistance (ISR) similar to that produced by rhizobacteria39,41, making it conceivable that NH4NO3 could elicit ISR and prevent the colonization of G. diazotrophicus in sugarcane16. The nitrogen effect over the population of G. diazotrophicus is more evident inside the plant than in the rhizosphere (Figs. 2 and 3). This could be due to physiological changes occurring inside the plant and directly affecting the bacterial population; however, in the rhizosphere, firstly the metabolites have to be exported to provoke changes in the environment.
Taken together, the results in this study show that the decrease of G. diazotrophicus associated with sugarcane occurs due to changes in the physiology of the plant rather than by the direct effect that NH4NO3 could exert on bacterial cells.
Ethical disclosuresProtection of human and animal subjectsThe authors declare that no experiments were performed on humans or animals for this study.
Confidentiality of dataThe authors declare that no patient data appear in this article.
Right to privacy and informed consentThe authors declare that no patient data appear in this article.
Conflict of interestThe authors declare that they have no conflicts of interest.
We are grateful to CONACYT (000000000156576), PRODEP and VIEP-BUAP-2014 for the support of this work. Osvaldo Rodríguez-Andrade and Dalia Molina Romero were awarded a CONACYT fellowship. This work is dedicated to the memory of Jesús Caballero-Mellado.
