




Los linfocitos B (LB) se consideran el centro de la desregulación inmune en pacientes con lupus eritematoso sistémico (LES), principalmente, por su producción de autoanticuerpos. Recientemente, se demostró la existencia de LB, incluidos en los B transicionales, con capacidad reguladora (Breg) y fenotipo CD19+CD24hiCD38hi. En humanos se demostró la importancia de CD80 y CD86 en su función reguladora. El papel de CD1d aún no ha sido evaluado.
ObjetivoEvaluar la frecuencia de LB maduros, memoria y transicionales, en controles y pacientes con LES, además de la expresión de CD1d y correlacionarla con la actividad de la enfermedad medida por SLEDAI (Systemic Lupus Erythematosus Disease Activity Index).
Materiales y métodosSe evaluó por citometría de flujo la frecuencia de subpoblaciones de LB basados en la expresión de CD19, CD24 y CD38, además de CD1d, en controles con otras enfermedades autoinmunes (OEA), individuos sanos y pacientes con LES, y se correlacionó con SLEDAI.
ResultadosSe evidenció una disminución significativa en el porcentaje de LB de memoria en pacientes LES y OEA, sin alteraciones en las subpoblaciones de LB maduros y transicionales. La expresión de CD1d no evidenció diferencias significativas en ninguna de las subpoblaciones ni se correlacionó con SLEDAI.
ConclusiónLa disminución de la subpoblación de memoria fue previamente descrita en LES y se ha asociado a algunos tipos de tratamiento. Aunque CD1d se ha asociado a la función de Breg en murinos, no hubo diferencias significativas en su expresión en las subpoblaciones y queda por clarificar su papel en la función de las Breg humanas.
B lymphocytes are considered the center of immune dysregulation in Systemic Lupus Erythematosus (SLE). It has recently been demonstrated that there is a B cell with regulatory capacities (Breg) included in transitional B lymphocytes with the phenotype CD19+CD24hiCD38hi. The importance of CD80 and CD86 in the regulatory function of the Bregs has been demonstrated in humans, but the role of CD1d has not been evaluated.
ObjectiveTo evaluate the frequency of mature, memory and transitional B cells in SLE patients and controls, the expression of CD1d among these cells, and its correlation with the activity of the disease measured using the Systemic Lupus Erythematosus Disease Activity Index (SLEDAI).
Materials and methodsThe frequency of the B cell subsets was evaluated by flow cytometry based on the expression of CD19, CD24 and CD38, as well as CD1d in these cells in SLE patients and controls, and were correlated with the activity of the disease measured using the SLEDAI.
ResultsA significant reduction in the percentage of memory B cells was observed in SLE patients and other autoimmune conditions, with no changes in the mature or transitional B cell subsets. Similarly, no significant differences were observed in the expression of CD1d in any of the subsets, nor was there any correlation with the SLEDAI.
ConclusionThe reduction of the memory subset has been previously described in SLE, and has been associated with some types of treatment. The expression of CD1d in all the subsets was observed, but its role in the regulatory function of the CD19+CD24hiCD38hi cells is still not clear.
Típicamente, las células B han sido descritas como elementos fundamentales en la patogénesis de muchas enfermedades autoinmunes, como el Lupus Eritematoso Sistémico (LES), debido a la producción de autoanticuerpos dirigidos contra diferentes antígenos solubles y celulares, observándose una correlación positiva entre el número de células productoras de autoanticuerpos y la actividad de la enfermedad1. Recientemente, se ha reportado la presencia de células B reguladoras (Breg); en diferentes modelos de enfermedades autoinmunes tales como colitis autoinmune2, inflamación intestinal crónica3, encefalitis autoinmune4 y artritis inducida por colágeno5.
El papel de las Breg ha sido estudiado, principalmente, en modelos murinos como el NZB/W, en el cual se describió la presencia de células B IL-10+CD19+CD5+CD1dhi, denominadas B10, las cuales poseen actividad inhibitoria de la inflamación6,7. Ratones NZB/W deficientes de células B (CD19−/−), presentan una glomerulonefritis más severa, acompañada de un incremento en la tasa de mortalidad y ausencia de la subpoblación reguladora B10 CD1dhiCD5+ respecto a ratones silvestres NZB/W CD19+/+, los cuales aumentan el número de células B10 durante el transcurso de la enfermedad. Estos hallazgos pudieran explicarse por la presencia de células B10 en los ratones tipo silvestre NZB/W CD19+/+ comparados con los ratones CD19−/−8.
Además de las células B10, se ha descrito que la subpoblación de células B transicionales precursoras de zona marginal (T2-MZP), contienen por lo menos un subgrupo de células B con capacidad reguladora. La transferencia adoptiva de células B T2-MZP previamente estimuladas con agonista de CD40 a ratones MRL/lpr con lupus, indujo una remisión de los síntomas clínicos, disminución de la actividad de la enfermedad y del daño renal9, lo cual sugiere que estas células T2-MZP presentan una actividad reguladora la cual depende de la molécula CD40.
Dado que las células transicionales CD19+CD24hiCD38hi se han descrito como los análogos en sangre periférica de las células T2 en humanos10,11, se ha sugerido que esta subpoblación de células B pudiera contener a las células B reguladoras. Un estudio realizado en pacientes con LES, demostró la presencia de un subgrupo de células B CD19+CD24hiCD38hi que adquiría capacidad reguladora luego de la estimulación por CD40, suprimiendo la diferenciación de las células T efectoras CD4+ hacia una respuesta Th1, mediante un mecanismo dependiente de IL-10. Además, se observó que la frecuencia de estas células en sangre periférica de pacientes con LES estaba aumentada pero estas fueron refractarias a la estimulación por CD40 produciendo menos IL-10, lo cual reducía su capacidad reguladora12.
Además de la importancia del CD40, en la función reguladora ejercida por las células CD19+CD24hiCD38, se ha descrito la participación de moléculas coestimuladoras como CD80 y CD8612,13.
Paralelamente, en un modelo murino de enfermedad intestinal crónica, se describió la importancia de la molécula CD1d en la función reguladora de las células B, ya que se pudo evidenciar que la transferencia de células B deficientes de CD1d permitió la progresión de la enfermedad, contrario a lo observado luego de la transferencia de células B tipo silvestre, evidenciando la existencia de una subpoblación de células B localizada en el tejido linfoide asociado al intestino, la cual, bajo un ambiente inflamatorio, se caracteriza por la regulación positiva de CD1d, además de la producción de IL-10 con capacidad de suprimir la inflamación, por medio de la regulación negativa de la cascada de señalización de la IL-1 y la activación del factor de transcripción STAT3.
Otros estudios también han demostrado la importancia de moléculas como CD1d o CD19 en la acción reguladora ejercida por las células B. Deficiencias en la expresión de estas moléculas, provocan una reducción significativa de la subpoblación de células B productoras de IL-10, lo cual sugiere que CD1d y CD19 son de gran importancia para el desarrollo de LB y para su función reguladora medida por la producción de IL-106.
La regulación mediada por CD1d podría explicarse por una interacción de las células que la expresan con las iNKT, subtipo de células NK con aparente capacidad reguladora, como se ha demostrado en algunos modelos de enfermedades autoinmunes14. Se conoce también que la porción intracelular de CD1d está asociada a la cascada de señalización de la IL-1015.
Por ende, es necesario caracterizar no solo las subpoblaciones de células Breg presentes en pacientes con LES, sino también evaluar la expresión de marcadores como el CD1d en las células B reguladoras, lo cual ayudaría a explicar las alteraciones funcionales previamente descritas en esta subpoblación de células en pacientes con LES y si estas son explicables por diferencias en las frecuencias de las subpoblaciones de células B o por la expresión diferencial de CD1d como molécula moduladora de la función de las Breg, lo que a su vez podría derivar en nuevos blancos terapéuticos.
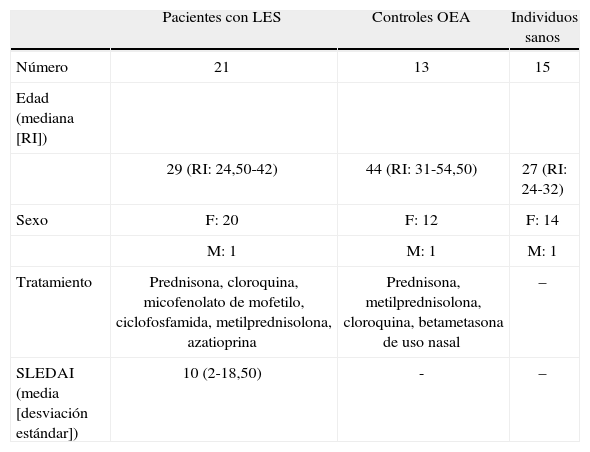
Materiales y métodosPacientes y controlesSe llevó a cabo, entonces, un estudio de tipo cuasi experimental de corte transversal, en el cual el tamaño muestral se decidió a conveniencia. Se incluyó un grupo de 21 pacientes diagnosticados con LES de acuerdo con los criterios del Colegio Americano de Reumatología (ACR)16 y se clasificaron de acuerdo al índice de actividad de la enfermedad, en actividad severa (≥4) y moderada o baja (<4) de acuerdo con la calificación dada por el SLEDAI (Systemic Lupus Erythematosus Disease Activity Index)17.
Además, se incluyó un grupo de controles sanos (n=15) y un grupo de sujetos con otras enfermedades autoinmunes (OEA) diferentes a LES (n=13), tales como esclerosis sistémica6, artritis reumatoide3, artritis psoriásica1, síndrome de Sjögren1, poliangeítis granulomatosa1 y sarcoidosis1, los cuales recibían tratamiento inmunosupresor similar al de los pacientes con LES, incluyendo esteroides a dosis varias, metotrexate, cilofosfamida y micofenolato por diferentes tiempos, con una mediana de 45 meses de tratamiento inmunosupresor (rango intercuartílico de 1 a 216). Se excluyeron aquellos pacientes o controles que presentaban infecciones al momento de la toma de la muestra y pacientes que presentaban inmunodeficiencias primarias o secundarias, o aquellos sujetos que no firmaran el consentimiento informado.
Tanto los pacientes con LES como los OEA fueron reclutados en el Servicio de Reumatología de la Universidad de Antioquia en el Hospital San Vicente Fundación (tabla 1).
Datos clínicos y demográficos de pacientes con LES, controles OEA e individuos sanos.
| Pacientes con LES | Controles OEA | Individuos sanos | |
| Número | 21 | 13 | 15 |
| Edad (mediana [RI]) | |||
| 29 (RI: 24,50-42) | 44 (RI: 31-54,50) | 27 (RI: 24-32) | |
| Sexo | F: 20 | F: 12 | F: 14 |
| M: 1 | M: 1 | M: 1 | |
| Tratamiento | Prednisona, cloroquina, micofenolato de mofetilo, ciclofosfamida, metilprednisolona, azatioprina | Prednisona, metilprednisolona, cloroquina, betametasona de uso nasal | – |
| SLEDAI (media [desviación estándar]) | 10 (2-18,50) | - | – |
F: femenino; M: masculino.
Para la obtención de las células mononucleares de sangre periférica (PBMC) se les realizó venopunción a pacientes y controles para obtener aproximadamente 8 mililitros de sangre periférica, la cual fue anticoagulada con EDTA. Posteriormente, la muestra se diluyó en una proporción 1:1 en buffer salino fosfatado (PBS) 1X y se llevó a cabo una separación por gradiente de densidad con Ficoll Histopaque 1077 (Sigma), mediante centrifugación por 30 minutos a 800 × g y se realizaron los respectivos lavados y la evaluación de la viabilidad celular con azul de tripano al 0.1%.
Para la tinción extracelular se tomaron 0.5×106 PBMC y se resuspendieron en 50uL de una solución de PBS 1X, 4% suero bovino fetal inactivado (SBFi) y 0.01% de azida de sodio (buffer de bloqueo), por 10 minutos a temperatura ambiente. Posteriormente, se agregó una solución de anticuerpos anti-CD19-Pacific blue (2,5μl) (Clona HIB19), anti-CD24-FITC (2,5μl) (Clona ML5), anti-CD38-PerCP (5μl) (Clona HIT2) y anti-CD1d-PE (Clona 51.1) o su control isotípico (2,5μl) (Clona MG2v-57) (Biolegend Roselle Street, San Diego), y se incubó por 25 minutos a 4°C en oscuridad. Las células fueron fijadas con 2% de paraformaldehído (PFA) durante 20 minutos a 4°C y basados en su marcaje se determinaron 3 subpoblaciones de linfocitos B (LB) por citometría de flujo: células B maduras (CD19+CD24intCD38int), de memoria (CD19+CD24hiCD38−) y transicionales 2 (CD19+CD24hiCD38hi) e igualmente se evaluó la expresión de CD1d en estas subpoblaciones. La lectura de las células se realizó en un citómetro de flujo Facs Canto II (BD Biosciences, San Diego CA).
Análisis estadísticoSe utilizaron pruebas no paramétricas, por lo cual se muestran los datos en medianas y rango intercuartílico (RI). Sin embargo, se realizó prueba de normalidad de los datos (D’Agostino & Pearson).
Para comparar el porcentaje de las diferentes subpoblaciones de células B y el porcentaje de las células positivas para CD1d, así como las diferencias en la expresión de CD1d entre las subpoblaciones celulares de pacientes y controles, se utilizó la prueba de Kruskal Wallis y el pos-test de Dunn's. Para correlacionar la expresión de CD1d y el porcentaje de las diferentes subpoblaciones con la actividad de la enfermedad (SLEDAI), se utilizó la correlación de Spearman.
Los datos de las citometrías se analizaron con el programa FLOWJO 7.6.1. Tree Star y los análisis estadísticos se realizaron con el programa Prisma (GraphPad Versión 5.00, La Jolla, EE.UU.).
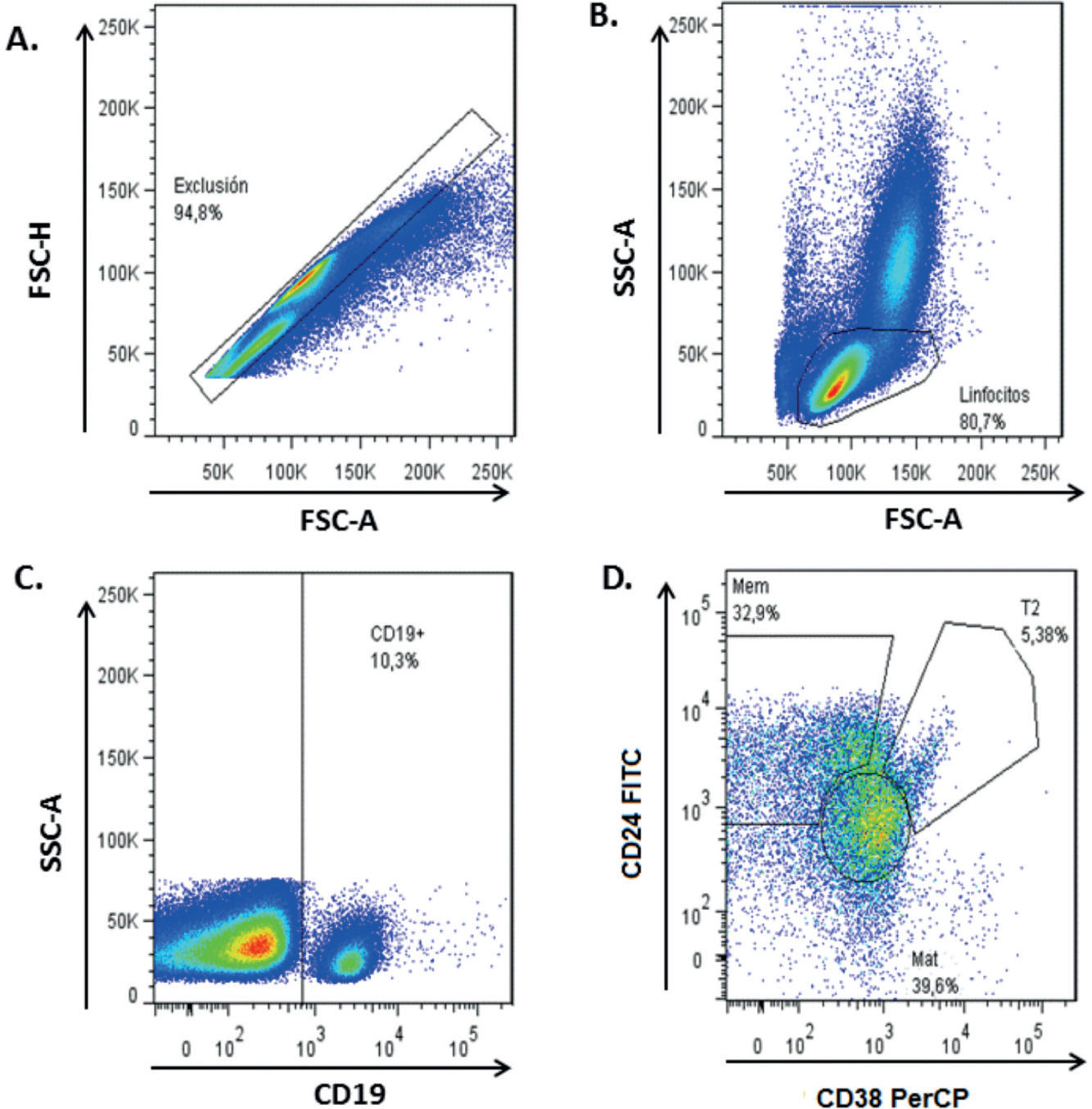
ResultadosFenotipificación de las diferentes subpoblaciones de linfocitos B (LB)Para evaluar la frecuencia de las subpoblaciones de LB de pacientes y controles, se realizó un análisis partiendo de las células positivas para CD19 y dentro de la región CD19+, basados en la expresión diferencial de CD24 y CD38 se definieron las células B maduras (CD19+CD24intCD38int), de memoria (CD19+CD24hiCD38low) y transicionales 2 (T2) (CD19+CD24hiCD38hi) (fig. 1).
, seguido de la determinación de la región de linfocitos por tamaño y granularidad (B) y evaluando así, dentro de esta región, el porcentaje de células CD19+ (C) y dentro de esta región se evaluó la expresión de CD38 y CD24 para determinar así la subpoblación de células B maduras (CD19+CD24intCD38int), de memoria (CD19+CD24hiCD38low) y las T2 (CD19+CD24hiCD38hi) (D).")
Estrategia de análisis de subpoblaciones de linfocitos B por citometría de flujo. El análisis de las subpoblaciones se inició con una exclusión de agregados con los parámetros de área y altura (A), seguido de la determinación de la región de linfocitos por tamaño y granularidad (B) y evaluando así, dentro de esta región, el porcentaje de células CD19+ (C) y dentro de esta región se evaluó la expresión de CD38 y CD24 para determinar así la subpoblación de células B maduras (CD19+CD24intCD38int), de memoria (CD19+CD24hiCD38low) y las T2 (CD19+CD24hiCD38hi) (D).
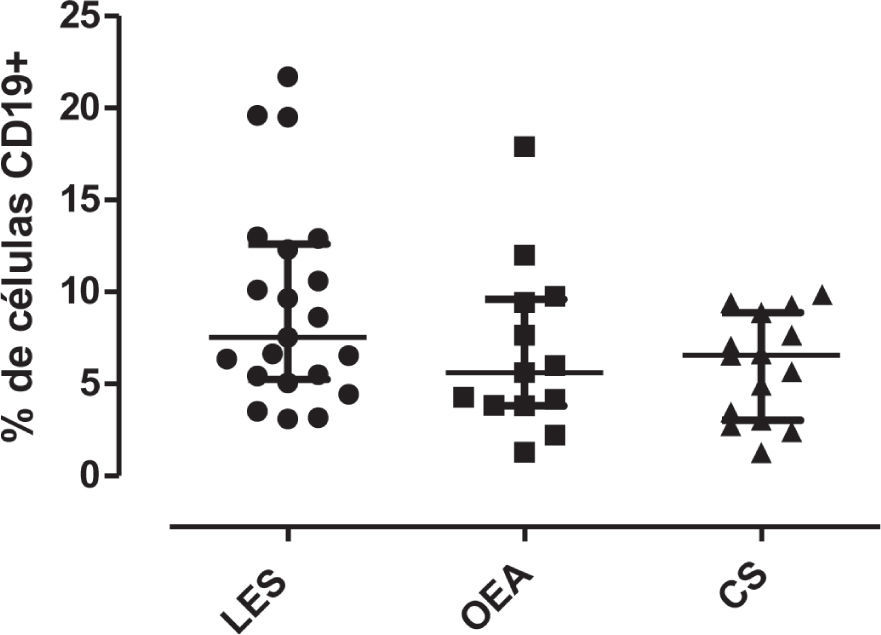
Inicialmente, se evaluó el porcentaje de LB CD19+ en pacientes y controles. No se observaron diferencias estadísticamente significativas en el porcentaje de células CD19+ entre pacientes con LES (mediana: 7.54%, RI: 5.24–12.60), controles con OEA (5.62%, RI: 3.8–9.6) y controles sanos (6.57%, RI: 3.02–8.88) (fig. 2).

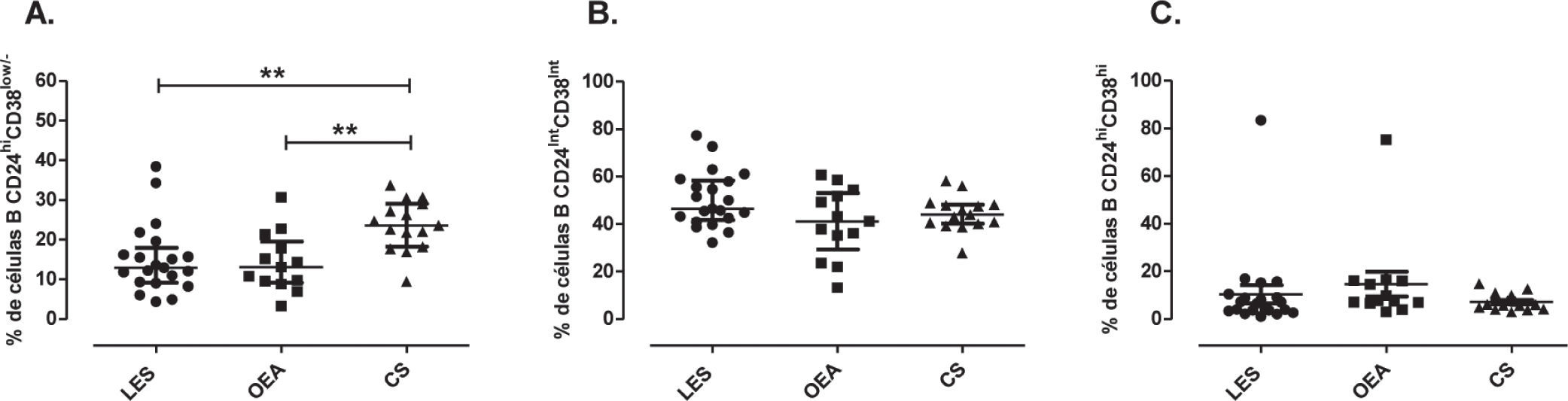
El análisis de las subpoblaciones de linfocitos B evidenció una disminución en el porcentaje de células de memoria en pacientes con LES (Mediana: 12.90%, rango intercuartílico (RI): 9.17–17.90), comparado con controles OEA (13.10%, RI: 9.16–19.55) y controles sanos (23.6%, RI: 18.20–29.0) (p=0.0016) (fig. 3A). No se observaron diferencias significativas en el porcentaje de células B maduras entre controles sanos (44%; RI: 40.10–48.20), pacientes con LES (46.40%; RI: 41.70–58.40) y controles OEA (41.20%; RI: 29.35–53.05) (fig. 3B). De igual forma, el porcentaje de células B transicionales 2 no difirió entre controles sanos (6.09%; RI: 4.10–9.95), pacientes con LES (6.49%; RI: 3.50–9.70) y controles OEA (7.69%; RI: 6.74–16.15) (fig. 3C).
 (A), maduras (CD24intCD38int) (B) transicionales (CD24hiCD38hi) (C) en pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal-Wallis y el post-test de Dunn")
Distribución de subpoblaciones de células B en pacientes y controles. Porcentaje de células B de memoria (CD24hiCd38low/−) (A), maduras (CD24intCD38int) (B) transicionales (CD24hiCD38hi) (C) en pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal-Wallis y el post-test de Dunn's. Se presentan medianas y rangos intercuartílicos (RI) (** P<0.01).
Debido que CD1d ha sido previamente descrito en algunos modelos murinos de autoinmunidad como un marcador fenotípico de las Breg, con papel en la acción reguladora mediada por estas células6,18,19. Se evaluó el porcentaje de células CD1d+ en las tres subpoblaciones de LB evaluadas y su expresión medida como intensidad media de fluorescencia (IMF).
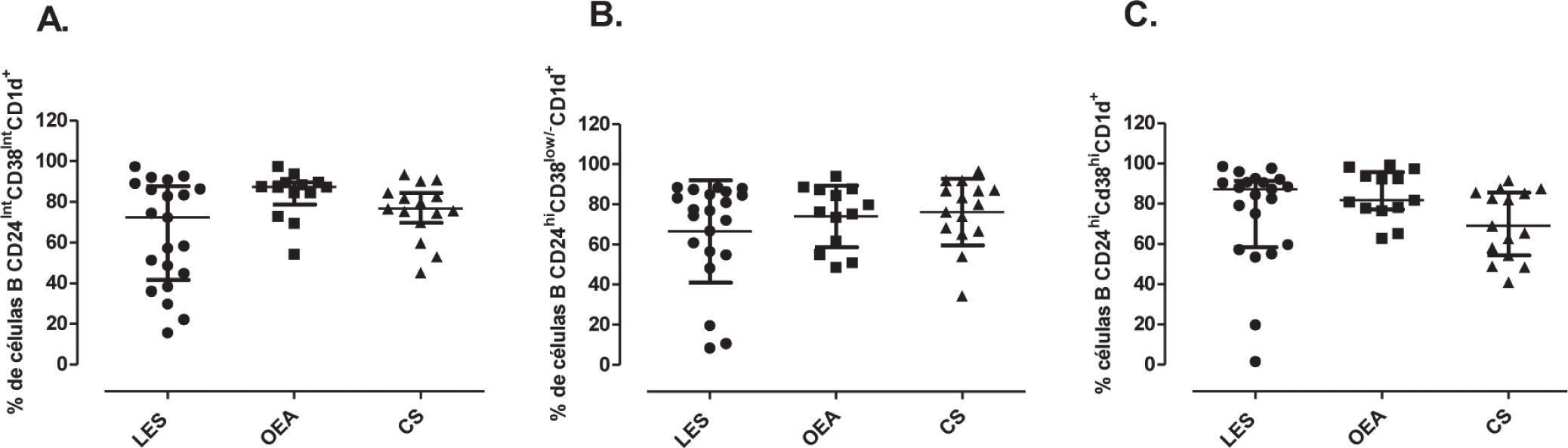
No se observaron diferencias significativas en el porcentaje de LB CD1d+ maduros entre pacientes con LES (72.2%, RI: 41.5–87.7), controles con OEA (87.3%, RI: 78.6–85.7) y controles sanos (76.6%, RI: 69.8–84.5) (fig. 4A). El porcentaje de LB de memoria CD1d+ tampoco mostró diferencias entre pacientes con LES, controles con OEA y controles sanos (76.9%, RI: 55.7–85.7; 76.1%, RI: 58.4–87.6; 79.9%, RI: 66.7–87.1, respectivamente) (fig. 4B). Asimismo, en el porcentaje de LB transicionales 2 CD1d+ no se encontraron diferencias significativas entre pacientes con LES, controles con OEA y controles sanos (87.2%, RI: 58.4–91.3; 81.7%, RI: 77.2–95.6; 68.9%, RI: 54.2–85.5, respectivamente) (fig. 4C).
 (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal Wallis y el post-test de Dunn")
Subpoblaciones de células B CD1d+. Porcentaje de células B CD1d maduras (CD24intCD38int) (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal Wallis y el post-test de Dunn's. Se presentan medianas y rangos intercuartílicos (RI).
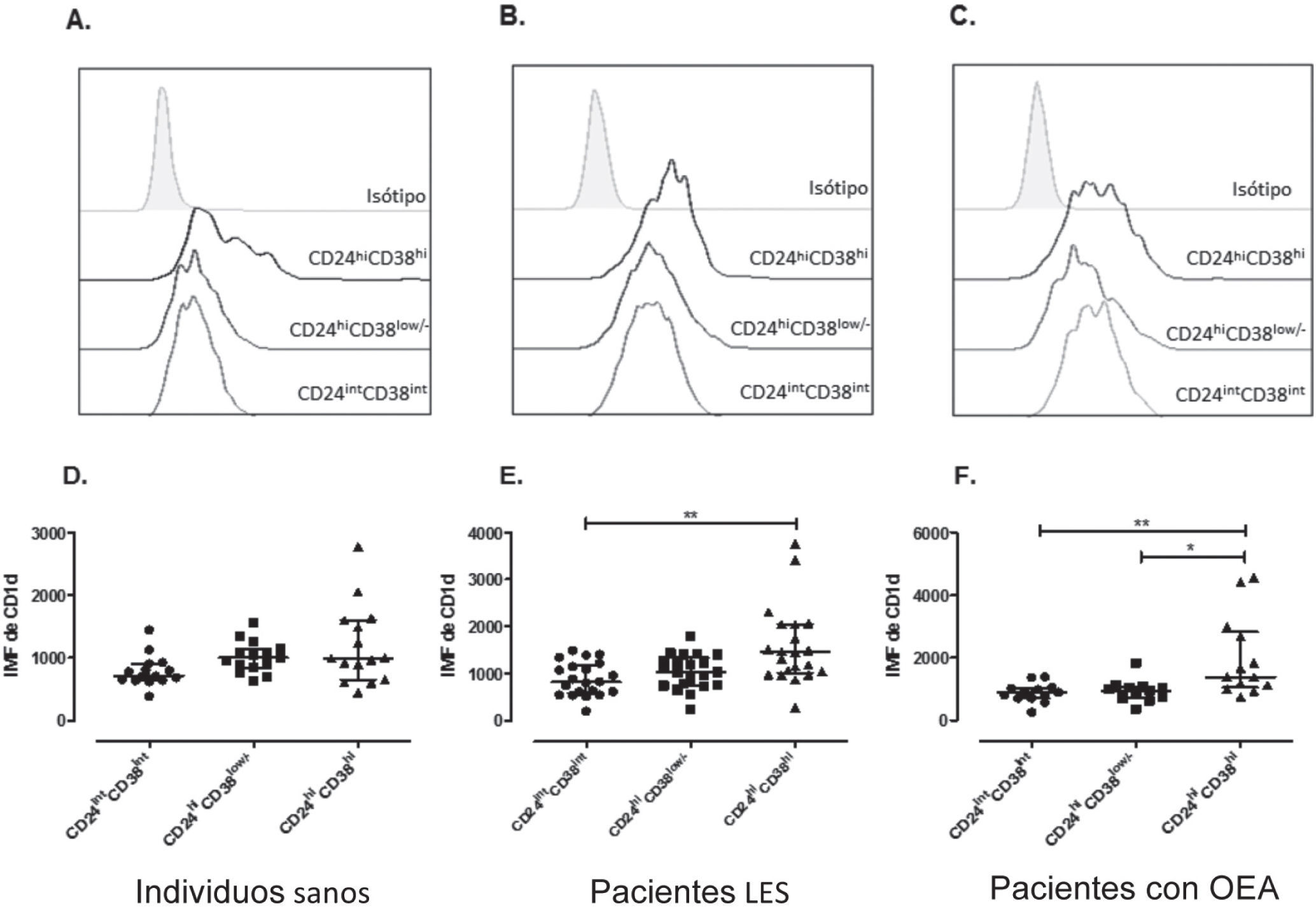
Aunque, al igual de como se había descrito previamente, las células B transicionales CD24hiCD38hi tienen una mayor expresión de CD1d, en este estudio se evidenció un aumento significativo en la expresión de esta molécula en la subpoblación CD24hiCD38hi en los pacientes LES y OEA evaluados20 (fig. 5D-F). Sin embargo, no se observaron diferencias en la IMF de CD1d en los LB maduros entre los pacientes con LES, OEA y controles sanos (IMF LES: 819, RI: 582–1184; IMF OEA: 898, RI: 707–1036; IMF Control Sano: 710, RI: 640–911, respectivamente) (fig. 6A), ni en los LB de memoria de pacientes con LES (IMF: 1026, RI: 748–1336), controles con OEA (IMF: 935, RI: 738–1075) y controles sanos (IMF: 1002, RI: 837–1140) (fig. 6B). De igual forma en la IMF de CD1d de los LB transicionales 2 de pacientes con LES (IMF: 1455, RI: 1010–2046), controles con OEA (IMF: 1394, RI: 1063–2839) y controles sanos (IMF: 994, RI: 648–1601) (fig. 6C).
, de memoria (CD24hiCd38low/−) y transicionales (CD24hiCD38hi), gráfica representativa de un individuo sano (A), un paciente con LES (B), y un control con OEA (C). Intensidad media de fluorescencia (IMF) de CD1d en células B maduras (CD24intCD38int), de memoria (CD24hiCd38low/−) y transicionales (CD24hiCD38hi) de controles sanos (n=15) (D), pacientes con LES (n=21) (E) y controles con OEA (n=13) (F). Estos datos fueron analizados por medio de la prueba de Kruskal-Wallis y el post-test de Dunn")
Expresión de CD1d en las subpoblaciones de células B. Histogramas superpuestos de la expresión de CD1d en células B maduras (CD24intCD38int), de memoria (CD24hiCd38low/−) y transicionales (CD24hiCD38hi), gráfica representativa de un individuo sano (A), un paciente con LES (B), y un control con OEA (C). Intensidad media de fluorescencia (IMF) de CD1d en células B maduras (CD24intCD38int), de memoria (CD24hiCd38low/−) y transicionales (CD24hiCD38hi) de controles sanos (n=15) (D), pacientes con LES (n=21) (E) y controles con OEA (n=13) (F). Estos datos fueron analizados por medio de la prueba de Kruskal-Wallis y el post-test de Dunn's. Se presentan medianas y rangos intercuartílicos (RI) (*P<0.05 **P<0.01).
 de células CD1d en células B maduras (CD24intCD38int) (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal Wallis y el post-test de Dunn")
Expresión de CD1d en subpoblaciones de células B. Intensidad media de fluorescencia (IMF) de células CD1d en células B maduras (CD24intCD38int) (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de Kruskal Wallis y el post-test de Dunn's. Se presentan medianas y rangos intercuartílicos (RI).
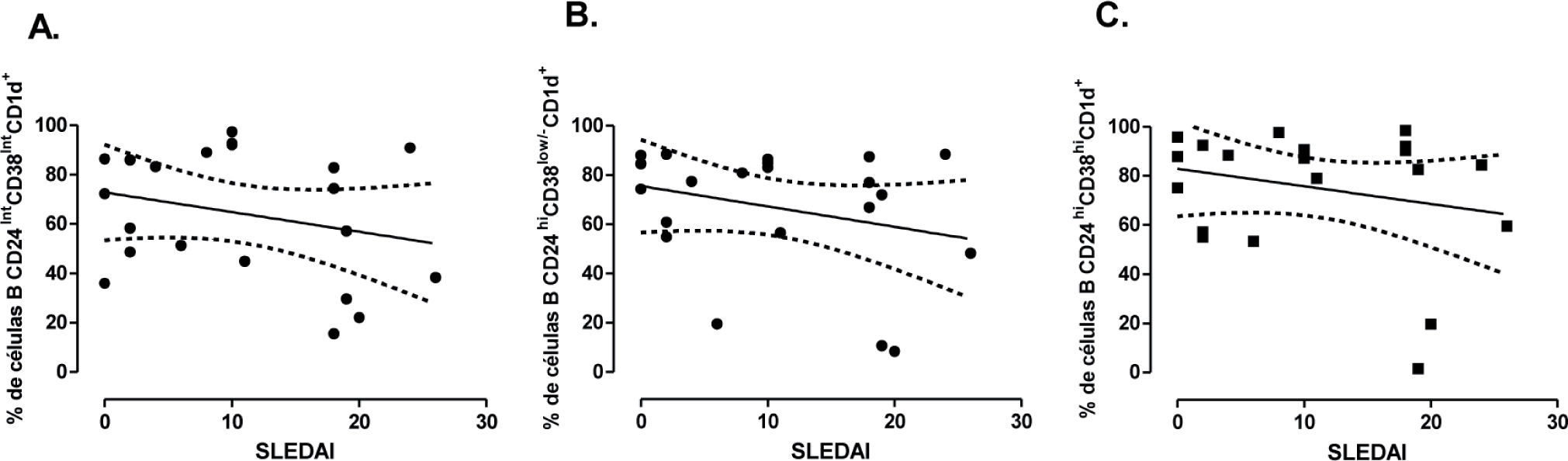
Paralelamente, se quiso evaluar una posible relación entre las subpoblaciones de células B CD1d+ y la actividad del LES. Sin embargo, no se encontró una correlación entre el SLEDAI como medida de la actividad de la enfermedad y el porcentaje de LB CD1d+ maduros (r= −0,213), de memoria (r= −0,274) o transicionales 2 (r= −0,196) ni tampoco se encontró correlación entre la IMF de CD1d de ninguna de las tres subpoblaciones y el SLEDAI (fig. 7).
 (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de correlación de Spearman.")
Correlación del porcentaje de subpoblaciones de LB y la actividad del LES. Correlación entre la actividad de la enfermedad medida por SLEDAI y el porcentaje de células B CD1d+ maduras (CD24intCD38int) (A), de memoria (CD24hiCd38low/−) (B) y transicionales (CD24hiCD38hi) (C) de pacientes con LES (n=21), controles OEA (n=13) y controles sanos (n=15). Estos datos fueron analizados por medio de la prueba de correlación de Spearman.
Muchas investigaciones en los últimos años se han enfocado en la búsqueda de una manera clara para definir el fenotipo que mejor permita diferenciar y clasificar a las células B con capacidad reguladora, convirtiéndose en un reto para esta línea de investigación.
Hasta la fecha se han descrito diferentes fenotipos de células B con capacidad reguladora, entre ellos LB transicionales 2 (T2) CD19+CD24hiCD38hi en humanos, cuyos equivalentes murinos tienen un fenotipo IgMhi IgDhi CD5+CD10+CD20+CD27− CD1dhi, los cuales se han descrito como los análogos de los linfocitos B T2 en humanos12. De igual forma se ha descrito en humanos una subpoblación CD19+CD24hiCD27+ similar a las células B10 murinas, denominadas B10pro, las cuales han sido caracterizadas con los marcadores CD19hi CD38hi CD1dhiCD20hiCD21hiCD23hi CD24hiCD25hiCD27hi21 y posteriormente se describió en sangre periférica en humanos, una subpoblación de células B productoras de altos niveles de IL-10 y TGF-b con un fenotipo CD25hiCD27hiCD86hiCD1dhi22.
La subpoblación de LB CD19+CD24hiCD38hi es una de las subpoblaciones de células Breg que más ha sido estudiada, en especial en enfermedades autoinmunes. Nuestros hallazgos no revelaron diferencias significativas en el porcentaje de células Breg entre pacientes con LES y controles sanos o controles OEA. Esto discrepa de lo observado en el estudio de Londres-Reino Unido realizado por Blair et al., quienes describieron un incremento en la frecuencia de esta subpoblación en pacientes con LES, comparado con controles sanos y pacientes con otras enfermedades autoinmunes12. Sin embargo, resultados similares a los nuestros han sido previamente reportados con otra subpoblación de células Breg.
En cuanto a las otras subpoblaciones evaluadas en este estudio, se observó una disminución en el porcentaje de células B de memoria con fenotipo CD24hiCD38low/− en pacientes con LES y OEA, comparado con controles sanos. Estos resultados concuerdan con los presentados por Blair et al., en 2010, en los cuales se observó una disminución en el porcentaje de células B de memoria CD24hiCD38low/− en pacientes con LES12. Las razones para este hallazgo se han explicado como una anormalidad permanente en el porcentaje de células B de memoria CD27 IgD IgM de los pacientes con LES, además de ser un evento asociado al tratamiento inmunosupresor que reciben23,24. Ahora bien, cabe resaltar que en el estudio del grupo de Blair dentro de sus resultados se muestra esta misma disminución en el porcentaje de células B de memoria12.
La disminución en las células B de memoria también se ha descrito en pacientes con AR en tratamientos con anti-TNF25. Pero es necesario aclarar que ninguno de los pacientes evaluados en este estudio recibía este tipo de tratamiento.
Terapias inmunosupresoras como el micofenolato se han asociado con cambios en las frecuencias de LB en pacientes con LES26. Así mismo, se ha evidenciado una disminución significativa de células B de memoria CD27− en pacientes con LES, la cual es explicada por largos periodos de inmunosupresión con medicamentos como la azatioprina y la prednisolona27,28. No obstante, la frecuencia aumentada de estas células se ha correlacionado positivamente con la actividad de la enfermedad y la presencia de ciertos autoanticuerpos (anti-dsDNA, anti-Smith, anti-RNP y anti-9G4)28. Cabe resaltar que las cinéticas de la repoblación de LB debido a las terapias inmunosupresoras que reciben los pacientes altera las frecuencias de las subpoblaciones de LB en pacientes con LES, hecho que tal vez explica la reducción de la subpoblación de memoria en el grupo de pacientes con LES y con OEA que se analizaron en nuestro estudio.
En cuanto a la expresión de CD1d, se observó que entre las subpoblaciones de células B evaluadas, los LB transicionales CD24hiCD38hi tenían una expresión más alta de esta molécula, lo que concuerda con resultados previos12.
En un estudio realizado en 2002 por Mizoguchi et al., se pudo evidenciar, en un modelo de colitis autoinmune experimental, que la regulación positiva de CD1d confería a las células B la capacidad de suprimir la progresión de la inflamación intestinal crónica, en ratones enfermos, por medio de un aumento en la producción de IL-103. Entre los LB con capacidad reguladora, los más ampliamente estudiados son los B10, cuyo fenotipo es CD19+CD5+CD1dhi y han sido descritos como potentes productores de IL-10, con capacidad de regular diferentes respuestas inmunes en diferentes contextos patológicos, principalmente enfermedades autoinmunes como AR, esclerosis múltiple, LES y colitis. Esta regulación es mediada principalmente por la IL-10 y depende de la presencia de múltiples moléculas como CD19, CD1d, CD40, CD80, CD86, MHC-II e IL-21R, induciendo una reducción en la frecuencia de LT de los perfiles pro-inflamatorios Th1 y Th17 y promoviendo la diferenciación de células T reguladoras6–8,18,29.
Por esta razón, en este estudio se quiso evaluar la expresión de CD1d, como una explicación parcial de las alteraciones funcionales previamente reportadas en las células Breg CD24hiCD38hi de pacientes con LES. Sin embargo, nuestros resultados no demostraron diferencias ni en el porcentaje de células B maduras, de memoria o transicionales CD1d+, ni en la expresión de CD1d en las subpoblaciones de células B evaluadas entre pacientes y controles.
Es de notar que durante la elaboración de este manuscrito, Bosma et al., reportaron una disminución en la expresión de CD1d en células B totales de pacientes con LES comparados con controles sanos, además de defectos en su reciclaje. Sin embargo, en este estudio no se evaluó la expresión de CD1d en subpoblaciones de LB20.
CD1d se encarga de presentar lípidos a células iNKT (por su sigla en inglés “Invariant Natural Killer T cells”), para las cuales se han descrito funciones reguladoras como la inhibición de producción de IL-17 e inducción del reclutamiento de células Treg30. Al respecto se ha observado que los pacientes con LES presentan una reducción numérica de las células iNKT y que además son refractarias a la estimulación, lo que probablemente se explique por la deficiencia observada en la expresión de CD1d en células B, ya que se ha demostrado que estas células son necesarias para el mantenimiento del número y la función de las iNKT20. Además, en este mismo estudio Bosma et al., pudieron observar una disminución en la expresión de CD1d en la subpoblación de LB inmaduros CD24hiCD38hi.
Dado que las células B CD24hiCD38hi son las que mayor expresión de CD1d presentan, es probable que esta subpoblación tenga un importante papel en la regulación ejercida sobre las células iNKT. Por esta razón la evaluación de la expresión de CD1d en esta subpoblación podría ayudar a entender mejor las alteraciones que se presentan en las células B reguladoras y en general en los mecanismos de tolerancia en el LES. Si bien en el presente estudio no se observaron diferencias en la expresión de CD1d en las células reguladoras CD24hiCD38hi, entre pacientes y controles, esto puede explicarse, al menos parcialmente, por el tamaño muestral, ya que en el estudio de Bosma se incluyó una cohorte de 254 pacientes, por lo cual el aumentar el número de pacientes y controles se presenta como una perspectiva en nuestra investigación. Además, el estudio de otros mecanismos reguladores como el ejercido por CD1d en las células B reguladoras, ayudará a entender la inmunopatología y a mejorar las alternativas terapéuticas no sólo en el lupus sino también en otras enfermedades en las que están implicadas estas células.
ConclusiónFinalmente, quedan por resolver múltiples aspectos de la biología de las células Breg en el humano y el modelo murino que permitan un mejor entendimiento de los mecanismos regulatorios de estas células, así como la comprensión del papel que cumplen estas células en la fisiopatología de enfermedades autoinmunes y su implicación en la etiología de enfermedades inflamatorias crónicas.
FinanciaciónComité para el desarrollo de la Investigación (CODI) Uni versidad de Antioquia, proyecto E001600, menor cuantía. Programa Joven Investigador Universidad de Antioquia. Programa Sostenibilidad 2013–2014 Universidad de Antioquia.
Conflicto de interesesLos autores declaran no tener ningún conflicto de intereses.
Grupo de Reumatología Universidad de Antioquia. Unidad de Citometría, Sede de Investigación Universitaria (SIU). Universidad de Antioquia. Hospital Universitario San Vicente Fundación. Pacientes y controles.
