


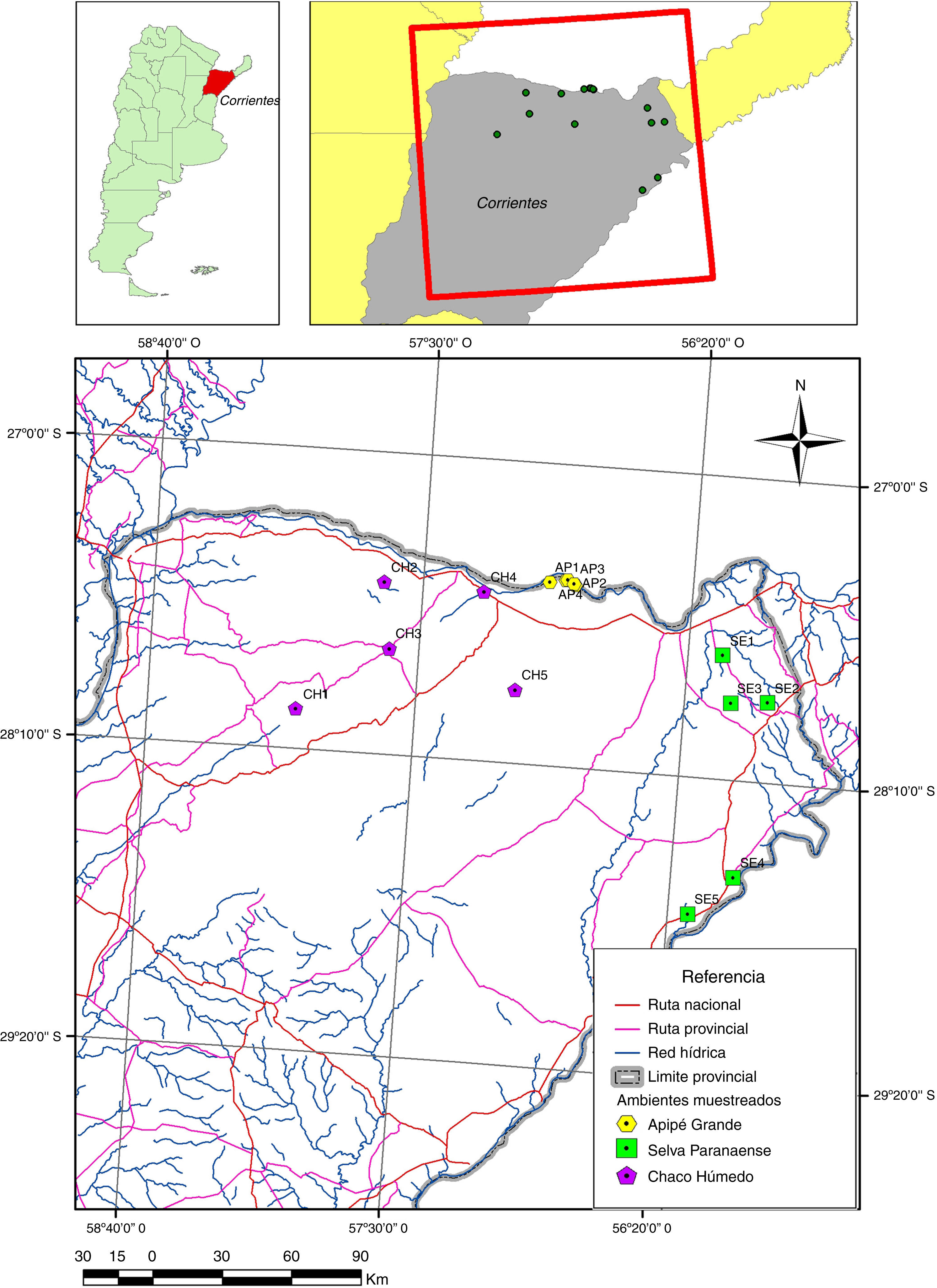


La isla Apipé Grande se encuentra al norte de la provincia de Corrientes, Argentina, siendo el Chaco Húmedo y la Selva Paranaense las regiones fitogeográficas adyacentes a ella. Aquí se determina la composición del ensamble de artrópodos de parches de bosques de Apipé Grande, se evalúa si presenta diferencias con comunidades de artrópodos de ambientes continentales similares, y se determina con cuál región fitogeográfica adyacente comparte más especies. Se muestrearon 5 sitios en cada una de las siguientes áreas: Apipé, Chaco Húmedo y Selva Paranaense, con un G-Vac sobre la vegetación. Se generaron curvas de rango-abundancia para comparar la composición de las comunidades, y con un NMDS se analizó el grado de similitud de los ensambles de especies/morfoespecies entre sitios. Se calculó el índice de Raup-Crick estandarizado, a fin de determinar si las comunidades se encuentran estructuradas al azar o no. Apipé presentó un ensamble propio, compartiendo un porcentaje de fauna con las comunidades continentales. Si bien muchos artrópodos compartidos entre el continente y Apipé no tienen altas capacidades de dispersión, esta limitación en la dispersión parece no ser un condicionamiento para estructurar los ensambles, siendo los requerimientos ambientales de las especies importantes para determinar la composición de las comunidades.
Apipé Grande Island is localized north of the province of Corrientes (Argentina), with the phytogeographic regions of the Chaco Húmedo and the Selva Paranaense adjacent to it. The objectives of the study were to determine the composition and arthropod assemblages in forest environments of Apipé Grande Island, and evaluate if they differ of continental arthropod communities of the same environment, and determine with which of those adjacent phytogeographic region (Chaco Húmedo or Selva Paranaense) shares more arthropod species. Five sites were sampled in each of the next studied areas: Apipé, Chaco Húmedo and Selva Paranaense, with a G-Vac over vegetation. Rank-abundance curves were generated to compare communities; and with a NMDS we analyzed the degree of assemblages association of species/morphospecies between sites. Raup-Crick standardized index was calculated to determine whether ecological communities have a nonrandom structure. Many arthropods species did not have high dispersal capabilities, but they were shared between the continental and Apipé communities, which suggests that the dispersal limitation of these species is not a constraint in the process of structuring assemblies. Environmental requirements of the species seem to be an important factor in determining the composition of these communities.
Las islas han sido frecuentemente citadas como un escenario ideal para estudiar procesos evolutivos porque en ellas existen altos niveles de endemismos y son ambientes que favorecen la formación de especies (Carson y Templeton, 1984; Grant y Grant, 1996; Losos, Jackman, Larson, de Queiroz y Rodrigues-Schettino, 1998), asimismo, han sido de gran interés para el estudio de procesos ecológicos (Pozo y Llorente-Bousquets, 2002). La principal característica de una isla es su aislamiento, no obstante, la fauna de los ambientes adyacentes contribuye al grupo de especies disponibles para colonizarla (Cowie y Holland, 2006; Gillespie y Roderick, 2002; Ricklefs, 2001). Por tal motivo, las zonas aledañas a la isla representan la fuente de especies potencialmente inmigrantes. Las islas difieren en tamaño, en distancia al continente más cercano u otras fuentes de colonización y en la heterogeneidad del hábitat, y son excelentes modelos para estudiar los efectos de estos rasgos sobre la diversidad y el recambio de especies (Rohde, 2005).
La isla Apipé Grande es una isla continental que se encuentra al norte de la provincia de Corrientes, frente al departamento Ituzaingó, a 2km río abajo de la represa hidroeléctrica de Yacyretá y separada de las costas continentales por no más de 10km en todo su perímetro. Integra el sistema de reservas de la provincia en el que se incluyen Rincón Santa María, la Unidad de Conservación Galarza y la Reserva Natural Iberá (Colonia Pellegrini), las 2 últimas en el Sistema de los Esteros del Iberá. La isla Apipé Grande fue declarada reserva en 1994, como parte del plan de mitigación de las obras del complejo hidroeléctrico de Yacyretá, encontrándose dentro del área de influencia de la represa (Fontana, 2008). Es la única isla argentina del alto Paraná con asentamientos urbanos permanentes y un 40% de su superficie se ve afectada directamente por las inundaciones periódicas, ocasionadas habitualmente por la apertura de las compuertas de la represa. La diversidad de ambientes y de tipos de vegetación hacen de la isla un excelente sitio para la conservación.
Si bien Cabrera (1976) sostiene que Apipé Grande pertenece a la provincia fitogeográfica Paranaense, a partir de diversos estudios sobre la vegetación, Carnevali (1994), Eskuche y Fontana (1996a, 1996b) y Hansen (1998) coinciden en que la isla se ubica en una zona de transición entre las provincias fitogeográficas Paranaense y Chaqueña Oriental. Por otro lado, Matteucci (2012) considera Apipé Grande dentro del complejo Lagunas y Esteros del Noroeste en los Esteros del Iberá. De esta manera, existen diferentes posturas y criterios sobre a qué provincia fitogeográfica pertenece esta isla.
En este trabajo se analiza si la fauna de artrópodos de los bosques de la isla Apipé Grande, por ser una zona de transición entre las provincias fitogeográficas Paranaense y Chaqueña Oriental, presenta elementos faunísticos de ambas regiones. El objetivo del presente trabajo es determinar la composición de los ensambles de artrópodos de bosques de la isla Apipé Grande y compararlos con los de bosques continentales, Chaco Húmedo y selva Paranaense, para conocer el aporte de cada uno de ellos sobre la fauna de la isla.
Materiales y métodosEl área de estudio comprendió la isla Apipé Grande (Corrientes, Argentina) y 2 regiones fitogeográficas aledañas, el Chaco Húmedo y la selva Paranaense (27°30’-28°50’ S y 55°30’-58°10’ O). La isla tiene una extensión de 30,000ha y se caracteriza por presentar una importante superficie de su territorio, aproximadamente el 70%, compuesta por zonas bajas inundables integradas por madrejones, esteros y lagunas (Hansen, 1998). En la zona central de la isla, siguiendo una dirección noroeste-sudeste, se presenta una zona baja donde se destacan las lagunas Hermosa y Cambá Cué, con unas áreas de aproximadamente 300 y 1,000ha, respectivamente. El Chaco Húmedo ocupa, en la provincia de Corrientes, la zona noroeste, posee un gran número de humedales distribuidos por toda la región, cubriendo más del 80% de dicha área (Brown, Martínez-Ortiz, Acerbi y Corcuera, 2006). La selva Paranaense se extiende sobre el extremo noreste de Corrientes y posee los bloques boscosos remanentes más grandes y de más alto valor en biodiversidad, junto con los Yungas (Placci y di Bitetti, 2006).
Se tomaron muestras de artrópodos en 5 sitios de bosque en cada uno de los ambientes estudiados: Apipé Grande, Chaco Húmedo y selva Paranaense (fig. 1), en primavera y otoño de los años 2006 y 2007, respectivamente. En cada sitio se tomaron 10 muestras al azar con un aspirador (G-Vac) Stihl® sobre la vegetación (Bell, Wheater, Henderson y Cullen, 2000; Borges y Brown, 2003; Dogramaci, DeBanco, Kimoto y Wooster, 2011; Smith, 1999). El material correspondiente a cada muestra fue colocado en bolsas de polietileno con alcohol etílico al 70% y se trasladaron al laboratorio para su procesamiento.
, Apipé Grande (AP) y selva Paranaense (SE).")
En el laboratorio se separó el material de artrópodos, el cual fue identificado en una primera instancia a nivel de especies/morfoespecies a través de una base de datos de fotografías digitales de los caracteres distintivos de cada una de ellas (TAXIS versión 3.5) (Meyke, 1999-2004). Luego, con la ayuda de claves taxonómicas disponibles (Borror, Triplehorn y Johnson, 1989; Buzzi y Miyazaki, 1999; De Santis, 1969; Dippenaar-Schoeman y Joqué, 1997; Elzinga, 2000; Ramírez, 1999), cada especie/morfoespecies fue identificada a nivel de orden y familia. En el análisis de datos se generaron curvas de distribución de rango-abundancia a fin de evaluar y comparar la estructura de dominancia de las comunidades de artrópodos recolectados en los 3 ambientes (Larrivée y Buddle, 2009; Magurran, 2004). Para analizar el grado de similitud de la fauna entre sitios se realizó un análisis de escalamiento multidimensional no métrico (NMDS, por sus siglas en inglés), utilizando como distancia el índice de Sorensen cuantitativo (Bray-Curtis). La validez del ordenamiento obtenido se determinó a partir de los valores de estrés y el valor de p, siguiendo la regla de Clarke (1993), que considera un estrés de 5-10 como un buen ordenamiento, sin riesgo real de inferencia falsa. Para el NMDS se calcularon primero los estadísticos descriptivos de la matriz principal, la que luego fue relativizada de acuerdo con la distancia de Hellinger. Esta es una medida recomendada para el ordenamiento de datos de abundancia de especies (Rao, 1995), y es una alternativa apropiada, ya que da poco peso a las especies raras (Legendre y Gallagher, 2001). Adicionalmente, la significación estadística de los grupos obtenidos en cada nivel fue evaluada a través de procedimientos de permutación de respuestas múltiples, empleando como medida de distancia el índice de Sorensen (Bray-Curtis). Estos análisis se realizaron con el programa PC-ORD versión 6 (McCune y Mefford, 2011), siguiendo la rutina propuesta por Peck (2010). Para determinar el porcentaje de fauna compartida entre los agrupamientos obtenidos en el NMDS se utilizó el índice de disimilitud, o recambio de especies, de Morisita para 3 comunidades (Chao, Jost, Chiang, Jiang y Chazdon, 2008), calculado por medio del programa SPADE (Chao y Shen, 2003-2005). Se calculó este índice, ya que los de similitud por pares no caracterizan completamente múltiples comunidades, debido a que la información que es compartida por las 3 comunidades es ignorada (Chao et al., 2008). Siguiendo la metodología propuesta por Chase, Kraft, Smith, Vallend e Inouye (2011), se calculó el índice de similitud de Raup-Crick que provee información sobre el grado en que las comunidades son más diferentes o similares que lo esperado por azar (Raup y Crick, 1979), utilizando el software PAST versión 2.14 (Hammer, Harper y Ryan, 2001). Los valores obtenidos se estandarizaron, de acuerdo con la forma propuesta por Chase et al. (2011), en valores de −1 a +1. De esta manera, para valores de 0 (cero) la disimilitud es igual a la esperada por azar; para valores positivos y negativos, la disimilitud es mayor o menor de la esperada por azar, respectivamente.
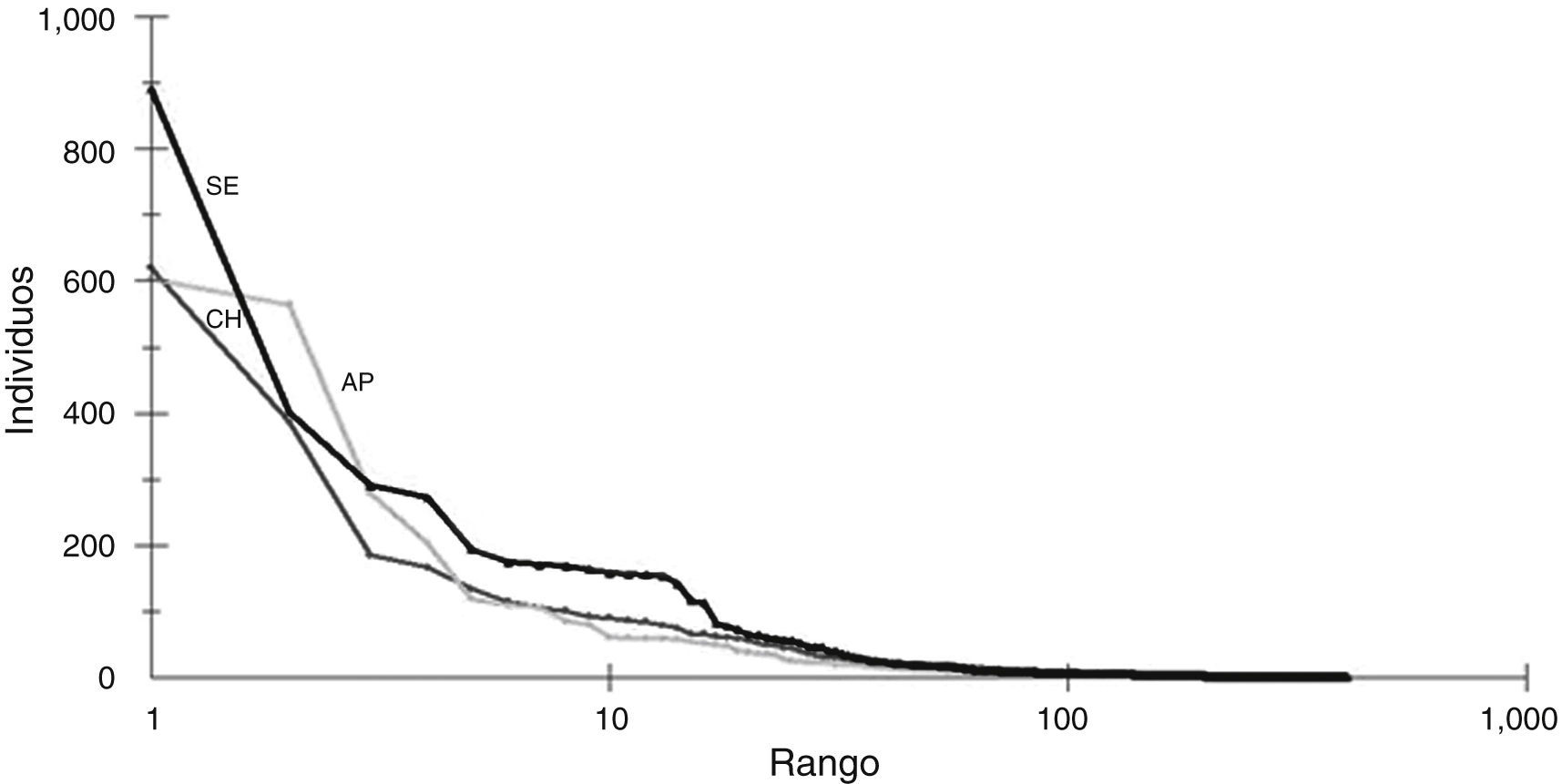
ResultadosLa fauna de Apipé Grande estuvo representada por un número de especies y una abundancia de artrópodos menor que los ambientes continentales (tabla 1). Al mismo tiempo, la comunidad de artrópodos de Apipé Grande tuvo más especies dominantes que los otros 2 ambientes, varias especies con abundancias intermedias y un menor número de especies raras. Por otra parte, el Chaco Húmedo y la selva Paranaense mostraron un menor número de especies dominantes, numerosas especies con abundancias intermedias y muchas especies raras (fig. 2).
Riqueza, abundancia y número de familias de los órdenes de artrópodos presentes en los ambientes de Chaco Húmedo, Apipé Grande y selva Paranaense en la provincia de Corrientes, Argentina
| CH | AP | SE | |||||||
|---|---|---|---|---|---|---|---|---|---|
| S | N | Fam | S | N | Fam | S | N | Fam | |
| Diptera | 122 | 2,148 | 21 | 111 | 2,556 | 21 | 142 | 3,269 | 25 |
| Araneae | 146 | 1,579 | 24 | 114 | 836 | 23 | 131 | 1,152 | 20 |
| Hymenoptera | 151 | 772 | 22 | 151 | 681 | 22 | 155 | 868 | 21 |
| Collembola | 10 | 320 | 2 | 7 | 146 | 2 | 8 | 1,217 | 2 |
| Acari | 21 | 493 | - | 21 | 311 | - | 19 | 790 | - |
| Coleoptera | 102 | 415 | 22 | 63 | 189 | 18 | 105 | 281 | 22 |
| Hemiptera | 90 | 243 | 20 | 87 | 322 | 20 | 96 | 243 | 21 |
| Trichoptera | 7 | 29 | - | 11 | 95 | - | 6 | 72 | - |
| Thysanoptera | 13 | 53 | - | 3 | 17 | - | 12 | 30 | - |
| Psocoptera | 7 | 11 | - | 11 | 25 | - | 4 | 12 | - |
| Neuroptera | 3 | 4 | 3 | 5 | 23 | 5 | 5 | 13 | 5 |
| Dictyoptera | 3 | 9 | - | 6 | 28 | - | 1 | 3 | - |
| Ephemeroptera | 3 | 15 | - | 3 | 5 | - | 2 | 8 | - |
| Lepidoptera | 3 | 8 | - | 3 | 11 | - | 2 | 6 | - |
| Pseudoescorpionida | 1 | 17 | - | 1 | 5 | - | 1 | 1 | - |
| Orthoptera | 4 | 4 | 4 | 3 | 3 | 1 | 3 | 3 | 2 |
| Isopoda | 0 | 0 | - | 0 | 0 | - | 1 | 2 | - |
| Odonata | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 |
| Embioptera | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 |
| Dermaptera | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mecoptera | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 689 | 6,123 | 601 | 5,254 | 694 | 7,971 | |||
AP: Apipé Grande; CH: Chaco Húmedo; SE: selva Paranaense; Fam: familias; N: abundancia; S: riqueza.
, Chaco Húmedo (CH) y selva Paranaense (SE) en la provincia de Corrientes, Argentina.")
En el ordenamiento resultante del NMDS se pudieron reconocer 3 grupos de sitios (fig. 3) correspondientes a cada uno de los ambientes estudiados. El ordenamiento en 2 ejes explicó poco más del 70% de la varianza (eje 1=0.401, eje 2=0.303), y con un estrés final de 9.50. El procedimiento de permutación de respuestas múltiples comprobó la existencia de los 3 grupos con una diferencia altamente significativa entre las comunidades de artrópodos analizadas (A=0.2094, p=0.00049). El primer eje del ordenamiento separó los sitios de los ambientes continentales de los de la isla, mientras que el segundo eje mostró una variación, sin revelar un patrón evidente, en la distribución de los sitios.
 y con un estrés=9.5052.AP: Apipé Grande; CH: Chaco Húmedo; SE: selva Paranaense.")
Ordenamiento por medio de un análisis de escalamiento multidimensional no métrico de los sitios muestreados en el área de estudio, mostrando diferencias en la fauna de artrópodos por ambiente (eje 1=0.401, eje 2=0.303) y con un estrés=9.5052.AP: Apipé Grande; CH: Chaco Húmedo; SE: selva Paranaense.
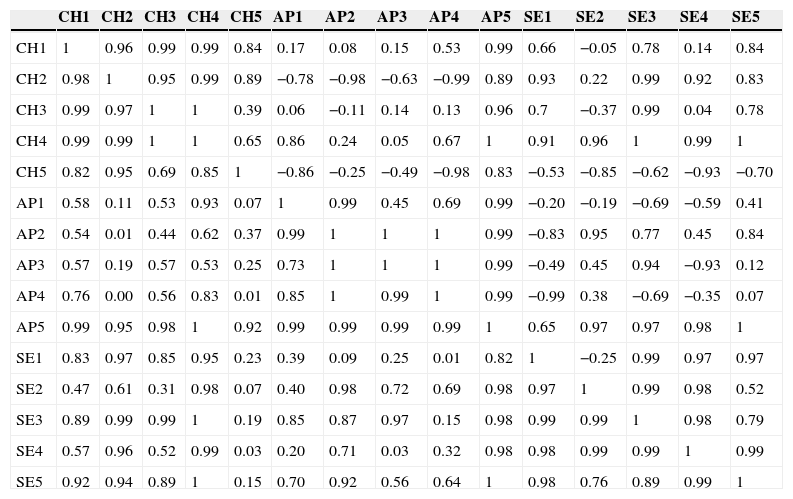
Los ensambles de artrópodos de los 3 ambientes estudiados compartieron muy pocas especies, tan solo 223. La comunidad de Apipé Grande resultó la más disímil, presentando una similitud con Chaco Húmedo del 36% (1-C22=0.639) y con selva Paranaense del 29% (1-C22=0.710). La tabla 2 muestra que las disimilitudes entre los sitios pueden explicarse en algunos casos por el azar, y en otros, no.
Similitud entre sitios de Chaco Húmedo, Apipé Grande y selva Paranaense en la provincia de Corrientes, Argentina
| CH1 | CH2 | CH3 | CH4 | CH5 | AP1 | AP2 | AP3 | AP4 | AP5 | SE1 | SE2 | SE3 | SE4 | SE5 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CH1 | 1 | 0.96 | 0.99 | 0.99 | 0.84 | 0.17 | 0.08 | 0.15 | 0.53 | 0.99 | 0.66 | −0.05 | 0.78 | 0.14 | 0.84 |
| CH2 | 0.98 | 1 | 0.95 | 0.99 | 0.89 | −0.78 | −0.98 | −0.63 | −0.99 | 0.89 | 0.93 | 0.22 | 0.99 | 0.92 | 0.83 |
| CH3 | 0.99 | 0.97 | 1 | 1 | 0.39 | 0.06 | −0.11 | 0.14 | 0.13 | 0.96 | 0.7 | −0.37 | 0.99 | 0.04 | 0.78 |
| CH4 | 0.99 | 0.99 | 1 | 1 | 0.65 | 0.86 | 0.24 | 0.05 | 0.67 | 1 | 0.91 | 0.96 | 1 | 0.99 | 1 |
| CH5 | 0.82 | 0.95 | 0.69 | 0.85 | 1 | −0.86 | −0.25 | −0.49 | −0.98 | 0.83 | −0.53 | −0.85 | −0.62 | −0.93 | −0.70 |
| AP1 | 0.58 | 0.11 | 0.53 | 0.93 | 0.07 | 1 | 0.99 | 0.45 | 0.69 | 0.99 | −0.20 | −0.19 | −0.69 | −0.59 | 0.41 |
| AP2 | 0.54 | 0.01 | 0.44 | 0.62 | 0.37 | 0.99 | 1 | 1 | 1 | 0.99 | −0.83 | 0.95 | 0.77 | 0.45 | 0.84 |
| AP3 | 0.57 | 0.19 | 0.57 | 0.53 | 0.25 | 0.73 | 1 | 1 | 1 | 0.99 | −0.49 | 0.45 | 0.94 | −0.93 | 0.12 |
| AP4 | 0.76 | 0.00 | 0.56 | 0.83 | 0.01 | 0.85 | 1 | 0.99 | 1 | 0.99 | −0.99 | 0.38 | −0.69 | −0.35 | 0.07 |
| AP5 | 0.99 | 0.95 | 0.98 | 1 | 0.92 | 0.99 | 0.99 | 0.99 | 0.99 | 1 | 0.65 | 0.97 | 0.97 | 0.98 | 1 |
| SE1 | 0.83 | 0.97 | 0.85 | 0.95 | 0.23 | 0.39 | 0.09 | 0.25 | 0.01 | 0.82 | 1 | −0.25 | 0.99 | 0.97 | 0.97 |
| SE2 | 0.47 | 0.61 | 0.31 | 0.98 | 0.07 | 0.40 | 0.98 | 0.72 | 0.69 | 0.98 | 0.97 | 1 | 0.99 | 0.98 | 0.52 |
| SE3 | 0.89 | 0.99 | 0.99 | 1 | 0.19 | 0.85 | 0.87 | 0.97 | 0.15 | 0.98 | 0.99 | 0.99 | 1 | 0.98 | 0.79 |
| SE4 | 0.57 | 0.96 | 0.52 | 0.99 | 0.03 | 0.20 | 0.71 | 0.03 | 0.32 | 0.98 | 0.98 | 0.99 | 0.99 | 1 | 0.99 |
| SE5 | 0.92 | 0.94 | 0.89 | 1 | 0.15 | 0.70 | 0.92 | 0.56 | 0.64 | 1 | 0.98 | 0.76 | 0.89 | 0.99 | 1 |
AP: Apipé Grande; CH: Chaco Húmedo; SE: selva Panaraense.
La parte inferior (blanco) contiene los valores originales de la medida de Raup-Crick, y la parte superior (gris) posee los valores modificados de la medida de Raup-Crick (βRC) de acuerdo con Chase et al. (2011). CH1 a 5: sitios 1 a 5 de Chaco Húmedo, respectivamente; AP1 a 5: sitios 1 a 5 de Apipé Grande, respectivamente, y SE1 a 5: sitios 1 a 5 de selva Paranaense, respectivamente.
BRC ∼0: Dis/Similitudobs∼Dis/Similitudesp (disimilitud igual a la esperada por azar); BRC∼1: Dis/Similitudobs>Dis/Similitudesp (las comunidades son más similares que lo esperado por azar); BRC∼−1: Dis/Similitudobs<Dis/Similitudesp (las comunidades son más disímiles que lo esperado por azar).
La riqueza de especies en una isla es el resultado de muchos procesos que afectan la colonización, la evolución y la persistencia de sus poblaciones (Ricklefs y Lovette, 1999). Así, en general, las biotas insulares se caracterizan por presentar una flora y una fauna depauperadas respecto a los ecosistemas continentales (Myers, 1990). Asociado a estos hechos, la reducida superficie que ocupa la isla hace que Apipé Grande presente una menor riqueza de artrópodos en comparación con el Chaco Húmedo y la selva Paranaense.
Además, la isla muestra un patrón en la estructura de sus comunidades de artrópodos diferente a los encontrados en los ambientes continentales, presentando un ensamble propio y particular que podría estar estructurado por diversos factores. Las alteraciones ambientales producto de las actividades humanas tienen un mayor impacto en las islas que en los ambientes continentales (Medel, Aizen y Zamora, 2009). Por su reducida superficie, posiblemente las perturbaciones antrópicas que imprimen los habitantes de la isla de Apipé Grande como quemas, deforestación, incremento de las zonas urbanizadas, jueguen un papel clave en la estructuración de las comunidades de artrópodos. A ello puede sumarse el efecto de las inundaciones periódicas ocasionadas por el derrame de las aguas del río Paraná sobre las barrancas costeras de la isla cuando se produce la apertura de las compuertas de la represa Yacyretá (Fontana, 2008), entre otras causas. Por otro lado, la endemicidad y los filtros impuestos a la inmigración de especies desde el continente (Whittaker y Fernández-Palacios, 2007) pueden también derivar en una biodiversidad particular en la isla, con muchas peculiaridades y diferencias con respecto a los ecosistemas continentales cercanos.
La existencia de un porcentaje de fauna compartido entre Chaco Húmedo, selva Paranaense y Apipé Grande es observada en los resultados del índice de similitud de Morisita para 3 comunidades; sin embargo, el conjunto de especies comunes a ellos es bajo (223 especies), en relación con el total de especies registradas en este estudio. Asimismo, el mayor porcentaje de artrópodos de la isla es compartido con el Chaco Húmedo, apoyando la idea de Matteucci (2012), quien considera Apipé Grande dentro del complejo de lagunas y esteros del noroeste en los Esteros del Iberá. Esto se contrapone a nuestra idea original, como lo habían propuesto varios autores, que la isla sería una zona de transición entre ambas regiones fitogeográficas (Carnevali, 1994; Eskuche y Fontana, 1996a, 1996b; Hansen, 1998). Esta mayor similitud observada entre el Chaco Húmedo y Apipé Grande puede estar relacionada con las características ambientales que comparten estos ambientes. Por un lado, ambos se caracterizan por un suelo permanentemente húmedo con amplias zonas inundables, y por otro, la gran variedad de ambientes presentes, como lagunas, esteros, madrejones, albardones, depresiones húmedas, dunas y lomadas (Fontana, 2008), que permitiría una mayor riqueza vegetacional y la presencia de un paisaje heterogéneo que es compartido por la isla y esta porción «continental».
Siguiendo la propuesta de Chase et al. (2011), quienes plantean la posibilidad de determinar si las comunidades están estructuradas o no al azar, pudimos identificar que no solo los filtros ambientales compartidos entre los sitios estarían estructurando nuestras comunidades. Otros mecanismos, tales como procesos no estocásticos, también están involucrados en la determinación de la composición de las comunidades de artrópodos en el área de estudio en la isla Apipé Grande. Por lo tanto, el aislamiento, la dispersión, las interacciones bióticas (competencia, depredación, etc.) y los requerimientos ambientales de las distintas especies (Borges y Brown, 2004; Borges et al., 2006; Chase et al., 2011; Jiménez-Valverde, Baselga, Melic y Tyasko, 2009) serían los eventos que están estructurando las comunidades analizadas. La dispersión, similitud observada igual a la esperada por azar, fue mayor entre Chaco Húmedo y Apipé Grande (tabla 2), donde los condicionamientos biológicos de las especies se encuentran centrados fundamentalmente en el poder de dispersión de los organismos existentes en el «pool continental» y en el poder de establecimiento de los que logran arribar a la isla. Así, únicamente los organismos con poder de dispersión a larga distancia van a ser capaces de atravesar la barrera de agua que separa los ambientes «continentales» de los de la isla (Fernández-Palacios, 2004).
Para futuros estudios sería interesante plantear nuevos interrogantes y muestreos a fin de verificar la existencia de un efecto insular entre el «continente» y Apipé Grande. Para esto sería conveniente también poder evaluar el aporte que hace a la comunidad de artrópodos de la isla la ecorregión Pastizales de la Mesopotamia, que se encuentra representada en la costa opuesta a la muestreada correspondiente a Paraguay. Se podría probar si la colonización, la evolución y el mantenimiento de las poblaciones en la isla difieren entre los taxones.
Los autores agradecen al Consejo Nacional de Investigaciones de la Universidad Nacional de Salta, al Consejo Nacional de Investigaciones Científicas y Técnicas y a la Universidad Nacional del Nordeste su apoyo y colaboración en el desarrollo de este proyecto.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.