




Se estudió a las moscas parásitas (Diptera: Streblidae) en una comunidad de murciélagos en la cueva de San Francisco, Chiapas. Los datos se obtuvieron entre febrero y agosto de 2013 mediante captura y revisión de 569 individuos de 12 especies de murciélagos. Se identificaron 3 especies de moscas: Trichobius joblingi, Metelasmus pseudopterus y Megistopoda aranea; T. joblingi se presentó en 6 especies de murciélagos mostrando los mayores valores de prevalencia y abundancia. En este trabajo se registra por primera vez en México la presencia simultánea de estas 3 especies como parásitos en Artibeus jamaicensis, además, la presencia de T. joblingi es un nuevo registro para A. jamaicensis y Pteronotus parnellii en Chiapas. La mayor prevalencia y abundancia de T. joblingi se presentó en hembras de Desmodus rotundus y en juveniles de A. jamaicensis, aunque no se encontró diferencia en la intensidad de infección, lo que demuestra que las características biológicas, como el sexo y la edad de los hospederos, pueden influir en la relación ectoparásito-hospedero. Se sugiere que la prevalencia, abundancia e intensidad de infección de ectoparásitos también podrían relacionarse con patrones de conducta social y hábitos de refugio.
Batflies (Diptera: Streblidae) were studied in a community of bats in the cave of San Francisco, Chiapas. Data were collected between February and August 2013 through capture and review of 569 individuals of 12 species of bats. Three bat flies species were identified: Trichobius joblingi, Metelasmus pseudopterus and Megistopoda aranea; T. joblingi occurred in 6 species of bats showing the highest values of prevalence and abundance. In this work, the co-occurrence of these 3 species in Artibeus jamaicensis reported for the first time in Mexico, and the presence of T. joblingi is a new record for A. jamaicensis and Pteronotus parnellii in Chiapas. The higher prevalence and abundance of T. joblingi occurred in females of Desmodus rotundus and juveniles of A. jamaicensis, although no differences in the intensity of infection were found, indicating that biological characteristics such as sex and age of the host may influence ectoparasite-host relationship. It is suggested that the prevalence, abundance and intensity of infection of external parasites may also be related to social behavior patterns and roosting habits.
El conocimiento de la relación parásito-hospedero se basa en la premisa de que el parásito afecta directa o indirectamente la homeostasis del hospedero (Combes, 2005). Se ha encontrado que los hospederos presentan una serie de atributos que los hacen más o menos propensos al parasitismo. Dentro de estos atributos existen muchos aspectos intrínsecos de las especies, como los hábitos alimentarios, la estructura social y los patrones de conducta, áreas de distribución, áreas de actividad, migración e historia evolutiva. Además, a nivel individual existen otros atributos que determinan el parasitismo como la edad, el tamaño corporal, el sexo, los aspectos genéticos individuales y el estado nutricional (Poulin y Morand, 2004).
Entre los ectoparásitos que presentan los murciélagos en América se han registrado a las familias de Nycteribiidae, Streblidae, Spinturnicidae, Ixodidae y Argasidae, principalmente (Beloto et al., 2005; Estrada-Peña y Sánchez-Acedo, 1998; Sheeler-Gordon y Owen, 1999). La familia Streblidae (Diptera) está conformada por moscas hematófagas (estréblidos) que ocupan las membranas de la piel y las alas de murciélagos (Dick, 2006; Dick y Patterson, 2006). Esta familia se encuentra en todas las regiones biogeográficas, principalmente en las zonas tropicales (Wenzel, 1976). Se han reconocido 5 subfamilias, 3 de ellas exclusivas del Nuevo Mundo: Trichobiinae, Strebliinae y Nycterophiliinae (Wenzel y Peterson, 1987). En el continente americano se ha registrado a la familia Streblidae como parásitos de murciélagos de las familias Emballonuridae, Noctilionidae, Molossidae, Natalidae, Furipteridae, Vespertilionidae y principalmente Phyllostomidae (Graciollii y Barros de Carvalho, 2001). Aunque existen estudios que han analizado la diversidad y riqueza de especies de ectoparásitos asociados a murciélagos, son pocos los que se han enfocado en analizar la relación parásito-hospedero (Muñoz, Aguilera y Casanueva, 2003; Patterson, Dick y Dittmar, 2007; Patterson, Dick y Dittmar, 2008; Presley y Willig, 2008).
En México se han realizado varios trabajos con ectoparásitos asociados a murciélagos, la mayoría de estos sobre taxonomía y descripción de especies (Guerrero y Morales-Malacara, 1996; Morales-Malacara, 2001; Morales-Malacara y Juste, 2002; Morales-Malacara y López, 1990; Morales-Malacara, Guzmán-Cornejo y López-Ortega, 2002). Algunos otros se han enfocado a documentar listados de especies y la asociación ectoparásito-hospedero (Hoffmann, 1990; Villegas-Guzmán, López-González y Vargas, 2005). Para Chiapas la información sobre este tema es escasa, existe solamente un estudio descriptivo de la diversidad de artrópodos asociada a una comunidad de murciélagos de la familia Phyllostomidae (Colín, 2006).
Sin embargo, los estudios enfocados en la relación parásito-hospedero son pocos. Es importante analizar dicha relación, puesto que se ha documentado que factores como la especie, sexo, edad, peso y condición reproductiva pueden influir en la magnitud del parasitismo (Bursten, Kimsey y Owing, 1997; Da Fonseca, 1957; Fernández, 1985; Linsay y Galloway, 1997; Loomis, 1956; Muñoz et al., 2003; Pérez, Granados, Soriguer y Ruiz-Martínez, 1996). El propósito de este estudio fue identificar las especies de moscas de la familia Streblidae asociadas a las diferentes especies de murciélagos en la cueva de San Francisco, La Trinitaria, Chiapas; se tomaron en cuenta la prevalencia, abundancia e intensidad de infección de las poblaciones componentes de ectoparásitos con el sexo y la edad de sus hospederos.
Materiales y métodosÁrea de estudioLa cueva de San Francisco se eligió para este estudio debido a la alta diversidad y abundancia de quirópteros que alberga. Esta cueva se localiza en las coordenadas 16°05’54.4” N, 92°02’45.6” S, a 2.5km al SE de la cabecera municipal de La Trinitaria, Chiapas, a una altitud de 1,542m snm (fig. 1). El tipo de vegetación alrededor de la cueva es bosque de encina, así como cultivos de maíz y pastizales. En las áreas adyacentes se localiza la cabecera municipal de La Trinitaria y una comunidad rural conocida como Pamala (fig. 1). El clima del área de estudio es semicálido, con lluvias en verano A(C) w”1. La temperatura media anual es de 19.9°C, los meses más calurosos son abril, mayo y junio, y los más fríos diciembre y enero. La precipitación promedio anual es de 1,030.5mm y la época lluviosa comprende de mayo a octubre (Álvarez-Castañeda y Álvarez, 1991; Chávez, 2008). La cueva de San Francisco se encuentra entre las 100 cuevas más grandes de México, con una profundidad de 1.7km. Presenta una entrada única de unos 30m de ancho por 10m de alto (Chávez, 2008).
Trabajo de campo
Los datos de campo se obtuvieron durante los meses de febrero a agosto del 2013 mediante la revisión de murciélagos capturados al salir de la cueva San Francisco (permiso SGPA/DGVS/01665/13). Se realizaron muestreos mensuales de 3 noches consecutivas, utilizando una red de niebla de 9m de longitud colocada a una distancia aproximada de 15m de la entrada a la cueva. El horario de operación de la red fue entre las 20:20h y las 02:00h. De esta forma, se aplicó un esfuerzo total de muestreo de 105horas-red. Los murciélagos capturados se midieron de la longitud de ambas orejas y antebrazos, y se pesaron para posteriormente ser identificados a nivel de especie con la ayuda de las guías de campo de Medellín, Arita y Sánchez (2008) y Reid (1997). Así también se obtuvieron los datos de clase de edad, sexo y estado reproductivo de cada ejemplar capturado, se aplicó un marcaje con tinta indeleble, con el fin de evitar contar varias veces a un mismo individuo.
Registro y recolección de ectoparásitosCada murciélago capturado se inspeccionó para localizar a los ectoparásitos. Una vez ubicados, los ectoparásitos se contaron y se extrajeron 3 ejemplares de cada morfoespecie con la ayuda de pinzas de disección y un pincel con alcohol. Las muestras de ectoparásitos fueron colocadas en un frasco con alcohol al 70% para su identificación en laboratorio, y se etiquetaron con los datos correspondientes como localidad, fecha, especie y número de registro individual del murciélago hospedero. Para cada murciélago se registró el número de ectoparásitos encontrados, así como el número y nombre de las morfoespecies presentes: abundancia y riqueza. Posteriormente los murciélagos fueron fotografiados y liberados cerca de la cueva.
Identificación de estréblidosPara identificar a cada ectoparásito se revisó cada individuo recolectado con la ayuda de un microscopio estereoscópico, y con claves especializadas (Guimarães y D’Andretta, 1956; Wenzel, 1976). Una vez identificados los ectoparásitos se separaron por especie, se registraron y organizaron los datos de riqueza, listado de especies y número de parásitos por individuo y por especie de murciélago hospedero. Todos los ejemplares estudiados (ejemplares voucher) están depositados en la colección entomológica de ECOSUR, San Cristóbal de Las Casas, Chiapas (ECO-SC-E).
Análisis de datosPara estudiar la población componente (definida por Bush, Lafferty, Lotz y Shosta, 1997 como todos los parásitos individuales que se encuentren en un estadio de desarrollo particular en un mismo lugar y tiempo en una especie de hospedero) de estréblidos dentro de la comunidad de murciélagos capturados, se utilizaron los parámetros de prevalencia, abundancia e intensidad de infección con relación a los atributos de cada población capturada de las especies de hospederos (Bush et al., 1997). La prevalencia es el porcentaje de hospederos infectados con uno o más individuos de una especie de ectoparásito en particular, dividido por el número de hospederos examinados. La prevalencia es una medida sencilla de presencia/ausencia de especies de ectoparásitos en una muestra de hospederos clasificados en 2 grupos: infectados y no infectados. La abundancia es el número de individuos de una especie de ectoparásito particular presente en una población de hospederos (Bush et al., 1997). La intensidad de la infección representa el número de individuos de una especie de ectoparásito particular que se encuentra presente en una sola especie de hospedero; para calcular este parámetro se considera únicamente la subpoblación de individuos infectados (Bush et al., 1997).
Para determinar si los atributos intrínsecos de los hospederos (sexo y edad) eran condicionantes de la prevalencia de ectoparásitos, se calculó la prevalencia para cada sexo (macho/hembra) y la categoría de edad (juveniles/adultos). Posteriormente, se compararon los valores de prevalencia por cada uno de los atributos mediante una prueba de Chi-cuadrado. Para determinar si el sexo (macho/hembra) y la edad (juveniles/adultos) del hospedero favorecían la abundancia y la intensidad de infección, se utilizó la prueba de U de Mann-Whitney. Todos los análisis estadísticos de infrapoblación de ectoparásitos se llevaron a cabo con el programa estadístico SPSS (SPSS, 1999).
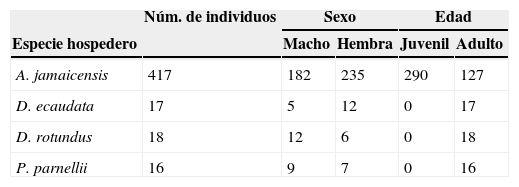
Debido a que el tamaño de muestra de algunas especies de hospederos fue demasiado pequeño, o no se contaba con suficientes individuos para dividirlos por grupo de sexo y edad, los análisis se realizaron solo para 4 de las 10 especies de murciélagos que presentaron estréblidos. De igual manera, para los hospederos Diphylla ecaudata, Desmodus rotundus y Pteronotus parnellii no fue posible realizar comparaciones entre clases de edad debido a que no se capturaron juveniles (tabla 1).
Número de individuos por sexo y clase de edad de murciélagos hospederos analizados en las pruebas de prevalencia, abundancia e intensidad de infección en la cueva de San Francisco, La Trinitaria, Chiapas, México.
| Núm. de individuos | Sexo | Edad | |||
|---|---|---|---|---|---|
| Especie hospedero | Macho | Hembra | Juvenil | Adulto | |
| A. jamaicensis | 417 | 182 | 235 | 290 | 127 |
| D. ecaudata | 17 | 5 | 12 | 0 | 17 |
| D. rotundus | 18 | 12 | 6 | 0 | 18 |
| P. parnellii | 16 | 9 | 7 | 0 | 16 |
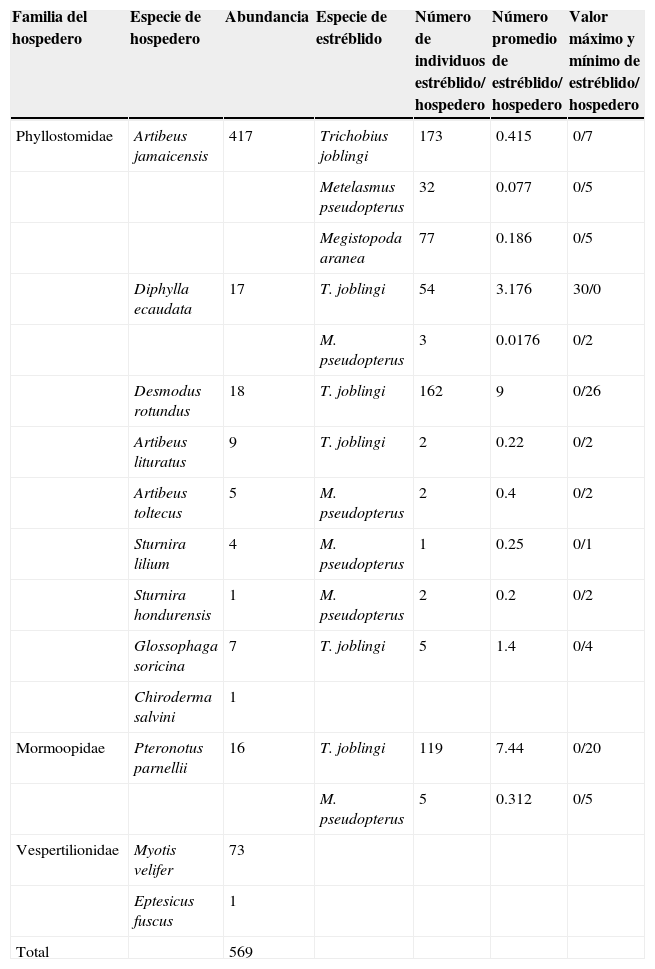
Se capturaron 569 individuos de 12 especies de murciélagos, de las cuales se registraron moscas estréblidas en 9 especies. El murciélago frugívoro Artibeus jamaicensis destacó por ser la especie de hospedero con mayor riqueza y abundancia de ectoparásitos, seguida por las 2 especies hematófagas, Diphylla ecaudata y Desmodus rotundus (tabla 2).
Lista de las especies de murciélagos capturados, su abundancia y presencia de estréblidos registrados por hospedero en la cueva de San Francisco, La Trinitaria, Chiapas, México.
| Familia del hospedero | Especie de hospedero | Abundancia | Especie de estréblido | Número de individuos estréblido/ hospedero | Número promedio de estréblido/ hospedero | Valor máximo y mínimo de estréblido/ hospedero |
|---|---|---|---|---|---|---|
| Phyllostomidae | Artibeus jamaicensis | 417 | Trichobius joblingi | 173 | 0.415 | 0/7 |
| Metelasmus pseudopterus | 32 | 0.077 | 0/5 | |||
| Megistopoda aranea | 77 | 0.186 | 0/5 | |||
| Diphylla ecaudata | 17 | T. joblingi | 54 | 3.176 | 30/0 | |
| M. pseudopterus | 3 | 0.0176 | 0/2 | |||
| Desmodus rotundus | 18 | T. joblingi | 162 | 9 | 0/26 | |
| Artibeus lituratus | 9 | T. joblingi | 2 | 0.22 | 0/2 | |
| Artibeus toltecus | 5 | M. pseudopterus | 2 | 0.4 | 0/2 | |
| Sturnira lilium | 4 | M. pseudopterus | 1 | 0.25 | 0/1 | |
| Sturnira hondurensis | 1 | M. pseudopterus | 2 | 0.2 | 0/2 | |
| Glossophaga soricina | 7 | T. joblingi | 5 | 1.4 | 0/4 | |
| Chiroderma salvini | 1 | |||||
| Mormoopidae | Pteronotus parnellii | 16 | T. joblingi | 119 | 7.44 | 0/20 |
| M. pseudopterus | 5 | 0.312 | 0/5 | |||
| Vespertilionidae | Myotis velifer | 73 | ||||
| Eptesicus fuscus | 1 | |||||
| Total | 569 |
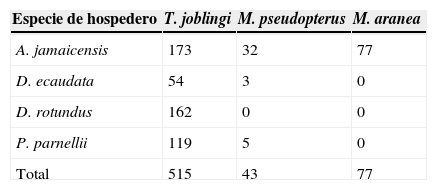
La mosca Trichobius joblingi tuvo mayor prevalencia dentro de la comunidad, estuvo presente en las 4 especies de hospederos analizadas. La mayor prevalencia se encontró en el hospedero Pteronotus parnellii, mientras que el valor más bajo correspondió a la población capturada de A. jamaicensis (fig. 2).

En el hospedero D. rotundus se encontraron diferencias significativas de la prevalencia de T. joblingi entre machos y hembras (Chi-cuadrado=4.500, n=18, gl=1 y p=0.034), los valores más altos de prevalencia correspondieron a las hembras. Por otra parte, para A. jamaicensis, D. ecaudata y P. parnellii los resultados mostraron que no hay diferencia significativa entre machos y hembras con relación a la prevalencia de los ectoparásitos registrados, por lo que se asumió que el sexo del hospedero no afecta a la prevalencia de ectoparásitos en estas especies de murciélagos.
Para A. jamaicensis se observó que la prevalencia de T. joblingi puede ser explicada por la edad, ya que se encontraron valores más altos de prevalencia en juveniles que en adultos (Chi-cuadrado=61.337, n=417, gl=1 y p=0.001).
AbundanciaEl estréblido T. joblingi fue la especie que presentó mayor abundancia en las 4 especies de hospederos analizadas, seguida de Megistopoda aranea, la cual solo se registró en un hospedero (A. jamaicensis). Finalmente Metelasmus pseudopterus fue la menos abundante, aunque cabe mencionar que esta se registró en 3 especies de hospederos (tabla 3).
En D. rotundus se encontró que el sexo sí influye en la abundancia de T. joblingi. En este caso se observaron diferencias significativas entre la abundancia en hembras y machos, las hembras presentaron mayor abundancia con relación a los machos (Mann-Whitney U=9.000, n=18 y p=0.009). Por su parte, no hubo diferencias en la abundancia de moscas entre machos y hembras de los hospederos A. jamaicensis, D. ecaudata y P. parnellii.
Para la población capturada de A. jamaicensis, la prueba de Mann-Whitney sugiere que T. joblingi (U=12,282.000, n=417, p=0.001) y M. pseudopterus (U=17,405.500, n=417, p=0.038) son significativamente más abundantes en juveniles que en adultos, mientras que para M. aranea los resultados no mostraron diferencias significativas (U=17,319, n=417, p=0.155).
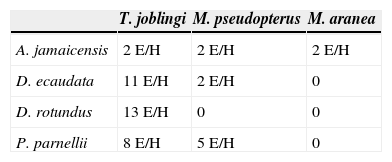
Intensidad de infecciónSe encontró que 3 especies de hospederos (D. ecaudata, D. rotundus y P. parnellii) presentaron la mayor intensidad de infección del ectoparásito T. joblingi. De acuerdo con los resultados obtenidos se encontró que la intensidad de infección dentro de la muestra de murciélagos capturados no mostró diferencia entre los atributos sexo y edad de cada especie de hospedero (tabla 4).
DiscusiónEn este estudio se revisaron 569 individuos pertenecientes a 12 especies de murciélagos. Se registró una especie de ectoparásito (T. joblingi) no documentada para A. jamaicensis y P. parnellii en Chiapas (Colín, 2006; Guerrero y Morales-Malacara, 1996). Sin embargo, sí existen registros de T. joblingi para la especie A. jamaicensis (González, Santos y Miranda, 2004; Ortega y Castro-Arellano, 2001). Aunque González et al. (2004) registran que este ectoparásito presenta bajas abundancias en A. jamaicensis, a diferencia a lo registrado en este estudio. Se considera que la ausencia de T. joblingi en el hospedero A. jamaicensis en los trabajos de Colín (2006) y Guerrero y Morales-Malacara (1996) puede deberse al tamaño de la muestra de los hospederos revisados, ya que para el primer estudio se revisaron 106 individuos de diversas especies de murciélagos, sin embargo, no se indica cuántos de estos individuos correspondieron a la especie de A. jamaicensis, ni a P. parnellii. En el caso del trabajo realizado por Colín (2006) indica que la muestra fue de 12 ejemplares de A. jamaicensis, mientras que nosotros obtuvimos un total de 417 ejemplares de esta especie. Por su parte, el ectoparásito M. aranea se registró únicamente en A. jamaicensis, lo cual concuerda con lo documentado por Overal (1980), quien señala que este parásito es más específico del hospedero A. jamaicensis, aunque la prevalencia que él registra (60%) difiere en gran medida de lo documentado en este estudio (11.4%).
Dentro de la comunidad de murciélagos cavernícolas capturados en este estudio, el ectoparásito T. joblingi resultó ser la especie más prevalente y la más abundante en 4 especies de hospederos. De acuerdo con los resultados obtenidos en el presente trabajo, y otros estudios en donde se registra T. joblingi en especies como Carollia perspicillata y Sturnira hondurensis (Guerrero y Morales-Malacara, 1996), se infiere que esta especie es generalista.
Se ha documentado que el tamaño de la población de los hospederos afecta de manera directa el aumento de la prevalencia y la intensidad de infestación de moscas de murciélagos (Dick y Patterson, 2006; Ter Hofstede y Fenton, 2005); es decir, en poblaciones abundantes de murciélagos se puede esperar mayor prevalencia de los parásitos. Así, se esperaría que A. jamaicensis y Myotis velifer que tuvieron mayor número de capturas deberían ser las especies con las tasas más altas de prevalencia y abundancia. Sin embargo, en este estudio el valor más alto de prevalencia del estréblido T. joblingi correspondió al hospedero P. parnellii, mientras que el más alto de intensidad de infección correspondió a D. rotundus, ambos hospederos con bajas tasas de captura con relación a A. jamaicensis. Diversos estudios realizados en Latinoamérica sobre la familia Streblidae documentan que la estructura social, la selección del refugio y el comportamiento de aseo de los murciélagos afectan la densidad de estréblidos (Dick y Patterson, 2006; Ter Hofstede y Fenton, 2005). Con frecuencia ocurre que en una comunidad con una alta densidad de murciélagos se presenta una alta densidad y riqueza de ectoparásitos (Dick y Gettinger, 2005; Dick y Patterson, 2006; Ter Hofstede y Fenton, 2005; Wenzel, 1976; Wenzel, Tipton y Kiewlicz, 1966).
Cabe destacar que en este estudio no todas las especies de murciélagos encontradas se registraron a lo largo de todo el muestreo. Algunas solo se presentaron en ciertos meses y otras exclusivamente en una o 2 ocasiones. Esto puede deberse a que no todas las especies usan las cuevas con la misma frecuencia, ya que para algunos murciélagos las cuevas son su principal refugio, mientras que otros las utilizan como sitios alternativos, y también existen otros que las pueden llegar a ocupar de forma esporádica (Arita, 1993). En este estudio las especies que se capturaron de manera constante a lo largo del trabajo de campo fueron: P. parnellii, D. rotundus, D. ecaudata y A. jamaicensis, a excepción de esta última, son quirópteros que utilizan las cuevas como su principal refugio (Arita, 1993). Así, se infiere que los hospederos A. jamaicensis, D. ecaudata, D. rotundus y P. parnellii utilizan la cueva San Francisco como refugio permanente, a diferencia de las otras especies registradas en la cueva, las cuales tuvieron poca o nula riqueza y abundancia de estréblidos.
Con relación a la especie M. velifer, de la cual se obtuvo una muestra suficiente (77 individuos) no se registraron estréblidos. Esta especie solo fue capturada durante los meses de junio y julio. Algunos autores señalan que dicha especie suele hibernar en Estados Unidos de América (Fitch, Shump y Shump, 1981). Para México no se ha registrado que M. velifer tenga este hábito. Sin embargo, dadas las fechas de captura se infiere que este hospedero usa la cueva como refugio alternativo durante la época de lluvias. Este factor pudiera responder el por qué en esta especie no se encontraron estréblidos, ya que se han documentado al menos 24 especies de ectoparásitos, de las cuales 2 pertenecen a la familia Streblidae (Jameson, 1959; Reisen, Kennedy y Reisen, 1976; Ubelaker, 1966). No obstante, cabe señalar que dichas especies de estréblidos no corresponden a las especies registradas en este estudio.
Prevalencia, abundancia e intensidad de infección con relación al sexo y edad del hospederoDebido a que los trabajos realizados sobre la influencia del sexo y edad de los murciélagos en la prevalencia, abundancia e intensidad de infección por parásitos son limitados, y que muchas veces la información existente es sobre especies de hospederos y ectoparásitos distintas a las encontradas en el presente estudio, es difícil realizar una comparación. Sin embargo, retomando trabajos realizados fuera de México con murciélagos como grupo de hospederos de una gran variedad de ectoparásitos se ha encontrado que la prevalencia de estos en murciélagos es muy variable (de 0% a 100%), según la especie de murciélago y de ectoparásito (Imaz, Aihartza y Totorika, 1999; Marinkelle y Grose, 1981). En el presente trabajo la prevalencia máxima fue de 87.5% y correspondió al hospedero P. parnellii por parte del estréblido T. joblingi, y el porcentaje más bajo de prevalencia correspondió al estréblido M. pseudopterus registrado en A. jamaicensis. En cuanto a la intensidad de infección los valores son muy diversos y dependientes de la especie de ectoparásito, como lo han señalado también otros autores (Durden, Best, Wilson y Hilton, 1992; Loomis, 1956; Marinkelle y Grose, 1981 y Reisen et al., 1976). En este trabajo la intensidad de infección fue baja (el valor máximo obtenido fue de 13 ectoparásitos por hospedero) con relación a trabajos realizados en otras especies de murciélagos en donde se han recolectado entre 90 y 112 individuos de ectoparásitos por individuo de hospedero (Reisen et al., 1976; Whitaker, Duff y Bellwood, 2000).
Los resultados obtenidos sugieren que la clase de edad y el sexo pueden influir sobre la prevalencia y abundancia de moscas parásitas en algunas especies de murciélagos. Por una parte, se encontró que en el hospedero D. rotundus las hembras presentaron mayor prevalencia y abundancia de T. joblingi, lo cual concuerda con lo documentado por Patterson et al. (2008), quienes encontraron que las hembras son más parasitadas. Los resultados obtenidos en este estudio también confirman las conclusiones principales de Presley y Willig (2008), quienes indican que las hembras son generalmente más fuertemente atacadas por ectoparásitos. Este fenómeno puede deberse a la diferencia en la selección de refugio entre machos y hembras (Patterson et al., 2008; Zahn y Rupp, 2004), ya que ellas generalmente seleccionan refugios de mejor calidad que atraen a un mayor número de estréblidos, lo que a su vez facilita mayores niveles de infestación, lo que las hace más vulnerables (Krasnov, Shenbrot y Khokhlova, 2002; Patterson et al., 2008; Zahn y Rupp, 2004). Esto se relaciona con la ecología y la conducta social y reproductiva, ya que las hembras de muchas especies de murciélagos forman colonias de maternidad densas (McCracken, 1984; Patterson et al., 2008). Al estar perchados en un mismo refugio, ya sea refugio de maternidad, de reproducción o de hibernación, los individuos están más cercanos unos a otros, lo que facilita el movimiento de estas moscas entre murciélagos (Overal, 1980). Por otra parte, las hembras de muchas especies de mamíferos disfrutan de tasas de sobrevivencia mayor, lo que podría explicar porqué los ectoparásitos tienen mayor afinidad por las hembras.
En otro estudio realizado con la especie Tadarida brasiliensis (Muñoz et al., 2003) se encontró que las hembras presentaron mayor prevalencia de ectoparásitos que los machos. Este resultado se atribuye al comportamiento del hospedero con relación a la mayor o menor actividad territorial y contacto con otros individuos de la especie (Bursten et al., 1997; Fernández, 1985; Muñoz et al., 2003), así como las diferencias químicas, como de la piel, hormonas circulantes, entre otros; o morfológicas del hospedero que hacen que un sexo sea mejor fuente de alimento que el otro (Bursten et al., 1997; Muñoz et al., 2003).
Los resultados obtenidos indican que los hospederos juveniles de A. jamaicensis tienen mayor prevalencia y abundancia de estréblidos. Beloto et al. (2005) señalan que la etapa de desarrollo con la mayor intensidad de infección es la juvenil, posiblemente debido a que la conducta de acicalamiento aún no está muy marcada. Las moscas también pueden parasitar a los juveniles como parte de una estrategia de dispersión, ya que estos buscan nuevos refugios, y por ende, nuevos sitios potenciales para los estréblidos. Otra explicación se enfoca en la estrecha relación de las hembras con sus crías, por lo que las primeras pueden transmitir las moscas estréblidas a sus descendientes.
Relación parásito, hospedero y refugioDurante el estudio se observó que las especies de murciélagos que presentaron mayor riqueza y abundancia de estréblidos (A. jamaicensis, P. parnellii, D. ecaudata y D. rotundus) comparten la característica de seleccionar refugios húmedos como cavernas, especialmente aquellas que contienen alguna fuente de agua, además de que estas 4 especies tienden a formar colonias numerosas (Lord, 1988; Ortega y Arita, 2004). Varios estudios sugieren que los hábitos de refugio pueden impactar significativamente los niveles de parasitismo (Bordes, Monrad y Ricardo, 2008; Patterson et al., 2007; Ter Hofstede y Fenton, 2005). Los hábitos de percha favorecen significativamente la carga parasitaria cuando hay una alta fidelidad a refugios cerrados, lo que da como resultado una alta prevalencia, abundancia y riqueza de especies de ectoparásitos en los hospederos (Patterson et al., 2007). Las cuevas en particular favorecen esta interacción de parásito-hospedero, lo que da por resultado altos niveles de parasitismo en murciélagos cavernícolas, los cuales incrementan si la cueva es utilizada como refugio permanente (Patterson et al., 2007). Lo anterior se debe, por un lado, a que las cuevas brindan las condiciones de humedad y temperatura adecuada para el desarrollo de los huevos de las moscas, y por el otro, para los parásitos es más fácil la localización rápida de un hospedero (Dick y Patterson, 2006; Graciolli y Barros de Carvalho, 2001; Patterson et al., 2007). Se considera que la presencia de ectoparásitos, así como su prevalencia, abundancia e intensidad, están más relacionadas con la selección y fidelidad que tienen ciertas especies a la cueva de San Francisco. La selección de un refugio y la permanencia de los individuos de las diferentes especies de hospederos en el refugio obedecen a factores intrínsecos de los murciélagos; por ejemplo, las necesidades metabólicas, preferencia por sitios descubiertos o cerrados, experiencia previa, economía energética con relación al tamaño corporal, organización social y selección sexual. Existen también factores extrínsecos a los murciélagos como la forma del refugio, abundancia y disponibilidad de sitios adecuados para percha, riesgos de depredación, distribución y abundancia de los recursos alimentarios, ambiente físico, entre otros (Brigham, Vonhof, Barclay y Gwilliam, 1997; Fenton, 1970; Gaisler, 1979; Humphrey, 1975; Kerth, Weissmann y König, 2000; Kunz, 1982; Ortiz-Ramírez, Lorenzo, Naranjo y León-Paniagua, 2006). En este estudio se concluye que las especies de murciélagos que más prevalencia y abundancia de ectoparásitos presentaron son aquellas que utilizan la cueva como refugio principal, mientras que las otras utilizan la cueva como refugio alterno para llevar a cabo funciones como la gestación, la hibernación, el descanso y la digestión.
La cueva de San Francisco desempeña un papel importante en la ecología de los murciélagos que la ocupan, como señalan otros autores (Álvarez-Castañeda y Álvarez, 1991; Arita, 1993; Chávez, 2008), quienes indican que este sitio alberga una importante diversidad de quirópteros. En este estudio se pudo comprobar que, además de su importancia como un refugio para los murciélagos, esta cueva también es de gran importancia para los ectoparásitos de la familia Streblidae, por lo cual se recomienda realizar más estudios que se enfoquen en la selección y uso de refugios de los murciélagos registrados en este sitio. De igual forma, también se recomienda realizar un estudio de la estructura de las colonias de murciélagos presentes en la cueva, para poder ampliar el conocimiento sobre la relación de prevalencia, abundancia e intensidad de infección con la edad de los hospederos.
Al biólogo Víctor Hugo Mendoza-Sáenz por su apoyo en el trabajo de campo. Al Dr. Hugo Perales por su ayuda con los análisis estadísticos. Al Consejo Nacional de Ciencia y Tecnología por la beca otorgada a la primera autora. Al Colegio de la Frontera Sur por el programa de apoyo a estudiantes de maestría. A la comunidad de Pamala y a las autoridades municipales de La Trinitaria por el permiso brindado para realizar el trabajo en la cueva de San Francisco. Al M. en C. Antonio García-Méndez por las revisiones realizadas a este documento desde su primera versión.
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.