



Guerrero es el cuarto estado más biodiverso en México, incluyendo 930 especies de vertebrados en su territorio. Sin embargo, ha mostrado una pérdida significativa de hábitats naturales originales. Se hace evidente la necesidad de generar información sobre los procesos involucrados en pérdida de hábitats naturales, así como identificar los impactos que tienen tales pérdidas sobre la distribución de especies. Se generaron modelos de la distribución potencial del hábitat adecuado para 47 especies, mediante la aplicación del método MaxEnt para la predicción de su distribución geográfica y se refinaron, a su vez, a modelos de la distribución actual de hábitats adecuados. Se analizó la relación entre la amplitud de la distribución potencial de hábitats adecuados y la proporción de hábitat natural remanente para cada especie, tanto a nivel estatal (Guerrero) como nacional (México), y se identificaron zonas de Guerrero necesarias para alcanzar metas fijas de conservación de las especies. Los resultados no mostraron una correlación significativa entre la amplitud de la distribución potencial de hábitats adecuados de las especies y las áreas remanentes de hábitats originales a nivel estatal o nacional. De hecho, existen diferencias significativas en las proporciones de hábitat remanente entre ambas escalas. Esto sugiere que la escala es un factor importante para establecer estrategias de conservación para las mismas especies a nivel local, regional o nacional.
The State of Guerrero is ranked fourth in terms of biodiversity in Mexico, including 930 vertebrate species in its territory. However, Guerrero shows significant loss of its original natural habitats. It is evident the need to generate information on the processes involved in loss of natural habitats and to identify the impact on the distribution of species. We modeled the potential distribution of suitable habitat for 47 species of mammals using MaxEnt, and those were further refined to produce models of the current distribution of suitable habitats. The relationship between the amplitude of the potential distribution of suitable habitats and the proportion of remaining natural habitat for each species were examined, both at the state (Guerrero) and nationwide levels (Mexico), and there were identified areas of Guerrero needed to achieve fixed conservation goals. The results showed no significant correlation between the amplitude of species potential distribution of suitable habitats and species distributions of remaining areas of original habitat, neither at the state or national scales. In fact, there are significant differences in the proportions of remaining habitat between the 2 scales, suggesting that scale is an important factor in establishing conservation strategies for the same species at local, regional or national scales.
Guerrero es el cuarto estado con mayor diversidad biológica en México, incluyendo 270 especies de anfibios y reptiles (Ochoa-Ochoa y Flores-Villela, 2006), 545 de aves (Almazán- Núñez y Navarro, 2000; Navarro, 1998) y 115 de mamíferos terrestres (Almazán-Catalán, Sánchez-Hernández y Romero- Almaraz, 2005). Sin embargo, a pesar de la elevada diversidad biológica del estado, existen pocos trabajos publicados sobre la distribución e historia natural de los mamíferos (Almazán-Catalán et al., 2005; Almazán-Catalán et al., 2009; Cervantes-Reza, Ramírez-Vite, Ramírez-Vite y Ballesteros, 2004; Cuervo-Robayo y Monroy-Vilchis, 2012; Jiménez-Almaraz, Juárez-Gómez y León-Paniagua, 1993; León-Paniagua y Romo-Vázquez, 1993; Ramírez-Pulido y Armella, 1987; Ramírez- Pulido, Armella y Castro-Campillo, 1993).
En contraste con su elevada biodiversidad, Guerrero cuenta con menos del 0.1% de su territorio decretado como área protegida (Bezauri-Creel, Torres, Ochoa y Castro-Campos, 2012) y se calcula que, a la fecha, ha perdido alrededor del 32% de su hábitat natural (Semarnat, 2009) y, menos del 30% de los hábitats naturales actuales pueden ser identificados como vegetación primaria (Semarnat, 2008). Anualmente, se pierde entre el 0.5 y 0.7% de la cobertura de bosques y 2.4 y 2.7% de selvas tropicales, además de encontrarse entre los estados con mayor fragmentación de bosques y selvas en México (22-24%), con una tasa anual entre 23.7 y 36.3% de sobre-pastoreo (Semarnat, 2008). Guerrero se ubica como uno de los estados con mayor marginación social y pobreza en México, situación que se traduce en fuertes presiones socioeconómicas para cambiar el uso de suelo de áreas con vegetación natural remanente a sistemas agropecuarios (Deininger y Minten, 1999; Figueroa, Sánchez- Cordero, Meave y Trejo, 2009).
En este escenario, la pérdida de hábitats naturales que impacta negativamente a la biodiversidad puede concebirse como un proceso cuantificable de disminución y modificaciones de la riqueza de especies, sus abundancias poblacionales, diversidad genética y sus patrones de distribución (Best, Bergin y Freemark, 2001; Challenger y Dirzo, 2009; Fahrig, 2003; Steffan-Dewenter, Munzenberg, Burger, Thies y Tscharntke, 2002; Venier y Fahrig, 1996). Diversos estudios han relacionado índices de pérdida de hábitats –representados por los principales tipos de vegetación– con la pérdida de la diversidad biológica (Kinnaird, Sanderson, O’Brien, Wibisono y Woolmer, 2003; Myers, 1998; Toledo, Carabias y González-Pacheco, 1989). Estos estudios establecen tendencias de riesgo de los tipos de vegetación, pero no permiten discernir el impacto de la pérdida de hábitat sobre la fauna a nivel de especie (Sánchez-Cordero, Illoldi-Rangel, Linaje, Sahotra y Peterson, 2005; Sánchez-Cordero, Munguía y Peterson, 2005), que es una de las mayores amenazas a las que está expuesta la biodiversidad, al asociarse con la disminución en la abundancia poblacional, diversidad genética y extensión en la distribución de las especies (Best et al., 2001; Fahrig, 2003; Steffan-Dewenter et al., 2002; Venier y Fahrig, 1996). Adicionalmente, se tiene que considerar que en México el grado de conservación de una especie de acuerdo con la Norma Oficial Mexicana (Nom-Ecol-059-2010), considera el riesgo en el que se encuentra la especie tomando en cuenta criterios a escala nacional, en algunos de los casos criterios numéricos incluidos en el método de evaluación de riesgo, que considera variables a esa misma escala (MER; Sánchez et al., 2007). Con este método, se evalúa el riesgo de extinción de las especies utilizando 4 criterios: 1) la amplitud de su distribución geográfica respecto al área total del país; 2) la calidad del hábitat en el que la especie se distribuye respecto a los requerimientos; 3) la vulnerabilidad intrínseca del taxón; 4) el impacto negativo de las actividades humanas sobre la especie (Sánchez et al., 2007). Sin embargo, estos criterios usualmente se consideran a nivel país y se ha demostrado que la pérdida de hábitat, uno de los impactos negativos provocado por las actividades humanas sobre las especies, difiere dependiendo de la región en donde estas se distribuyen (Sánchez-Cordero et al., 2009; Sánchez-Cordero, Illoldi-Rangel et al., 2005; Sánchez-Cordero, Munguía et al., 2005). Por tanto, la escala a la que se determina este impacto podría modificar la medida de intensidad del mismo.
Tomando en consideración lo anterior, el modelado de nicho ecológico o modelado de la distribución potencial de especies, es un enfoque teórico-metodológico útil para determinar el impacto de la pérdida de hábitat, a nivel de especie por especie, a distintas escalas de análisis (Elith y Leathwick, 2009; Mendoza, Fay y Dirzo, 2005; Ortega-Huerta y Peterson, 2004; Peterson, Egbert, Sánchez-Cordero y Price, 2000; Sánchez-Cordero et al., 2009; Sánchez-Cordero, Illoldi-Rangel et al., 2005; Sánchez-Cordero, Munguía et al., 2005). Durante las últimas décadas, ha aparecido una amplia variedad de enfoques similares, para generar modelos de distribución de especies (MDE): modelos bioclimáticos, proyecciones climáticas, modelos de hábitats adecuados/selección de hábitat, modelos de distribución de especies/hábitats, funciones de selección de recursos, modelos del nicho ecológico de las especies (MNE), análisis de gradientes, entre otros (Elith y Leathwick, 2009; Hirzel y Lay, 2008). Todos estos enfoques de modelado pueden definirse como modelos empíricos que relacionan observaciones de campo con variables ambientales de predicción, con base en superficies estadísticas o de respuesta esperada (i. e., derivada teóricamente) (Guisan y Zimmermann, 2000). Este enfoque de modelado predictivo de la distribución de especies o hábitats, se ha convertido en un método muy poderoso para probar hipótesis ecológicas sobre la distribución de las especies y para evaluar los posibles impactos de los cambios ambientales sobre tales distribuciones (Guisan y Hofer, 2003).
El modelado de nicho ecológico (MNE) es la denominación más comúnmente utilizada para referirse al marco conceptual que sustenta la aplicación de tales enfoques. En la definición de nicho existen 2 enfoques contrastantes propuestos por Leibold (1995) y que se relacionan con: 1) los requerimientos ambientales de las especies (Grinnell, 1917; Hutchinson, 1957) y 2) los impactos o función que las especies tienen en sus ambientales (Elton, MacArthur o Levins) (Guisan y Thuiller, 2005).
La definición de nicho propuesta por Chase y Leibold (2003) –en el sentido que el nicho de las especies está determinado por las condiciones ambientales que permiten a las especies satisfacer sus requerimientos mínimos para que las tasa de nacimientos de poblaciones locales sea igual o mayor a la de su tasa de muertes– parece ser la traducción moderna de la formulación original de Grinnell (Araujo y Guisan, 2006). Al mismo tiempo, existe el consenso que el modelado de nicho tiene como base conceptual las definiciones de Hutchinson de nicho fundamental y nicho realizado, aunque persisten las diferencias entre autores para interpretar si los modelos de nicho generados corresponden a alguna de tales versiones de nicho (Araujo y Guisan, 2006).
Los conceptos de nicho fundamental y nicho realizado nos ayudan a diferenciar si una predicción es resultado de restricciones fisiológicas teóricas, o si bien, es una simulación derivada de datos recabados en campo; en el caso de este último tipo de modelos –modelos estáticos de distribución derivados de superficies de respuesta– se considera que muy probablemente están prediciendo el nicho realizado o ecológico de las especies (Austin, 2002; Guisan y Zimmermann, 2000). En contraste, Peterson (2001) consideró que el nicho fundamental puede ser modelado mediante el examen del intervalo completo de la distribución de las especies (por ejemplo, abarcando la multiplicidad de comunidades en las que se encuentran las especies). Zaniewski, Lehmann y Overton (2002) también consideraron que los métodos de predicción basados solamente en presencias predicen distribuciones de especies, parecidas más a su nicho fundamental, mientras que los modelos basados en datos de presencia y ausencia reflejarían la distribución actual que se deriva del nicho realizado. Brotons, Thuiller, Araujo y Hirzel (2004), por su parte, consideran que ambos métodos proveen diferentes estimaciones del nicho realizado.
La discusión de la relación entre los modelos de nicho y la distribución de las especies ha estado caracterizada por dificultades semánticas, conceptuales y técnicas; algunos autores (Pearman, Guisan, Broennimann y Randin, 2008; Peterson, 2006; Soberón, 2007) sugieren que los modelos de nicho ecológico se aproximan más al modelado del nicho fundamental. Sin embargo, no queda clara la manera en que los métodos llamados “modelado del nicho ecológico” manejan las dificultades para describir ese nicho fundamental, a partir de la utilización de observaciones del paisaje asociadas a la presencia de las especies (Elith y Leathwick, 2009). De acuerdo con Pulliam, (2000), la aplicación del enfoque MNE debe tomar en cuenta, no solamente las bases conceptuales de nicho fundamental y realizado, sino también la dinámica “fuente-sumidero” y las situaciones de dispersión limitada, que se tendría que analizar con características intrínsecas de las especies, como su vagilidad y variables relacionadas con el tamaño, forma y cercanía de fragmentos de hábitats naturales.
Después de hacer algunas consideraciones conceptuales en torno a la definición de nicho fundamental versus el realizado, Araujo y Guisan (2006) cuestionan la pertinencia de hacer tal diferenciación, y proponen la posibilidad de ignorar tales conceptos para aceptar que cualquier caracterización de nicho será una descripción incompleta de los factores bióticos y abióticos, que posibilitan a las especies satisfacer sus requerimientos ecológicos. Jiménez-Valverde, Lobo y Hortal (2008) proponen un modelo conceptual a manera de gradiente, en cual se describe la dependencia que tienen los enfoques de distribución “potencial” o “realizada”, en la aplicación de marcos analíticos diferentes, así como la utilización y aplicación de datos de calibración y validación adecuados a cada enfoque.
En este estudio se adopta el enfoque de modelar los hábitats adecuados potenciales de las especies a partir del modelado de su distribución potencial (Guisan y Zimmermann, 2000). Una de las caracterizaciones del modelado del nicho ecológico de las especies, consiste en generar modelos sobre lo adecuado o idóneo de sus hábitats. Por ejemplo, Hirzel, Helfer y Metral (2001), mediante el modelado del nicho fundamental de una especie virtual, determinaron la idoneidad del hábitat bajo diferentes escenarios históricos de colonización; con dispersión y poco abundante, en equilibrio y muy abundante. Si los modelos de nicho ecológico relacionan un conjunto de variables ambientales con la aptitud de las especies, los modelos de idoneidad del hábitat pueden ser considerados como aplicaciones operativas de los primeros, ya que las variables ambientales se relacionan con la probabilidad de presencia de las especies (Hirzel y Lay, 2008). Los objetivos para obtener modelos de idoneidad del hábitat, descansan en la reconstrucción del nicho realizado (Grinnell, 1917) y su predicción derivada (Guisan y Zimmermann, 2000). La consideración de los modelos de distribución de especies como modelos de distribución de hábitats específicos, resulta pertinente si consideramos la concepción de nicho de Grinnell (1917), en el sentido de que este consiste básicamente en la subdivisión de su hábitat, en la cual la especie encuentra aquellas condiciones que le posibilitan su sobrevivencia y reproducción (Wiens, Stralberg, Jongsomjita, Howell y Snyder, 2009).
Aplicando un procedimiento mediante el cual, modelos de distribución de especies llamados “potenciales” son reducidos espacialmente a los hábitats naturales remanentes, Sánchez-Cordero, Illoldi-Rangel et al. (2005) concluyeron que más de una cuarta parte de las especies endémicas de mamíferos terrestres de México, perdieron >50% de su distribución potencial debido a pérdida de hábitat. Además, no observaron una relación entre esta tendencia y el estatus de riesgo asignado por la Norma Oficial Mexicana Nom-059-Ecol-2001; la pérdida de hábitat natural se relaciona más con la región del país en la que se distribuyen, que con la amplitud de su distribución. Estos estudios sugieren la necesidad de realizar análisis de los impactos de la pérdida de hábitats naturales a nivel de especie por especie y, a escalas regionales, no solamente la escala de país, que permitan anticipar los riesgos de las extinciones locales, así como el diseño e implementación de acciones de conservación local. Adicionalmente, es necesario considerar que el costo de conservación se incrementa conforme avanza el proceso de pérdida de hábitat natural (Fuller, Sánchez-Cordero, Illoldi-Rangel, Linaje y Sarkar, 2006). La planeación sistemática de la conservación es un enfoque estructurado, en varios pasos, para cartografiar sistemas de áreas para la conservación en las que se busca representatividad, complementariedad y persistencia de la biodiversidad (Margules y Pressey, 2000; Margules y Sarkar, 2009; Sarkar y Margules, 2002). Este trabajo aplica el sistema ConsNet (Ciarleglio, Barnes y Sarkar, 2009; Ciarleglio, Sarkar y Barnes, 2008), el cual permite analizar información sobre la distribución de múltiples especies en un contexto geográfico y también permite buscar soluciones de conservación que incluyan criterios de representatividad y complementariedad.
El objetivo general de este trabajo es cuantificar, a escala estatal (Guerrero) y nacional (México), la disponibilidad de hábitats adecuados asociados a la distribución de especies de mamíferos terrestres y se evalúa la manera en la que este factor afecta la selección de áreas de conservación para mamíferos en Guerrero.
Materiales y métodosSitios de presencia de las especies (recolectas científicas)Considerando que la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio) lleva a cabo controles de la calidad de las bases de datos que se integran al Sistema Nacional de Información sobre Biodiversidad (Snibhttp://www.conabio.gob.mx/biotica5/documents/cursoMarzo2013/Control%20de%20Calidad%20SIB_2013.pdf), se solicitó a dicha comisión datos sobre localidades de colecta de mamíferos terrestres a nivel nacional (proyectos de origen de los registros: Altamirano-González, Guzmán-Hernández, Luna-Reyes, Riechers-Pérez y Vidal-López, 2007; Álvarez-Solórzano y López-Vidal, 1998; Briones-Salas, 2006; Ceballos, 2006; Cervantes-Reza, 2005; León-Paniagua, 1999; León-Paniagua, Rojas-Soto y Ortiz, 2001; López-Vidal, 2006; López-Vidal et al., 2008; López-Wilchis, 1998; Lorenzo-Monterrubio, Bolaños-Citalán y Barragán-Torres, 2005; Martínez-Vázquez, 2001; Miranda, 1997; Morales-Pérez, 2005; Pozo de la Tijera, 1997; Ramírez- Pulido, 1999; Ramírez-Pulido, 2007; Sánchez-Cordero, 2006). A pesar de la rigurosa revisión que realiza la Conabio para incluir los datos en el SNIB, se identificaron algunas inconsistencias taxonómicas y geográficas, por lo que se revisó la taxonomía con base en Wilson y Reeder (2005) y la congruencia geográfica de los registros (Patterson et al., 2007), excluyéndose o modificándose todas aquellas especies o registros que: i) fueran sinonimias; ii) que no tuvieran datos de referencia geográfica (longitud y latitud) con una resolución mayor a 2 decimales de grado; iii) que la especie no hubiera sido registrada previamente para el estado de Guerrero; iv) que no fuera nativa de México; v) que los registros no coincidieran con la distribución actualmente reconocida de la especie; vi) que tuvieran menos de 10 registros, y vii) que fueran registros duplicados, es decir, múltiples registros para una misma localidad (Elith et al., 2011). La depuración de datos de presencia de las especies resultó en 9 324 registros que fueron utilizados para modelar la distribución del hábitat potencial de 47 especies de mamíferos terrestres documentados en Guerrero; esta cifra representa el 40% de las 115 especies de mamíferos terrestres que se distribuyen en el estado.
Variables ambientales de predicciónLas coberturas utilizadas para modelar la distribución de los hábitats potenciales de las especies, consistieron en 19 variables bioclimáticas del proyecto WorldClim, construidas con base en la interpolación del promedio mensual de datos de estaciones climáticas, obtenidos del periodo de 1950-2000 (http://www. worldclim.org/current), y 4 variables topográficas del proyecto Hydro1k (http://edc.usgs.gov/products/elevation/gtopo30/hydro/), con una resolución de ≈1km2 (tabla 1).
Variables bioclimáticas y topográficas utilizadas para realizar los modelos de distribución potencial.
| Variables bioclimáticas | Variables topográficas |
|---|---|
| 1. Temperatura media anual | 20. Aspecto |
| 2. Rango de temperatura media diurna | 21. Elevación |
| 3. Isotermalidad | 22. Pendiente |
| 4. Estacionalidad de temperatura (desviación estándar *100) | 23. Índice topográfico compuesto |
| 5. Temperatura máxima del mes más cálido | |
| 6. Temperatura mínima del mes más frío | |
| 7. Rango de temperatura anual | |
| 8. Temperatura media del trimestre más húmedo | |
| 9. Temperatura media del trimestre más seco | |
| 10. Temperatura media del trimestre más cálido | |
| 11. Temperatura media del trimestre más frío | |
| 12. Precipitación anual | |
| 13. Precipitación del mes más húmedo | |
| 14. Precipitación del mes más seco | |
| 15. Estacionalidad de precipitaciones | |
| 16. Precipitación del trimestre más húmedo | |
| 17. Precipitación del trimestre más seco | |
| 18. Precipitación del trimestre más cálido | |
| 19. Precipitación del trimestre más frío |
La preparación y el procesamiento de los modelos de distribución se llevaron a cabo con el software ArcGis 9.3 (Esri). De acuerdo con Elith et al. (2011), es recomendable restringir el área de modelado a aquella que se considera como accesible para la especie (M). Tomando lo anterior en consideración, cuando es factible es importante utilizar como límite de modelado para cada especie áreas que incluyan condiciones geográficas y ecosistemas que se puedan considerar como unidades que no contienen barreras que impidan su dispersión. Puesto que las Ecorregiones Terrestres de México (Inegi, Conabio, INE, 2008), son el resultado de un análisis reciente que incluye una serie de esfuerzos de análisis de expertos sobre trazos y límites correspondientes a unidades geográficas con flora, fauna y ecosistemas característicos, se utilizaron estos atributos para intersectarse con la distribución conocida (registros) de cada una de las 47 especies de mamíferos que cumplieron con el criterio de contar al menos con 10 registros en su distribución reconocida (Patterson et al., 2007); el resultado fue un grupo de 47 máscaras que representan la extensión accesible para cada una de las especies (Barve et al., 2011). Con cada una de estas máscaras, se extrajeron las 23 variables de predicción, obteniéndose un grupo de variables con distinta extensión, para cada una de las especies a modelar.
Modelado de distribución de especiesEl software MaxEnt (Phillips, Anderson y Schapire, 2006), versión 3.3.1 k, fue el sistema de cómputo utilizado para producir la predicción de distribuciones de especies incluidas (tabla 2). Este programa utiliza el principio de máxima entropía, estimando la distribución a lo largo de un espacio geográfico. Elith et al. (2011) hacen una caracterización de la manera que MaxEnt opera, con base en la comparación de densidades de probabilidades en el espacio de covariables; si se conoce la densidad condicional de covariables en sitios de presencia de determinada especie (f1 [z]), la densidad marginal de covariables a lo largo del área de estudio (f[z]) y la prevalencia, entendida como la proporción de presencias en el total de la muestra (Pr[y= 1]), es posible calcular la probabilidad condicional de la presencia. Sin embargo, como MaxEnt trabaja solo con datos de presencia, primero realiza transformaciones de las covariables: lineal, cuadrática, bisagra y de producto; con valores comprendidos entre 0 y 1. Con estas transformaciones se estima la relación de la densidad condicional de “características” en sitios de presencia (f1[z]), dividido entre la densidad marginal de características a lo largo del área de estudio (f[z]), es decir (f1[z]/f[z]), lo que se considera el núcleo de los modelos que produce MaxEnt y se le conoce como salida de tipo “raw”, en las cuales se estima la relativa idoneidad de un sitio respecto a otro. Una vez que se tiene esta información, se utiliza una función logarítmica [n(z)= log(f1(z)/(f(z)); “logistic output”], que permite tener un acercamiento a la probabilidad de presencia en sitios con las condiciones “típicas” para la especie, el cual es conocido como el parámetro τ. De esta manera, la probabilidad de presencia presentada para cada una de las celdas del espacio geográfico en la salida logarítmica de MaxEnt, o modelo de distribución potencial de las especies, está dada por: Pr(y= 1/z)= (τen(z)-r/1-t+: τen(z)-r), en donde τ es la probabilidad de presencia en sitios con condiciones típicas, en(z) es la relación de f1(z)/(fz), r es la entropía relativa del estimado de MaxEnt de f1(z) con respecto a f(z).
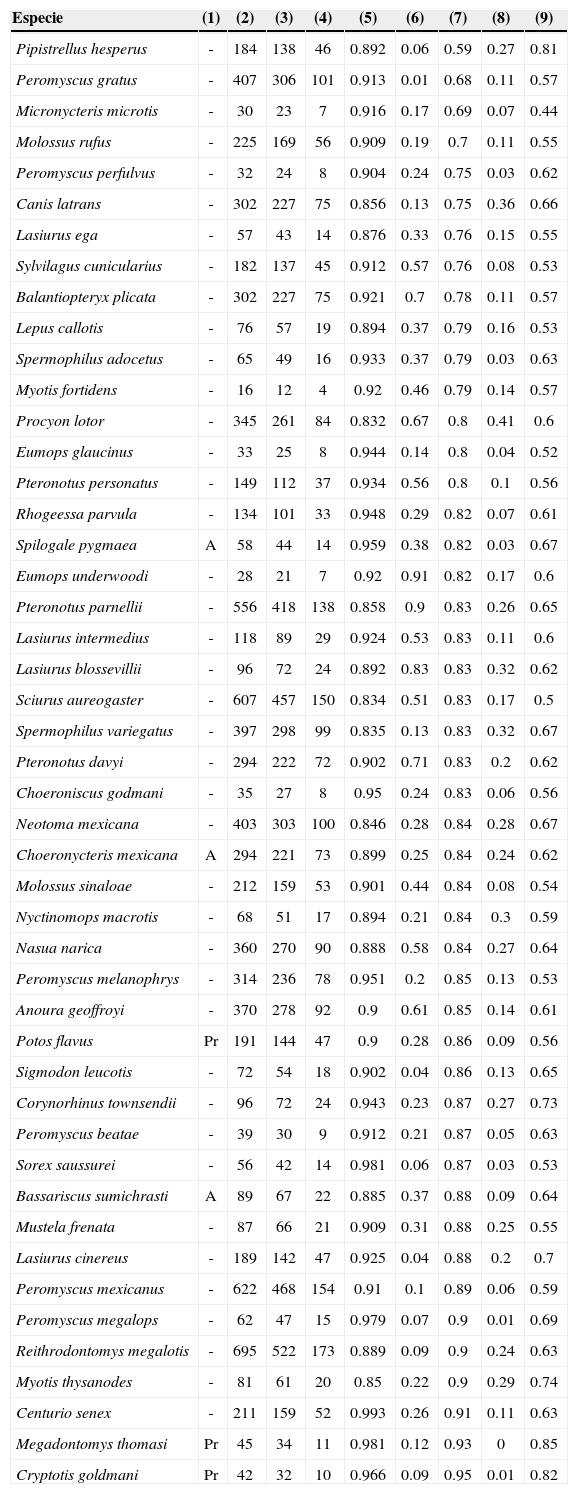
Información sobre las especies de mamíferos incluidas en los análisis: (1) categoría de riesgo de acuerdo con la Norma Oficial Mexicana (Nom-Ecol-059-2010); donde A= Especie amenazada y Pr= Especie en peligro de extinción; (2) total de registros; (3) número de registros utilizados para generar los modelos de distribución; (4) número de registros utilizados para validación de modelos; (5) área bajo la curva del análisis “ROC”; (6) proporción distribución potencial/área total para Guerrero; (7) proporción distribución actual/ distribución potencial para Guerrero; (8) proporción distribución potencial/ área total para México; (9) proporción distribución actual/ distribución potencial para México. La lista de especies aparece en orden ascendente respecto a la columna (7).
| Especie | (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | (9) |
|---|---|---|---|---|---|---|---|---|---|
| Pipistrellus hesperus | - | 184 | 138 | 46 | 0.892 | 0.06 | 0.59 | 0.27 | 0.81 |
| Peromyscus gratus | - | 407 | 306 | 101 | 0.913 | 0.01 | 0.68 | 0.11 | 0.57 |
| Micronycteris microtis | - | 30 | 23 | 7 | 0.916 | 0.17 | 0.69 | 0.07 | 0.44 |
| Molossus rufus | - | 225 | 169 | 56 | 0.909 | 0.19 | 0.7 | 0.11 | 0.55 |
| Peromyscus perfulvus | - | 32 | 24 | 8 | 0.904 | 0.24 | 0.75 | 0.03 | 0.62 |
| Canis latrans | - | 302 | 227 | 75 | 0.856 | 0.13 | 0.75 | 0.36 | 0.66 |
| Lasiurus ega | - | 57 | 43 | 14 | 0.876 | 0.33 | 0.76 | 0.15 | 0.55 |
| Sylvilagus cunicularius | - | 182 | 137 | 45 | 0.912 | 0.57 | 0.76 | 0.08 | 0.53 |
| Balantiopteryx plicata | - | 302 | 227 | 75 | 0.921 | 0.7 | 0.78 | 0.11 | 0.57 |
| Lepus callotis | - | 76 | 57 | 19 | 0.894 | 0.37 | 0.79 | 0.16 | 0.53 |
| Spermophilus adocetus | - | 65 | 49 | 16 | 0.933 | 0.37 | 0.79 | 0.03 | 0.63 |
| Myotis fortidens | - | 16 | 12 | 4 | 0.92 | 0.46 | 0.79 | 0.14 | 0.57 |
| Procyon lotor | - | 345 | 261 | 84 | 0.832 | 0.67 | 0.8 | 0.41 | 0.6 |
| Eumops glaucinus | - | 33 | 25 | 8 | 0.944 | 0.14 | 0.8 | 0.04 | 0.52 |
| Pteronotus personatus | - | 149 | 112 | 37 | 0.934 | 0.56 | 0.8 | 0.1 | 0.56 |
| Rhogeessa parvula | - | 134 | 101 | 33 | 0.948 | 0.29 | 0.82 | 0.07 | 0.61 |
| Spilogale pygmaea | A | 58 | 44 | 14 | 0.959 | 0.38 | 0.82 | 0.03 | 0.67 |
| Eumops underwoodi | - | 28 | 21 | 7 | 0.92 | 0.91 | 0.82 | 0.17 | 0.6 |
| Pteronotus parnellii | - | 556 | 418 | 138 | 0.858 | 0.9 | 0.83 | 0.26 | 0.65 |
| Lasiurus intermedius | - | 118 | 89 | 29 | 0.924 | 0.53 | 0.83 | 0.11 | 0.6 |
| Lasiurus blossevillii | - | 96 | 72 | 24 | 0.892 | 0.83 | 0.83 | 0.32 | 0.62 |
| Sciurus aureogaster | - | 607 | 457 | 150 | 0.834 | 0.51 | 0.83 | 0.17 | 0.5 |
| Spermophilus variegatus | - | 397 | 298 | 99 | 0.835 | 0.13 | 0.83 | 0.32 | 0.67 |
| Pteronotus davyi | - | 294 | 222 | 72 | 0.902 | 0.71 | 0.83 | 0.2 | 0.62 |
| Choeroniscus godmani | - | 35 | 27 | 8 | 0.95 | 0.24 | 0.83 | 0.06 | 0.56 |
| Neotoma mexicana | - | 403 | 303 | 100 | 0.846 | 0.28 | 0.84 | 0.28 | 0.67 |
| Choeronycteris mexicana | A | 294 | 221 | 73 | 0.899 | 0.25 | 0.84 | 0.24 | 0.62 |
| Molossus sinaloae | - | 212 | 159 | 53 | 0.901 | 0.44 | 0.84 | 0.08 | 0.54 |
| Nyctinomops macrotis | - | 68 | 51 | 17 | 0.894 | 0.21 | 0.84 | 0.3 | 0.59 |
| Nasua narica | - | 360 | 270 | 90 | 0.888 | 0.58 | 0.84 | 0.27 | 0.64 |
| Peromyscus melanophrys | - | 314 | 236 | 78 | 0.951 | 0.2 | 0.85 | 0.13 | 0.53 |
| Anoura geoffroyi | - | 370 | 278 | 92 | 0.9 | 0.61 | 0.85 | 0.14 | 0.61 |
| Potos flavus | Pr | 191 | 144 | 47 | 0.9 | 0.28 | 0.86 | 0.09 | 0.56 |
| Sigmodon leucotis | - | 72 | 54 | 18 | 0.902 | 0.04 | 0.86 | 0.13 | 0.65 |
| Corynorhinus townsendii | - | 96 | 72 | 24 | 0.943 | 0.23 | 0.87 | 0.27 | 0.73 |
| Peromyscus beatae | - | 39 | 30 | 9 | 0.912 | 0.21 | 0.87 | 0.05 | 0.63 |
| Sorex saussurei | - | 56 | 42 | 14 | 0.981 | 0.06 | 0.87 | 0.03 | 0.53 |
| Bassariscus sumichrasti | A | 89 | 67 | 22 | 0.885 | 0.37 | 0.88 | 0.09 | 0.64 |
| Mustela frenata | - | 87 | 66 | 21 | 0.909 | 0.31 | 0.88 | 0.25 | 0.55 |
| Lasiurus cinereus | - | 189 | 142 | 47 | 0.925 | 0.04 | 0.88 | 0.2 | 0.7 |
| Peromyscus mexicanus | - | 622 | 468 | 154 | 0.91 | 0.1 | 0.89 | 0.06 | 0.59 |
| Peromyscus megalops | - | 62 | 47 | 15 | 0.979 | 0.07 | 0.9 | 0.01 | 0.69 |
| Reithrodontomys megalotis | - | 695 | 522 | 173 | 0.889 | 0.09 | 0.9 | 0.24 | 0.63 |
| Myotis thysanodes | - | 81 | 61 | 20 | 0.85 | 0.22 | 0.9 | 0.29 | 0.74 |
| Centurio senex | - | 211 | 159 | 52 | 0.993 | 0.26 | 0.91 | 0.11 | 0.63 |
| Megadontomys thomasi | Pr | 45 | 34 | 11 | 0.981 | 0.12 | 0.93 | 0 | 0.85 |
| Cryptotis goldmani | Pr | 42 | 32 | 10 | 0.966 | 0.09 | 0.95 | 0.01 | 0.82 |
Parametrización y evaluación de modelos. Para la generación de los modelos se utilizó el 75% de los registros de 47 especies de mamíferos como puntos de entrenamiento o calibración, mientras que el 25% restante de los registros fueron utilizados como puntos de prueba independientes para validar los modelos de distribución (Pawar et al., 2007). Se utilizó MaxEnt con las especificaciones o características estándar de modelado, las cuales han demostrado producir resultados robustos (Elith et al., 2011; Elith y Leathwick, 2009; Morueta- Holme, Flojgaard y Svenning, 2010; Phillips y Dudlk, 2008). Al no contar con un criterio robusto para definir sesgos de colecta, no se incluyó dicha variable en la generación de los modelos. Sin embargo, todos los modelos se validaron utilizando el análisis incluido en MaxEnt, correspondiente al área bajo la curva (AUC, por sus siglas en inglés) de la característica de receptor operativo (ROC, por sus siglas en inglés). Los modelos que obtuvieron AUC> 0.75, fueron considerados como significativamente robustos (Morueta-Holme et al., 2010; Pawar et al., 2007). MaxEnt también provee análisis estadísticos básicos, con base en un porcentaje de datos de presencia que se asigna, para utilizarse como puntos de prueba para el modelo. Estos análisis proporcionan umbrales potenciales para ser utilizados en la transformación de los modelos continuos a modelos binarios (presencia/ausencia). En este trabajo se aplicó el umbral del décimo percentil de los puntos de entrenamiento, que se basa en el intervalo de valores que incluye al 90% de los registros de presencia y excluye al 10% restante, como fuera del intervalo (Morueta-Holme et al., 2010). Este criterio permite excluir valores que hayan sido mal referenciados, mal identificados, o en nuestro caso, registros que no fueron filtrados por la fecha de colecta, es decir, aquellos que no correspondieron a las condiciones actuales en el modelado (Liu, Berry, Dawson y Pearson, 2005; Morueta-Holme et al., 2010).
Posprocesamiento de modelosLos modelos de distribución de especies generados por MaxEnt, cuya extensión corresponde a su área accesible delimitada por las ecorregiones de México, en donde la especie tiene representación, fueron acotados espacialmente a la extensión geográfica de Guerrero, mediante el polígono correspondiente a ese estado (Conabio, 2011). Se generaron 2 archivos, 1 para cada escala, mediante el programa Maxent to Consnet, para ser analizados mediante el programa computacional de planeación sistemática de la conservación ConsNet. Este programa fue seleccionado porque permite incluir información espacial, a cualquier escala, sobre la distribución de especies, hacer análisis del área de “distribución potencial” y de “distribución actual” (Sánchez-Cordero, Illoldi-Rangel et al., 2005), además de poseer algoritmos con los que se pueden buscar soluciones de representación de metas definidas de distribución especie por especie (Ciarleglio et al., 2008; Ciarleglio et al., 2009).
Modelos de distribución de hábitats adecuados a las especies (potencial versus actual)Este estudio considera a los modelos generados por MaxEnt, como modelos de la distribución de hábitats adecuados de las especies. Tomando como base conceptual el modelo de nicho ecológico de las especies propuesto por Grinnell (1917), en una primera aproximación de modelado y mediante la utilización de variables bioclimáticas y topográficas, se generaron modelos de la distribución potencial de hábitats adecuados para las especies. Sin embargo, en una segunda aproximación, las distribuciones potenciales de hábitats obtenidos se restringieron (espacialmente), a aquellas áreas que, por la cobertura del terreno, se ajustaran mejor la idoneidad de los hábitats para cada especie. Es decir, se utilizó la capa de uso de suelo y vegetación a nivel nacional disponible más actualizada para México, la Carta digital de uso de suelo y vegetación 1:250,000, Serie IV (Inegi, 2009), como información base, para crear una máscara utilizada para excluir de los modelos de distribución potencial, todos aquellos pixeles coincidentes con coberturas del terreno, con poca o nula idoneidad de hábitat para las especies: agricultura de humedad, agricultura de riego, agricultura de temporal, asentamientos humanos, zonas desprovistas de vegetación, el palmar inducido, pastizal cultivado, pastizal inducido y las zonas urbanas. Aunque este procedimiento apunta al modelado de los hábitats relacionados con el nicho realizado de las especies, lo cierto es que los modelos finales continúan relacionándose con el modelado de una parte del nicho fundamental de las especies, sobre todo, porque no se incluyen las interacciones bióticas y los procesos de dispersión y dinámica de meta-poblaciones. La consideración de los modelos de distribución de hábitat adecuados resuelve, por lo menos en parte, el dilema de referirnos a distribuciones de especies potenciales versus actuales, ya que los hábitats son intrínsecamente áreas o lugares que potencialmente pueden estar ocupados o desocupados por las especies. Por lo tanto, aquí nos referimos a la “distribución potencial” de hábitats adecuados a aquellos modelos obtenidos mediante la predicción de variables ambientales climáticas y topográficas (hábitats potenciales), mientras que la “distribución actual” de hábitats adecuados, son los mismos modelos, aunque reducidos a aquellas áreas que aún presentan la cubierta del terreno correspondiente a los hábitats adecuados a las especies (hábitats actuales).
Correlación entre distribuciones potenciales versus actuales de hábitats adecuadosBasados en la propuesta de Sánchez-Cordero, Illoldi-Rangel et al. (2005) y Sánchez-Cordero, Munguía et al. (2005), y una vez obtenida la proporción de hábitat remanente dentro de la distribución potencial de hábitat de las especies, tanto para México como para Guerrero (Conabio, 2011), se evaluó la correlación entre el área correspondiente a la distribución potencial del hábitat respecto al área de la distribución actual del hábitat de las especies. Se utilizó una prueba de correlación de Spearman porque los datos no cumplieron con la normalidad, definiendo p= 0.05 como nivel de significancia (Wilson et al., 2005; Zar, 1999), mediante el programa Statistica 10.0 (StatSoft, Inc., 2011). Finalmente, se realizó una comparación entre las proporciones de hábitats (potenciales) adecuados de las especies, para México versus Guerrero, utilizando una prueba de Wilcoxon (Zar, 1999), mediante el programa Statistica 10.0 (StatSoft, Inc., 2011).
Identificación de áreas de conservaciónMediante ConsNet, se hizo un análisis de prioridades de conservación utilizando una meta de conservación del 20% de la distribución de los hábitats de las especies bajo 2 escenarios; el primero, considerando la pérdida de hábitat en el estado (hábitats actuales), y el segundo, sin considerar este factor (hábitats potenciales) (Fuller et al., 2006). Para ambos escenarios, se buscó una solución óptima de conservación, utilizándose como objetivo de conservación la selección de la menor cantidad de área posible y la menor relación entre perímetro y área de las zonas seleccionadas (Ciarleglio et al., 2009), corriéndose tantas iteraciones como fueron necesarias, para llegar a una asíntota en la que no se encontraron nuevas soluciones óptimas, en cuanto a los criterios considerados (área y relación perímetro/ área). Para determinar la diferencia a nivel geopolítico, usualmente el considerado para realizar acciones de manejo y conservación, de las áreas seleccionadas. La mejor solución para cada uno de los 2 escenarios se superpuso a la capa de municipios de Guerrero y se cuantificó el número de pixeles seleccionados para cada uno de ellos. Para comparar ambas soluciones, realizamos una prueba de Wilcoxon (Zar, 1999), mediante el programa Statistica 10.0 (StatSoft, Inc., 2011).
ResultadosDistribución actual del hábitat adecuado de las especiesDe acuerdo con la Carta digital de uso de suelo y vegetación 1:250 000, Serie IV (Inegi, 2009), se observó que el 68% del área del estado cuenta con hábitat natural remanente. Para México, se observó una tendencia positiva no significativa entre la amplitud de la distribución potencial de los hábitats adecuados de las especies y sus distribuciones actuales (rs= 0.184, p= 0.216). También en la escala del país, 20 especies conservaron menos del 60% de vegetación natural respecto a su distribución potencial, 22 especies conservaron entre el 60 y 70%, 2 especies entre el 70 y 80% y, 3 especies conservaron entre el 80 y 90% (tabla 2); la mediana fue del 61%.
Se observó una tendencia negativa no significativa, entre la amplitud de la distribución potencial de los hábitats adecuados de las especies y sus distribuciones actuales en Guerrero (rs= –0.254, p= 0.08). Dentro del mismo ámbito estatal, de las 47 especies, solo 1 especie conservó menos del 60% de su distribución potencial, 2 especies conservaron entre el 60 y 70%, 10 especies entre el 70 y 80%, 31 especies entre el 80 y 90% y, 3 especies conservaron más del 90% de hábitat natural remanente en la distribución potencial de sus hábitats adecuados (ver tabla 2; columna (7)); la mediana fue de 83%. Adicionalmente, se encontraron diferencias significativas, al comparar la proporción de hábitats naturales que se conservan en la distribución potencial de los hábitats adecuados de las especies, entre México y Guerrero (N= 47, T= 26, Z= 5.6, p< 0.01).
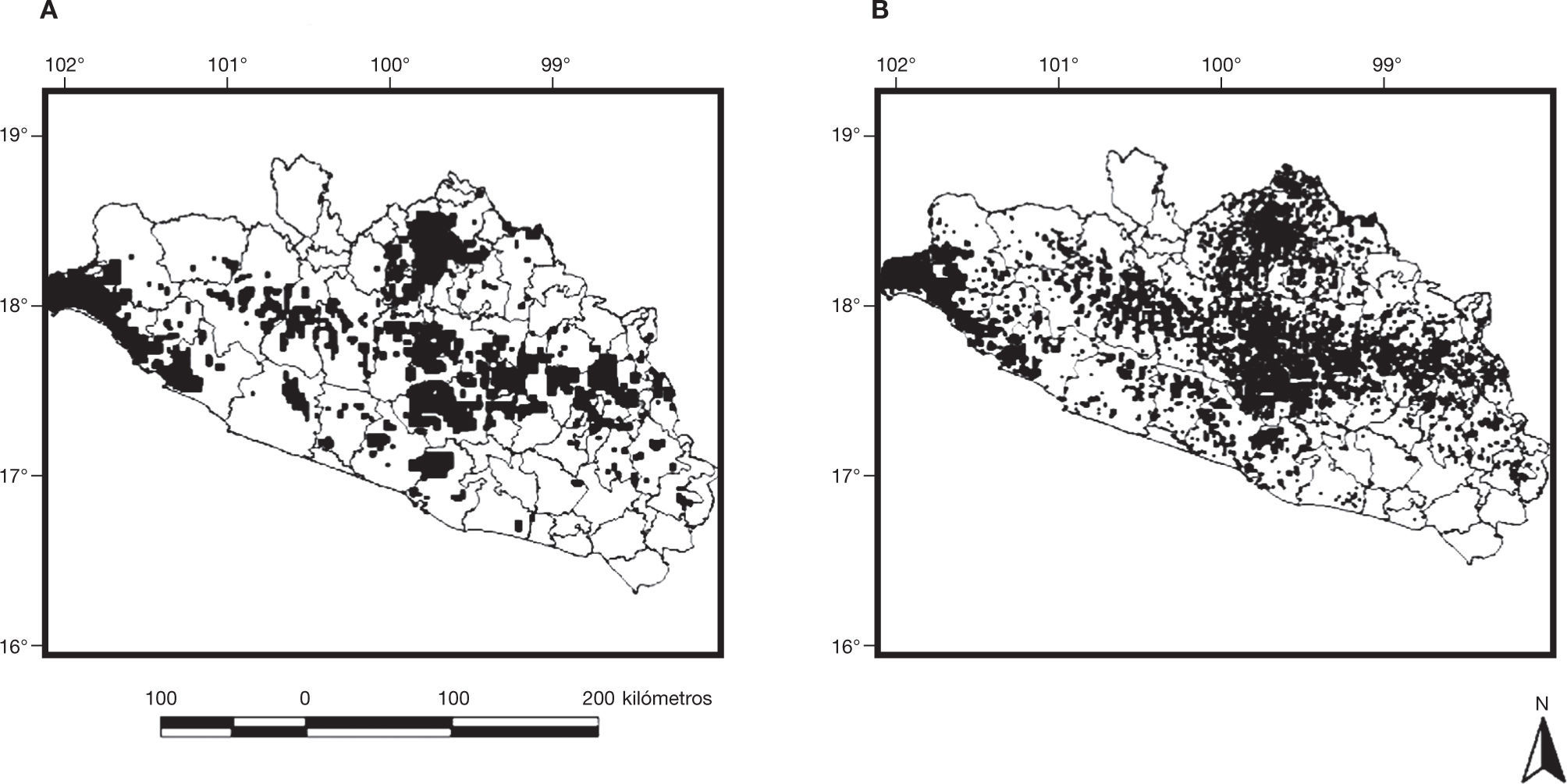
Áreas de conservaciónLa mejor solución de conservación obtenida, sin considerar la exclusión de áreas cuyos hábitats naturales se han transformado (hábitats adecuados potenciales), requirió de 11 549km2 para representar el 20% de la distribución potencial de los hábitats adecuados para las 47 especies, seleccionándose 135 áreas con un perímetro de 5445 kilómetros y una relación perímetro/ área de 0.47 (fig. 1A). La mejor solución de áreas de conservación obtenida considerando la exclusión de áreas transformadas (hábitats adecuados actuales), requirió de la misma área seleccionada, para representar el 20% de la distribución de los hábitats adecuados actuales para las 47 especies (11 549km2). Sin embargo, el número de áreas seleccionadas se incrementó a 1 212 con un perímetro de 15 599 km y una relación perímetro/ área de 1.35 (fig. 1B).
 y áreas de conservación seleccionadas considerando pérdida de hábitat natural (B).")
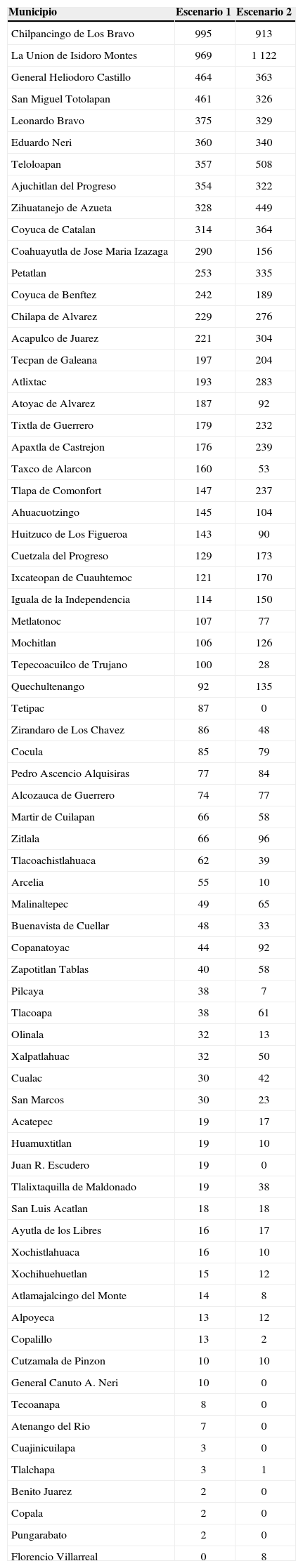
La solución de conservación en el escenario de la distribución de hábitats adecuados actuales incluyó áreas pertenecientes a 70 de los 81 municipios del estado; no se encontraron diferencias significativas al comparar la localización, de acuerdo con el municipio, de las áreas seleccionadas en ambos escenarios (N= 69, T= 1 163.5, Z= 0.263, p= 0.79). Los municipios con mayor proporción de área seleccionada fueron: Chilpancingo de Los Bravo, La Unión de Isidoro Montes de Oca, General Heliodoro Castillo, San Miguel Totolapan, Leonardo Bravo, Eduardo Neri, Teloloapan, Ajuchitlán del Progreso, Josué Azueta, Coyuca de Catalán, Coahuayutla de José María Izazaga, Petatlán, Coyuca de Benítez, Chilapa de Álvarez y Acapulco de Juárez (ver tabla 3).
Número de celdas seleccionadas en municipios del estado de Guerrero. El Escenario 1 corresponde a los modelos de distribución del hábitat adecuado potencial, sin considerar pérdida de hábitat natural; mientras que el Escenario 2 corresponde al modelado del hábitat adecuado actual, considerando pérdida de hábitat natural. El orden de la tabla es descendente de acuerdo con la columna del Escenario 1.
| Municipio | Escenario 1 | Escenario 2 |
|---|---|---|
| Chilpancingo de Los Bravo | 995 | 913 |
| La Union de Isidoro Montes | 969 | 1 122 |
| General Heliodoro Castillo | 464 | 363 |
| San Miguel Totolapan | 461 | 326 |
| Leonardo Bravo | 375 | 329 |
| Eduardo Neri | 360 | 340 |
| Teloloapan | 357 | 508 |
| Ajuchitlan del Progreso | 354 | 322 |
| Zihuatanejo de Azueta | 328 | 449 |
| Coyuca de Catalan | 314 | 364 |
| Coahuayutla de Jose Maria Izazaga | 290 | 156 |
| Petatlan | 253 | 335 |
| Coyuca de Benftez | 242 | 189 |
| Chilapa de Alvarez | 229 | 276 |
| Acapulco de Juarez | 221 | 304 |
| Tecpan de Galeana | 197 | 204 |
| Atlixtac | 193 | 283 |
| Atoyac de Alvarez | 187 | 92 |
| Tixtla de Guerrero | 179 | 232 |
| Apaxtla de Castrejon | 176 | 239 |
| Taxco de Alarcon | 160 | 53 |
| Tlapa de Comonfort | 147 | 237 |
| Ahuacuotzingo | 145 | 104 |
| Huitzuco de Los Figueroa | 143 | 90 |
| Cuetzala del Progreso | 129 | 173 |
| Ixcateopan de Cuauhtemoc | 121 | 170 |
| Iguala de la Independencia | 114 | 150 |
| Metlatonoc | 107 | 77 |
| Mochitlan | 106 | 126 |
| Tepecoacuilco de Trujano | 100 | 28 |
| Quechultenango | 92 | 135 |
| Tetipac | 87 | 0 |
| Zirandaro de Los Chavez | 86 | 48 |
| Cocula | 85 | 79 |
| Pedro Ascencio Alquisiras | 77 | 84 |
| Alcozauca de Guerrero | 74 | 77 |
| Martir de Cuilapan | 66 | 58 |
| Zitlala | 66 | 96 |
| Tlacoachistlahuaca | 62 | 39 |
| Arcelia | 55 | 10 |
| Malinaltepec | 49 | 65 |
| Buenavista de Cuellar | 48 | 33 |
| Copanatoyac | 44 | 92 |
| Zapotitlan Tablas | 40 | 58 |
| Pilcaya | 38 | 7 |
| Tlacoapa | 38 | 61 |
| Olinala | 32 | 13 |
| Xalpatlahuac | 32 | 50 |
| Cualac | 30 | 42 |
| San Marcos | 30 | 23 |
| Acatepec | 19 | 17 |
| Huamuxtitlan | 19 | 10 |
| Juan R. Escudero | 19 | 0 |
| Tlalixtaquilla de Maldonado | 19 | 38 |
| San Luis Acatlan | 18 | 18 |
| Ayutla de los Libres | 16 | 17 |
| Xochistlahuaca | 16 | 10 |
| Xochihuehuetlan | 15 | 12 |
| Atlamajalcingo del Monte | 14 | 8 |
| Alpoyeca | 13 | 12 |
| Copalillo | 13 | 2 |
| Cutzamala de Pinzon | 10 | 10 |
| General Canuto A. Neri | 10 | 0 |
| Tecoanapa | 8 | 0 |
| Atenango del Rio | 7 | 0 |
| Cuajinicuilapa | 3 | 0 |
| Tlalchapa | 3 | 1 |
| Benito Juarez | 2 | 0 |
| Copala | 2 | 0 |
| Pungarabato | 2 | 0 |
| Florencio Villarreal | 0 | 8 |
La base conceptual del presente estudio parte del reconocimiento de que existen varias formas para referirse al objeto y tipo de modelos, que tienen como meta predecir la probabilidad de presencia de las especies, a partir de variables ambientales (Guisan y Zimmermann, 2000). Tales enfoques incluyen al “modelado de la distribución de las especies”, el “modelado del nicho ecológico de las especies” y “modelado de la idoneidad del hábitat”, para mencionar los más reconocidos (Elith y Leathwick, 2009). Este estudio adopta las definiciones como la de Whittaker y Likens (1973), quienes consideraban que el concepto propuesto por Grinnell (1917) enfatizaba al hábitat de las especies, razón por la cual se hablaba de un “nicho de lugar” o “nicho de hábitat”. De manera particular, se considera que los modelos propuestos en este estudio se ajustan mejor a las características de los modelos del hábitat adecuado de las especies (Hirzel y Lay, 2008), debido a la naturaleza de las variables de predicción utilizadas, las escalas de análisis abordadas, así como el algoritmo de predicción aplicado. De esta manera, utilizamos las categorías “potencial” y “actual” para diferenciar la idoneidad predicha de los hábitats de las especies, dados los cambios en las condiciones históricas de la cobertura vegetal (Sánchez-Cordero et al., 2009; Sánchez-Cordero, Illoldi-Rangel et al., 2005; Sánchez-Cordero, Munguía et al., 2005). La utilización de la categoría “hábitat adecuado”, en lugar de “distribución de las especies” es específica, se adapta a los análisis de selección de sitios para la conservación y tiene conceptualmente una rigurosidad científica dados los alcances de los análisis presentados.
Guerrero se caracteriza por ser uno de los estados más biodiversos en México. Sin embargo, los cambios de uso de suelo y vegetación han resultado en una pérdida significativa del 32% de hábitats naturales originales, respecto al 28.2% a nivel nacional. Asumiendo que la pérdida del hábitat natural en la distribución de las especies es una de las causas documentadas con mayor efecto negativo en su supervivencia (Best et al., 2001; Challenger y Dirzo, 2009; Fahrig, 2003; Kinnaird et al., 2003; Mendoza et al., 2005; Steffan-Dewenter et al., 2002; Venier y Fahrig, 1996), es importante notar algunos patrones espaciales en la distribución de las especies, sobre todo, aquellas consideradas como vulnerables. A la escala de Guerrero, mientras especies como Cryptotis goldmani –cuya distribución de sus hábitats adecuados potenciales está restringida a <1% de la superficie del estado– se estima han perdido apenas el 5% de la cobertura vegetal original, otras especies con distribuciones potenciales igualmente restringidas en el estado, como Pipistrellus hesperus, se estima han perdido hasta del 41% de la vegetación original de sus hábitats adecuados potenciales (tabla 1). Esto pone en evidencia que existen diferencias notables en cómo son afectados, de distinta manera, los hábitats adecuados potenciales, aún para el caso de especies con distribución restringida, a una porción pequeña a escala estatal. Esta tendencia y la falta de correlación significativa observada tanto a nivel nacional como a nivel estatal, entre la amplitud de la distribución de hábitats adecuados potenciales y el porcentaje de estos que aún conservan su vegetación natural original, sugiere que es indispensable considerar el nivel de afectación de especie por especie y la escala en la que se pretenden realizar acciones de conservación.
Tomando en consideración estimados globales obtenidos de todas las especies, tal como la mediana, es evidente que el porcentaje de hábitats adecuados actuales (naturales) en Guerrero (83%), es mucho mayor a ese mismo porcentaje a nivel nacional (61%). De hecho, se encontraron diferencias significativas al analizar la proporción de hábitats naturales que conservan las especies a nivel estatal, en relación con lo que sucede a nivel nacional. Esto podría sugerir escenarios favorables para la conservación de las especies de mamíferos terrestres en Guerrero.
Por otro lado, dentro de los criterios de inclusión de especies en la Norma Oficial Ecológica Mexicana, usualmente se considera a la amplitud de la distribución de la especie como un factor clave y relevante (Feria, Olson, García-Mendoza y Solano, 2009; Sánchez et al., 2007); las especies de distribución restringida son presumiblemente más vulnerables a impactos de pérdida de hábitat, tales como extirpaciones poblacionales o extinciones locales, en relación con especies que muestran distribuciones amplias (Sánchez-Cordero et al., 2009; Sánchez- Cordero, Illoldi-Rangel et al., 2005; Sánchez-Cordero, Munguía et al., 2005). En el presente estudio, las 6 especies incluidas en alguna de las categorías de riesgo de la Norma Oficial Ecológica Mexicana (Nom-059-Semarnat-2010 [Semarnat, 2010]) se encuentran entre las que mostraron un porcentaje significativo de sus hábitats adecuados actuales, respecto a la distribución de sus hábitats adecuados potenciales (>80%; ver tabla 2). De acuerdo con nuestros resultados, este grupo de especies podrían mostrar un estado de conservación mucho más favorable en Guerrero, que a nivel nacional, en donde los rangos se encuentran entre el 56 y 85%, lo cual podría explicarse por la proporción elevada de hábitats originales que se han perdido en otros estados, como Veracruz y Tabasco. Por tanto, es necesario impulsar, a corto plazo, acciones en Guerrero que promuevan las condiciones de conservación actuales.
Los resultados de este estudio nos permiten confirmar que la escala de análisis puede marcar tendencias del estado de conservación de una especie en ambos sentidos; una especie puede mostrar una categoría de amenaza o riesgo de extinción local, sin mostrar un estado de conservación desfavorable a nivel nacional y viceversa (Sánchez-Cordero et al., 2009). Dentro del proceso de planificación de conservación local y regional, o al medir el impacto de actividades productivas, mediante estudios de impacto ambiental, se hace necesario utilizar algún índice de riesgo específico a nivel de especie, que considere el estado de conservación estatal o regional, además de observar la categoría de riesgo según la Norma Oficial Ecológica Mexicana. Feria et al. (2009) resaltan el uso de los sistemas de información geográfica para establecer criterios de riesgo de conservación de especies. Este trabajo pone de relieve la necesidad de considerar análisis a escalas múltiples: nacional, regional, estatal, local; para definir el estado de conservación de una especie (Margules y Sarkar, 2009; Sánchez-Cordero et al., 2009).
Este enfoque teórico-metodológico deberá considerar requerimientos más específicos de hábitat que los considerados en este trabajo. Por ejemplo, se estima que Spermophilus adocetus ha perdido alrededor del 21% de hábitat adecuado en su distribución potencial para Guerrero y, aproximadamente, el 37% para México. Aun cuando esta es una especie endémica y, por tanto, de distribución restringida, podría no verse drásticamente afectada por los cambios de uso de suelo, puesto que se ha descrito como una especie que puede incrementar sus poblaciones ante el avance de la frontera agrícola (Sánchez-Cordero, Illoldi-Rangel et al., 2005). En contraste, Spilogale pygmaea, también endémica e incluida en la Norma Oficial Mexicana como especie “amenazada” y como especie “vulnerable” dentro de la Lista Roja de la International Union for Conservation of Nature (UICN) de Especies Amenazadas (IUCN, 2013), ha perdido 18 y 28% de la cubierta vegetal original dentro de su distribución potencial en Guerrero y el país, respectivamente. Sin embargo, aun siendo capaz de invadir zonas agrícolas y pastizales, esta especie depende de su hábitat natural en buen estado para mantener poblaciones a largo plazo (Cuarón y Helgen, 2008). Por otro lado, Choeronycteris mexicana, incluida como especie “amenazada” en la Norma Oficial Mexicana y como “casi amenazada” por la Lista Roja de la IUCN (IUCN, 2013), ha perdido aparentemente solo el 16% de la cubierta vegetal en sus hábitats adecuados potenciales en el estado, pero depende de un nivel alto de calidad de hábitat (primario) y se estima que sus poblaciones han disminuido un 30% a nivel nacional (Arroyo-Cabrales y Pérez, 2008).Respecto a las áreas de conservación seleccionadas, en contraste con el análisis de conservación realizado recientemente en el estado que se concentra en una sola especie (Cuervo-Robayo y Monroy-Vilchis, 2012), el ejercicio aquí realizado para identificar soluciones de conservación para distintas especies, coinciden con sitios anteriormente identificados como prioritarios para la conservación de vertebrados en Guerrero (Álvarez-Mondragón y Morrone, 2004; García-Marmolejo, Escalante y Morrone, 2008; García-Trejo y Navarro, 2004; Ruiz-Gutiérrez, 2012). Es interesante notar que la cantidad de área seleccionada coincide bajo los 2 escenarios en los que se realizó el análisis (hábitats adecuados potenciales versus hábitats adecuados actuales) y que no existieron diferencias significativas entre las áreas seleccionadas a nivel municipal. Fuller et al. (2006) demostraron que en el caso de las especies endémicas de mamíferos de México, el costo de conservación se incrementa conforme el proceso de pérdida de hábitat natural avanza. Es más complicado alcanzar metas realistas de conservación de biodiversidad, a partir de una matriz de hábitat natural conservado, que a partir de un hábitat natural fragmentado. Sin embargo, para el grupo de especies de mamíferos incluidas, aparentemente la pérdida de hábitat aún no redunda en mayores requerimientos de áreas para proteger el 20% de la distribución de las especies. Esta situación podría modificarse si se buscara una meta de representación más elevada.
Al analizar el sistema de áreas seleccionadas, resulta preocupante que la solución de conservación entre ambos escenarios fue radicalmente distinta, respecto al número de áreas seleccionadas (135 para hábitats adecuados potenciales versus 1,212 para hábitats adecuados actuales) y, también, en cuanto al perímetro (5,445 para hábitats adecuados potenciales versus 15,599 para hábitats adecuados actuales) factores que, debido a su repercusión crítica en la conservación de la biodiversidad (Fahrig, 2003), requieren ser investigados de manera específica, mediante trabajos enfocados a realizar propuestas de conservación.
Considerando la ausencia de áreas de conservación en el estado de Guerrero, identificamos como prioridad, a corto plazo, el inicio de estrategias significativas de conservación de la biodiversidad estatal. Se recomienda la aplicación de los siguientes enfoques: 1) expandir nuestro análisis a otros grupos taxonómicos, para establecer una plataforma de información indispensable que sirva para elaborar propuestas integrales de conservación; 2) incluir metas variables de conservación de acuerdo con modelos refinados de la distribución actual de las especies y estimados de sus niveles de vulnerabilidad intrínseca y 3) buscar un objetivo de conservación multicriterio que promueva mayor conectividad entre las zonas seleccionadas en el estado.
F. Botello agradece el apoyo del Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México y del Consejo Nacional de Ciencia y Tecnología (Conacyt; CVU 48454). Los autores agradecen a la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad por proporcionar los puntos de colecta (oficio: DTAP/096/2012). Asimismo, agradecen las sugerencias de 2 revisores anónimos que ayudaron a aclarar conceptos y a mejorar significativamente la presentación del manuscrito.