


During an inventory of the helminths of amphibians in Mexico, specimens of an undescribed species of Haematoloechus were collected from the lungs of Rana zweifeli and Rana sp. in southwestern Mexico. The objective of this study is to describe this species. The new species differs from other known species of Haematoloechus by the combination of the following characters: spindle shaped body, oral sucker/acetabulum ratio 1:0.31, longitudinal extracecal uterine loops extending to the level of posterior testis, ovary irregular, lobed and testes elongate, lobed.
Durante el desarrollo del inventario de helmintos de anfibios en México se recolectaron ejemplares de una especie aún no descrita del género Haematoloechus de los pulmones de Rana zweifeli y Rana sp. en el suroeste de México. El objetivo de este trabajo es describir esta especie. La especie nueva difiere de las conocidas de Haematoloechus por la combinación de los siguientes caracteres: cuerpo en forma de huso, relación ventosa oral/acetábulo 1:0.31, asas uterinas longitudinales extracecales extendiéndose hasta el nivel del testículo posterior, ovario irregular, lobulado y testículos alargados, lobulados.
Haematoloechus spp., is a species-rich genus with more than 50 described species around the globe. These digeneans are common parasites occurring in the lungs of frogs, and are rarely found in other groups of amphibians (toads or salamanders). Morphological variability within species of this genus has caused controversy regarding species validity (Kennedy, 1981; Odening, 1960; Prokopic & Krivanek, 1974), and molecular characters have demonstrated to be useful in the differentiation of species in this genus, validating the use of some morphological characters (León-Règagnon, Brooks, & Pérez-Ponce de León, 1999; León-Règagnon, Brooks, & Zelmer, 2001; León-Règagnon & Paredes-Calderón, 2002). Particularly, DNA barcoding (Hebert, Cywinska, Ball, & deWaard, 2003) has proven to be a useful tool in the prospection of cryptic species of Haematoloechus.León-Règagnon (2010) used partial COI sequences to differentiate species of Haematoloechus in Mexico including several specimens that according to morphology did not correspond to any of the known described species of the genus. The sequence of one of this specimens (GenBank HQ141707) appeared a as an independent lineage in the resulting trees, with a minimum genetic divergence of 9.9% with the other sequences, contrary to a maximum of 2.2% among sequences of the same species, representing a potential undescribed species.
The aim of this study is to morphologically describe and name this species that parasites R. zweifeli Hillis, Frost and Webb, 1984 and Rana sp. in southwest Mexico, and that was detected using DNA barcodes by León-Règagnon (2010).
Materials and methodsTwenty two specimens of Rana zweifeli were collected in Santiago Tamazola, Oaxaca (17°39′35.2′ N, 98°13′21.08′ W) in 2000, 21 in Amojileca, Guerrero (17°34′11.50′ N, 99°34′12.77′ W) in 2001, and 9 specimens of Rana sp. were collected in San Fernando, Chiapas (16°40′50.85′ N, 93°13′06.69′ W) in 1999. All specimens were collected under the scientific collection permit FAUT0056 issued to VLR, and representative specimens of the hosts were deposited at Museo de Zoología de la Facultad de Ciencias, UNAM (MZFC 14634, 14642, 14644, 14789-14792, 14887-14893, 12900).
Frogs were captured manually or using dip nets and killed by an overdose of sodium pentobarbital. Helminthological examination was done in the following 24hours from capture. Worms collected from freshly killed amphibians were initially placed in saline (0.65%). Some specimens were fixed in 95% ethanol for molecular study; for morphological study they were fixed by sudden immersion in hot 4% formaldehyde and then preserved in 70% ethanol. Specimens were stained with Mayer's paracarmine or Gomori's trichrome, dehydrated, cleared in methyl salicylate, and mounted permanent slides using Canada balsam. Some specimens were permanently mounted in Cobb slides. Measurements are presented as the range with means in parentheses and expressed in micrometers, unless otherwise is stated. Figures were drawn with the aid of a drawing tube. The following abbreviations are used: CNHE, Colección Nacional de Helmintos, Instituto de Biología, Universidad Nacional Autónoma de Mexico.
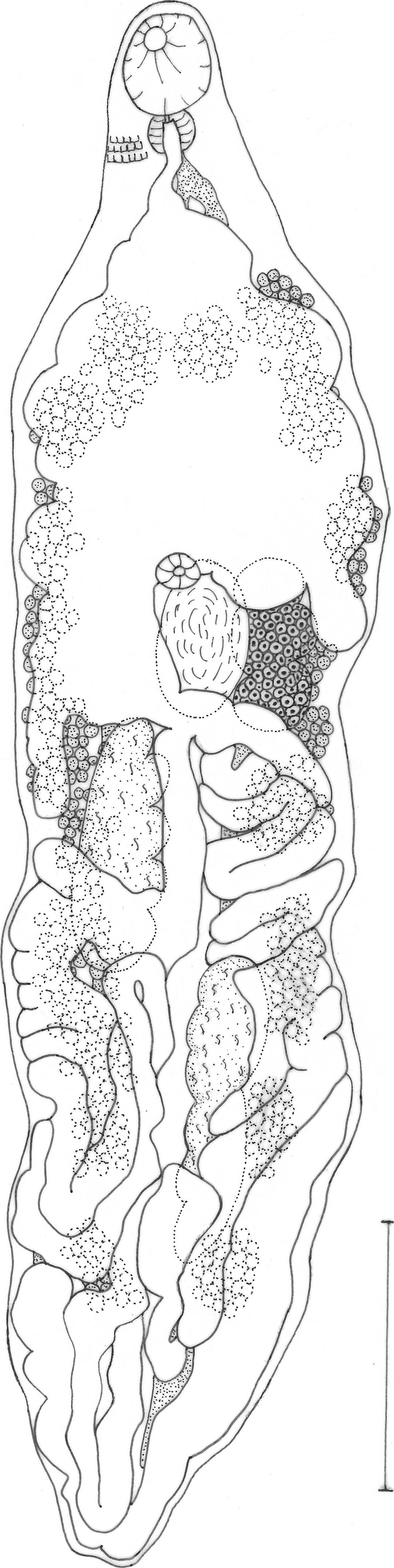
DescriptionHaematoloechus nicolasi n. sp. (Fig. 1)Description based on 12 mature specimens: body spindle shaped, with pointed anterior and posterior ends; 2.52–6.04 (3.89) mm long, 0.84–1.97 (1.25) mm of maximum width at the preovaric region. Tegument thick, no spines, showing plications forming a grid. Oral sucker terminal, round, 271.6–417.1 (348.39) long, 217.6–407.4 (339.5) wide; opening directed ventrally. Pharynx oval, 126.1–242.5 (183.42) long, 106.7–213.4 (168.43) wide; oral sucker/pharynx ratio 1:0.39–0.62 (0.52). Gland cells surrounding anterior region of pharynx. Esophagus 48.5–126.1 (76.21) long, surrounded anteriorly by gland cells, sometimes obscured by uterus. Ceca bifurcating at 465.6–649.9 (559.95) [10.6–16.8 (14.28)% of body length (BL)] from anterior extremity. Ceca narrow, terminating blindly near posterior extremity. Ventral sucker weakly developed, sometimes obscured by ascending uterus, 97–135.8 (106.7) long, 87.3–135.8 (109.93) wide, at 1.32–2.04 (1.66) mm [33.7–47.2 (41.33)% of BL] from anterior extremity. Suckers length ratio 1:0.23–0.36 (0.31). Testes 2, elongate slightly lobed, oblique, immediately posterior to ovary. Anterior testis opposite to ovary, 388–873 (583.94) long, 194–339.5 (258.02) wide. Posterior testis 582–1115.5 (725.56) long, 252.2–417.1 (308.63) wide. Cirrus sac obscured by ascending uterus. Ovary irregular, lobed, 339.5–679 (521.64) long, 242.5–436.5 (312.56) wide; at 1.2–2.39 (1.83) mm [40.6–56.81 (47.94)% of BL] from anterior extremity. Seminal receptacle adjacent partially overlapping ovary; 291–533.5 (430.68) long, 194–407.4 (291.97) wide. Mehlis gland dorsal to seminal receptacle. Laurer's canal not observed. Vitellaria forming clusters overlapping with each other, distributed sidelong, dorsally invading space between caeca in their anterior limit and in the postesticular region. Anterior limit of vitellaria 242.5–485 (419.04) [7.43–11.56 (10.91)% of BL] from anterior end; posterior limit at level of posterior testis on the ovarian side of body, at end of caeca on the opposite side of ovary. Descending uterus forming transversal and diagonal loops on ovarian side of body, partially overlapping testis, filling intra and extracecal space toward posterior end of body. Ascending uterus forming longitudinal loops oriented anteriorly reaching the level of posterior testis, and continuing anteriorly forming transversal and short longitudinal loops at the level of testes, partially overlapping anterior testis and seminal receptacle, and filling with transversal loops the entire preovarian region. Genital pore ventral to posterior border of oral sucker. Eggs dark brown, 16.95–26.63 (21.11) long, 9.68–16.95 (13.59) wide. Excretory vesicle not observed. Excretory pore terminal.
Taxonomic summary
Host: Rana zweifeli.
Site of infection: lungs.
Holotype: CNHE 10039.
Paratypes: CNHE 10040, 10041, 10043.
Locality: Santiago Tamazola, Oaxaca, Mexico.
Etymology: species name dedicated to my son, Nicolás Flores, who is my savior and motivation.
Other hosts and localities: R. zweifeli, Amojileca, Guerrero; Rana sp. San Fernando, Chiapas.
Other records: R. zweifeli, Santiago Tamazola, Oaxaca (León-Règagnon, 2010) based on COI sequences, referred as Haematoloechus sp. Ha65.
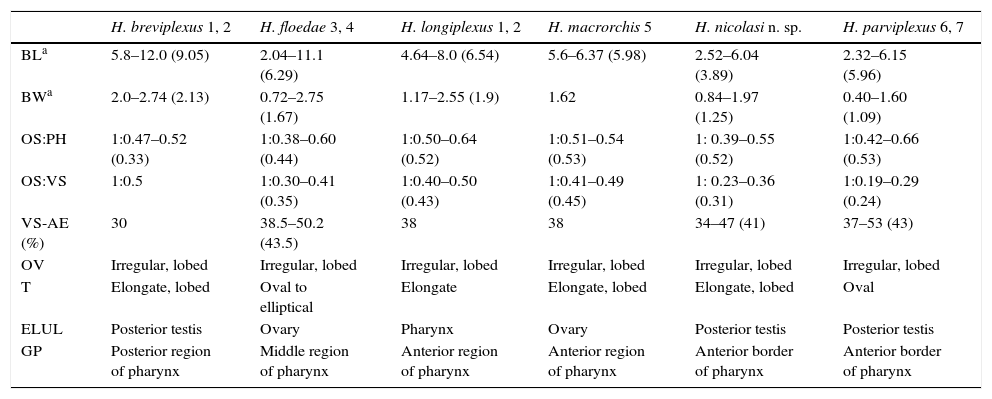
RemarksThe new species differs from other American species of Haematoloechus and resembles H. breviplexusStafford, 1902, H. floedaeHarwood, 1932, H. longiplexusStafford, 1902, H. macrorchisCaballero, 1941 and H. parviplexusIrwin, 1929 in having an acetabulum less than half of the size of the oral sucker and longitudinal extracecal uterine loops (Table 1).
Morphological comparison between American Haematoloechus species with acetabulum less than half of the size of the oral sucker and with extracecal longitudinal uterine loops.
| H. breviplexus 1, 2 | H. floedae 3, 4 | H. longiplexus 1, 2 | H. macrorchis 5 | H. nicolasi n. sp. | H. parviplexus 6, 7 | |
|---|---|---|---|---|---|---|
| BLa | 5.8–12.0 (9.05) | 2.04–11.1 (6.29) | 4.64–8.0 (6.54) | 5.6–6.37 (5.98) | 2.52–6.04 (3.89) | 2.32–6.15 (5.96) |
| BWa | 2.0–2.74 (2.13) | 0.72–2.75 (1.67) | 1.17–2.55 (1.9) | 1.62 | 0.84–1.97 (1.25) | 0.40–1.60 (1.09) |
| OS:PH | 1:0.47–0.52 (0.33) | 1:0.38–0.60 (0.44) | 1:0.50–0.64 (0.52) | 1:0.51–0.54 (0.53) | 1: 0.39–0.55 (0.52) | 1:0.42–0.66 (0.53) |
| OS:VS | 1:0.5 | 1:0.30–0.41 (0.35) | 1:0.40–0.50 (0.43) | 1:0.41–0.49 (0.45) | 1: 0.23–0.36 (0.31) | 1:0.19–0.29 (0.24) |
| VS-AE (%) | 30 | 38.5–50.2 (43.5) | 38 | 38 | 34–47 (41) | 37–53 (43) |
| OV | Irregular, lobed | Irregular, lobed | Irregular, lobed | Irregular, lobed | Irregular, lobed | Irregular, lobed |
| T | Elongate, lobed | Oval to elliptical | Elongate | Elongate, lobed | Elongate, lobed | Oval |
| ELUL | Posterior testis | Ovary | Pharynx | Ovary | Posterior testis | Posterior testis |
| GP | Posterior region of pharynx | Middle region of pharynx | Anterior region of pharynx | Anterior region of pharynx | Anterior border of pharynx | Anterior border of pharynx |
Ranges followed by average in parentheses; measurements obtained from original descriptions and redescriptions 1=Stafford, 1902; 2=Cort, 1915; 3=Harwood, 1932; 4=León-Règagnon et al., 2005; 5=Caballero, 1941; 6=Irwin, 1929; 7=Bolek and Janovy, 2007.
The new species differs from all known species of the genus by the combination of several characters that have been proven to be useful to differentiate species of Haematoloechus (Bolek & Janovy, 2007; León-Règagnon & Brooks, 2003; León-Règagnon, Brooks, & Pérez-Ponce de León, 1999; Zamparo, Ferrao, Brooks, Bettaso, & Mata-López, 2011), as follows.
It differs from other species in the shape of the body, which is spindle shaped, with pointed anterior and posterior ends; posterior end can be invaginated. Maximum width of body in the preovarian region. When worms are alive, it is notorious the manner in which they extended both extremities of the body while keeping the middle part unchanged in width. Specimens of other species extend or contract the entire body when alive. Body shape can vary depending on the fixation technique, and other species of the genus may show a similar shape in mounted specimens, this is why this character is useful to differentiate the new species only in conjunction with the other diagnostic characters.
It differs from H. floedae, H. longiplexus, and H. macrorchis in the extension of the extracecal longitudinal uterine loops, which reach the level of the ovary in H. floedae and H. macrorchis, the level of the pharynx in H. longiplexus and only the level of the posterior testis in the new species. It also differs from H. floedae in the shape of the testes, being oval or elliptical in that species and elongate and lobed in H. nicolasi, and in the position of the genital pore, which is at the medium level of the pharynx in H. floedae and in the anterior border of the pharynx in H. nicolasi (Harwood, 1932; León-Règagnon, Guillén-Hernández, & Arizmendi-Espinosa, 2005).
The new species also differs from H. longiplexus and H. macrorchis in the size of the acetabulum compared to the oral sucker, which is larger in those species [1:0.40–0.50 (0.43) in H. longiplexus, 1:0.41–0.49 (0.45) in H. macrorchis and 1:0.23–0.36 (0.31) in H. nicolasi] (Cort, 1915; Caballero, 1941; Stafford, 1902).
Haematoloechus nicolasi resembles H. breviplexus and H. parviplexus in the extent of the extracecal longitudinal uterine loops, which in the 3 species reach the level of the posterior testis. It differs from H. breviplexus in the size of the acetabulum compared with the oral sucker, which is larger in that species [1:0.5 vs. 1:0.23–0.36 (0.31)]; also differs in the position of the genital pore, being ventral to the posterior region of the pharynx in H. breviplexus (Cort, 1915; Stafford, 1902) and at the level of the anterior border of this organ in H. nicolasi.
The new species also differs from H. parviplexus in the size of the acetabulum compared with the oral sucker, which is smaller in that species [1:0.19–0.29 (0.24) vs. 1:0.23–0.36 (0.31)] and in the shape of the testes, oval in H. parviplexus and elongate, lobed in the new species.
Partial COI sequence of this species was published by León-Règagnon (2010). In that study, the author mentions that she was not able to differentiate this sample from others collected from the same host and locality using morphology. Nevertheless, COI sequences served to separate those samples in independent lineages, one corresponding to the new species described herein and other corresponding to H. longicollumLeón-Règagnon and Romero-Mayén (2017). These 2 species differ in the size of the acetabulum, which is less than half of the size of the oral sucker in H. nicolasi, and in the presence of longitudinal uterine loops, which are absent in H. longicollum. When the worms are alive, specimens of both species present an elongated anterior part of the body and it is difficult to observe the size of the acetabulum or the arrangement of the uterine loops. The shape of the posterior part of the body is different, but not enough to differentiate them by itself. It was only after measuring mounted specimens and at the light of the molecular evidence that both species were clearly diagnosed.
DiscussionDifferentiation and validation of species of Haematoloechus has been controversial because of the intraspecific variability of species (Kennedy, 1980, 1981), and also because of the lack of details in the original descriptions of many species (Seely, 1906; Stafford, 1902). Molecular markers have served to identify those characters that are reliable in the differentiation of species in the genus, some of which are the oral sucker/pharynx ratio, oral/ventral suckers ratio, the arrangement of the uterine loops in mature worms (presence of extracecal loops, presence and extension of longitudinal loops), the shape of ovary and testes and body shape (León-Règagnon & Brooks, 2003; León-Règagnon, Brooks, & Pérez-Ponce de León, 1999; León-Règagnon, Brooks, & Zelmer, 2001; León-Règagnon & Paredes-Calderón, 2002; León-Règagnon & Romero-Mayén, 2017; Zamparo et al., 2011). Although all of them show some intraspecific variation, the combination of several of them can be used to distinguish species of this genus with confidence; in the case of H. nicolasi n. sp., the combination of body shape, oral sucker/acetabulum ratio, the extension of extracecal longitudinal uterine loops and the shape of ovary and testes, define this new species.
León-Règagnon (2010) using DNA barcodes (COI sequences), detected several undescribed lineages of Haematoloechus, one of which is H. nicolasi n. sp. In that study, sequences of H. nicolasi n. sp. appear as the sister taxon of H. illimisCaballero, 1942. Morphologically, these 2 species differ in the size of the acetabulum, which is 83% the size of the oral sucker in H. illimis (vs. 31% in H. nicolasi); also differ in the extension of the extracecal longitudinal uterine loops, which are absent in H. illimis (Caballero, 1942), and reach the level of the posterior testis in H. nicolasi. Apparently, the size of suckers and the extension of extracecal longitudinal uterine loops, although very useful to differentiate species, are characters with no phylogenetic value, but further investigation on the evolution of these characters is needed in a phylogenetic framework involving more than one molecular marker.
Haematoloechus nicolasi is the second species of the genus to be described as parasite of frogs of the tarahumarae group sensuHillis and Wilcox (2005); the first was H. longicollum, from R. psilonota and R. zweifeli (León-Règagnon & Romero-Mayén, 2017). Both species were found infecting the same host species in the same localities. Other groups of frogs have been sampled in Mexico, but H. nicolasi was found preferably in R. zweifeli, and only in one specimen of Rana sp., which belongs to an undescribed species of leopard frogs (Pérez-Ramos, com. pers.). According to the evidence obtained with mitochondrial sequences (León-Règagnon, 2010), H. nicolasi and H. longicollum (GenBank HQ141704, HQ141706) are not each other closest relatives, suggesting 2 independent colonization events to the tarahumarae group of frogs. Further investigation on the phylogeny of Haematoloechus is needed in order to clarify its coevolutionary history.
I am grateful to María Antonieta Arizmendi-Espinosa, Elisa Cabrera-Guzmán, Elizabeth Martínez-Salazar, Rosario Mata-López, Edmundo Pérez-Ramos, Ulises Razo-Mendívil and Alejandro Zaldívar-Riverón, for their help in the collection of specimens and Edmundo Pérez-Ramos for the identification of hosts. This study was partially funded by PAPIIT-UNAM Proj. IN203911 and IN209414, Conacyt Proj. 54475 and 220408, and PASPA-UNAM 2016 to VLR.
Peer Review under the responsibility of Universidad Nacional Autónoma de México.