



Variations in the quality or quantity of a food source determines if an animal takes the risk of spending energy searching for or eating it. Hummingbirds have been traditionally catalogued as risk-averse foragers. However, the inclusion of more than 2 options in a foraging set for risk-sensitive experiments has resulted in the observation that some hummingbird species preferred intermediate risk even if they had been risk-averse in a traditional binary risk experiment. These contrasting results suggest an effect of having multiple foraging options that had been ignored due to the design of previous risk experiments. Here, we studied the influence of varying reward volume (Experiment 1) or sugar concentration (Experiment 2) on choice behavior of white-eared hummingbirds, Hylocharis leucotis, by recording their visits to feed from a sucrose solution located in 4 artificial floral arrays associated with constant, low, medium, and high variance. In both experiments, each of the vertical arrays was evaluated in a training stage and a test stage. The birds visited all the arrays without discriminating among them, and thus were indifferent to variations in the volume or sugar concentration of reward. Thus, there was no influence of variance of nectar volume and sugar concentration on the choice behavior of the birds, ruling out the possibility that white-eared hummingbirds are risk sensitive under these conditions of 4 foraging alternatives.
La variación en la cantidad o calidad de una fuente de alimento determina si un animal toma el riesgo de invertir energía en su búsqueda o consumo. Los colibríes han sido típicamente catalogados como forrajeadores aversivos al riesgo. Sin embargo, la inclusión de más de 2 opciones en una prueba de forrajeo para experimentos de sensibilidad al riesgo han resultado en la observación de que algunas especies de colibríes muestran riesgo intermedio incluso si habían sido aversivos al riesgo en pruebas binarias de riesgo. Estos resultados contrastantes sugieren un efecto de enfrentar opciones múltiples de forrajeo, el cual había sido ignorado debido al diseño de experimentos previos sobre riesgo. Aquí, se estudia la influencia de la variación en el volumen (Experimento 1) o la concentración de azúcar (Experimento 2) en la conducta de elección de colibríes oreji-blanca Hylocharis leucotis, al registrar sus visitas a 4 arreglos florales artificiales asociados con una varianza constante, baja, media y alta. En ambos experimentos, cada arreglo vertical fue evaluado a través de una fase de entrenamiento y una de prueba. Las aves visitaron todos los arreglos sin discriminar entre ellos, siendo indiferentes a la variación en el volumen o concentración de la recompensa ofrecida. Así, no hubo un efecto de la varianza en el volumen de néctar y la concentración de azúcar en la conducta de elección de estas aves, eliminando la posibilidad de que los colibríes oreji-blanca son sensibles al riesgo bajo condiciones de alternativas múltiples de forrajeo.
In hummingbird-pollinated plants, the variation in nectar volume and sugar concentration is affected by several factors including phylogeny and environmental conditions (Ornelas et al., 2007). However, nectar volume is mostly affected by floral traits (e.g., corolla size, age of the flower) and the physiology of the plant (flower-water relations), while the nectar composition is mainly affected by both the concentration and the composition of nutritional elements (mostly sugars) (De la Barrera and Nobel, 2004). The wide variation in these 2 variables means that nectar is a naturally risky resource (Lara, 2008). Thus, one might predict that hummingbirds should react to variability in the amount of energy obtained or the spatial distribution of nectar rewards (Pyke, 1978; Real and Caraco, 1986). In this risk sensitivity context, hummingbirds and invertebrate nectar foragers, such as honey bees and bumblebees, can show 3 behaviors when confronted with variance: (1) if a forager prefers a constant reward, then its preference is called risk-averse, (2) if a forager prefers the gamble then its preference is called risk-prone. However, (3) if an animal ignores the variance, then it is called risk-indifferent (Stephens and Krebs, 1986; Bateson and Kacelnik, 1998).
Most data on risk-sensitive foraging have been obtained from experiments in which the forager is presented with a binary choice of 2 foraging options differing in the variance in rate of reward (e.g., Stephens and Paton, 1986; Caraco et al., 1990; Reboreda and Kacelnik, 1991, Bateson and Kacelnik, 1996). However, an increasing body of experimental literature indicates that the direction of risk-sensitive preferences can depend on the context in which the foraging options are presented (Hurly and Oseen, 1999; Bateson, 2002; Hurly, 2003). Thus, the importance of variability to foraging preferences would be more convincing if experimental animals were able to choose between more than 2 foraging options.
Hummingbirds (specifically North American Selasphorus species) have been traditionally catalogued as risk-averse foragers (Stephens and Paton, 1986; Waser and McRobert, 1998). However, this behavioral response can be affected by energy thresholds such as starvation and reproduction (Hurly, 2003), prior experience of the individual (Bacon et al., 2010), and number of foraging options per se (Hurly and Oseen, 1999). Thus, the inclusion of more than 2 options in the foraging set has resulted in some hummingbird species preferring intermediate risk even when they were risk-averse in a traditional binary risk experiment (Hurly and Oseen, 1999; Bateson, 202; Hurly, 2003). For example, Rufous hummingbirds (Selasphorus rufus) were risk-averse when presented with the binary choice between flowers with constant nectar volumes versus flowers with variable nectar volumes. Binary comparisons were conducted both between a constant flower type and a moderately variable flower type, and one between a constant flower and highly variable flower type. When all 3 options were presented simultaneously, hummingbirds preferred the moderately variable flower to both the constant and highly variable flower options (Hurly and Oseen, 1999). Thus, hummingbird's behavior could be explained if we consider that discrimination of alternatives depends on the relative difference between the alternatives (Waddington and Gottlieb, 1990), as well as the number of these alternatives.
Here we present the results of a risk-sensitive foraging experiment with white-eared hummingbirds (Hylocharis leucotis). Given the potential importance of the studies showing contrasting results that depend on the foraging options offered in risk-sensitive experiments, our aim in this study was to replicate and extend these experiments in a Neotropical hummingbird species. We tested hummingbirds’ behavioral response for variance in nectar volume and sugar concentration in 4 possible combinations of constant, low variance, intermediate variance, and high variance.
Materials and methodsStudy site and species. The subjects were forty adult white-eared hummingbirds (Hylocharis leucotis) resident in La Malinche National Park, Tlaxcala, México (19°14' N, 98°58' W, 3 000m asl). All subjects were studied after receiving ethical approval from the relevant local authorities. From November 2006 to July 2008, the hummingbirds were mist-netted in the field for this study. All trials described below were conducted from 08:00-15:00h, and birds were housed individually in collapsible field cages (dimensions: 51x51x51cm), containing a perch. Cages were hung in the shade where the birds were captured, with ambient temperature and light. Prior to the experiments, naive individuals were allowed to acclimate to the cages for 10min and fed with an artificial flower (identical to the one used in the experiments) containing 5μl of 20% (by mass) sucrose solution. During this period, perching by the birds was taken as evidence that they were acclimatized to the enclosure. Non-perching individuals were released and not taken into account in the experiments.
For the experiments, we made artificial flower arrangements using conical plastic micropipette tips of 45mm length (corolla length). Because this hummingbird species uses mostly red flowers throughout the year in La Malinche, we decided to use red artificial flowers, having simulated “petals made of red plastic material”, to ensure that subjects choose flowers based on nectar reward rather than on inherent color preference. The length of corollas was within the range of hummingbird-pollinated flowers normally encountered at La Malinche (Lara, 2006).
Each of the 4 vertical floral arrays used in both experiments consisted of 3 artificial flowers spaced 3cm apart. These flowers were mounted on 30cm wooden stakes. Half of the individuals captured in the study were evaluated in arrangements where the variance in the volume was manipulated (Expt 1), and the other half were evaluated where the variance in sugar concentration was manipulated (Expt 2).
Experiment 1: volume variation. Experiments attempting to test risk-sensitivity in animals have 2 stages: a training stage and a testing stage (Kacelnik and Bateson, 1996). During the training stage, animals learn to associate the position of the flowers in the array, with its value. Once the association is learned, we can “ask” animals which resources they prefer. To assess risk-sensitivity to nectar volume, the experiment involved 20 captured hummingbirds, 12 male and 8 females (body mass ranging from 3.9-5g), that were individually tested and confronted simultaneously with 4 foraging arrays differing in the coefficient of variation (CV) of nectar contained in their flowers. First, each hummingbird was tested in a 10 minute training stage to ensure that it learned the position of the flowers on the arrays, or that a certain level of nectar variation was associated with each array. During this period, the repeated visit to flowers containing the greatest volumes of nectar in the 4 arrays was taken as evidence that birds had learned and perceived these floral arrays as being different. At the end of the first stage, the bird was manually captured and moved to another cage for 5 minutes before initiating the following 10 minute test stage, and the sugar solution was added to the flowers of each array previously used, to the CV values described below. Training periods ranging from 10 to 15 minutes have been previously shown be effective in assuring learning in hummingbirds (Lara et al., 2009; Pérez et al., 2011).
Based on the nectar variation reported for the flora visited by hummingbirds at the study site ranging around a mean of 5μl per flower (Lara, 2006), the CV for each array was calculated as the standard deviation divided by the mean (Shafir, 2000). In the context of foraging, CV measures risk per unit of expected return (Weber et al., 2004). Thus, the sugar solution of our 4 reward arrays contained the same sugar concentration (20% by mass) but differed in volume as follows: 1) constant, 3 flowers containing 5μl of sucrose solution; 2) low, 1 flower containing 8μl, 1 containing 4μl, and 1 containing 3μl of sucrose solution; 3) medium, 3 flowers containing 10μl, 3μl, and 2μl, respectively, and 4) high, 2 empty flowers and 1 containing 15μl of sucrose solution. Thus, the mean volume of nectar for any particular array was 5μl, but CV was either negligible (0) in the constant volume array, low (1.4), medium (3.8) or high (15).
Nectar presence in all flowers from the arrays was corroborated at the end of both stages, indicating that the repeated visits of the birds to a specific flower could not be affected by previous nectar depletion. For both stages, we noted for each individual: 1) the number of visits per flower in each array, and 2) the time between leaving a previous flower and until visiting the present flower (latency). The number of visits per flower (sometimes referred to as the proportion of flowers chosen or cumulative number of flowers visited) and the volume consumed per foraging bout, have been traditionally used as measures of behavioral response in hummingbird risk-sensitivity studies (see Waser et al., 1998; Hurly and Oseen, 1999; Hurly, 2003; Bacon et al., 2010). Due to the impossibility to evaluate the nectar consumed per foraging bout without interrupting our 10 minute stage and possibly affecting the performance of the subjects, we decided to use the number of visits as a measure of behavioral response. Following the exposure to the floral arrays in the experiment, birds were marked by clipping the 5th rectrice feather (to avoid using recaptured birds in subsequent experiments) and then released.
Experiment 2: sugar concentration variation. We tested 20 naive birds (11 males and 9 females) to evaluate if hummingbirds exhibited risk-sensitivity when faced with variation in sugar concentration. The mean sugar concentration calculated in the flowers of the plants visited by hummingbirds in our study site was 20% (Lara, 2006). In order to manipulate the CV of the concentration, the 3 flowers in the 4 arrays contained 5μl of nectar, but they differed only in the CV of its sugar concentration as follows: 1) constant, 3 flowers filled with 20% sucrose solution; 2) low, 1 flower with 29%, 1 with 20%, and 1 filled with 10% sucrose solution; 3) medium, 3 flowers filled with 35%, 15%, and 10%, respectively, and 4) high, 1 empty flower and 2 filled with 30% sucrose solution. Thus, the mean sugar concentration for the array was 20%, but variance was either negligible (0) in the constant volume array, low (4.55), medium (8.75) or high (15).
The identity (in terms of reward volume or sugar concentration) of flowers within arrays was kept constant. Thus, if 1 of 3 flowers in a variable array was empty, this flower was always in the same location (throughout the experiment most individuals stopped visiting empty flowers after 1-2 visits). Also, this was constant within and between subjects. All subjects used in this experiment followed the same registration protocol described in the Experiment 1. For both experiments and during each bout the subjects always visited more than 1 flower type.
Data analysis. For both experiments the subjects sampled flowers on each array to learn the volume and sugar concentration distributions, and their behavioral responses for an array can be tested by the number of visits performed to 1 array in comparison to the other 3. We recorded the cumulative number of visits to each flower by each subject tested. Thus, to compare subjects’ preferences quantitatively, we calculated the slopes of the cumulative choice lines through the 2 phases of both experiments. The comparisons were made with repeated measures analysis of variance (Anova), with both within- subject and between-subject factors. Data normality was previously tested using a Shapiro-Wilk test.
We used Survival Analysis (“time failure analysis”) to analyze hummingbird visitation in both stages in experiments 1 and 2. One of the advantages of using survival analysis is that it allows for use of censored data. Censored data points are those in which an event has not been observed because the study ended before the event could have happened to some individuals under observation. This feature is useful in field biology, where the observation period may be too brief for all possible events to occur (Muenchow, 1986). Thus, we recorded the beginning of our observations as time zero and subsequent foraging events as minutes from start time. If an event occurred for a given flower, then it became uncensored data, and if it never occurred, then it became censored data. We used the Kaplan-Meier product-limit non-parametric method for the computation of the probability that hummingbirds had not yet visited a flower in the arrangement 20min after the start of observation, and the logrank (Mantel-Cox) statistic to test for differences between flower types (Muenchow, 1986). In our case, “survivorship” was the probability that a hummingbird had not yet visited flowers a given number of minutes after the start of the observation period. All statistical analyses were done using StatView (Abacus Concepts, Inc., 1996).
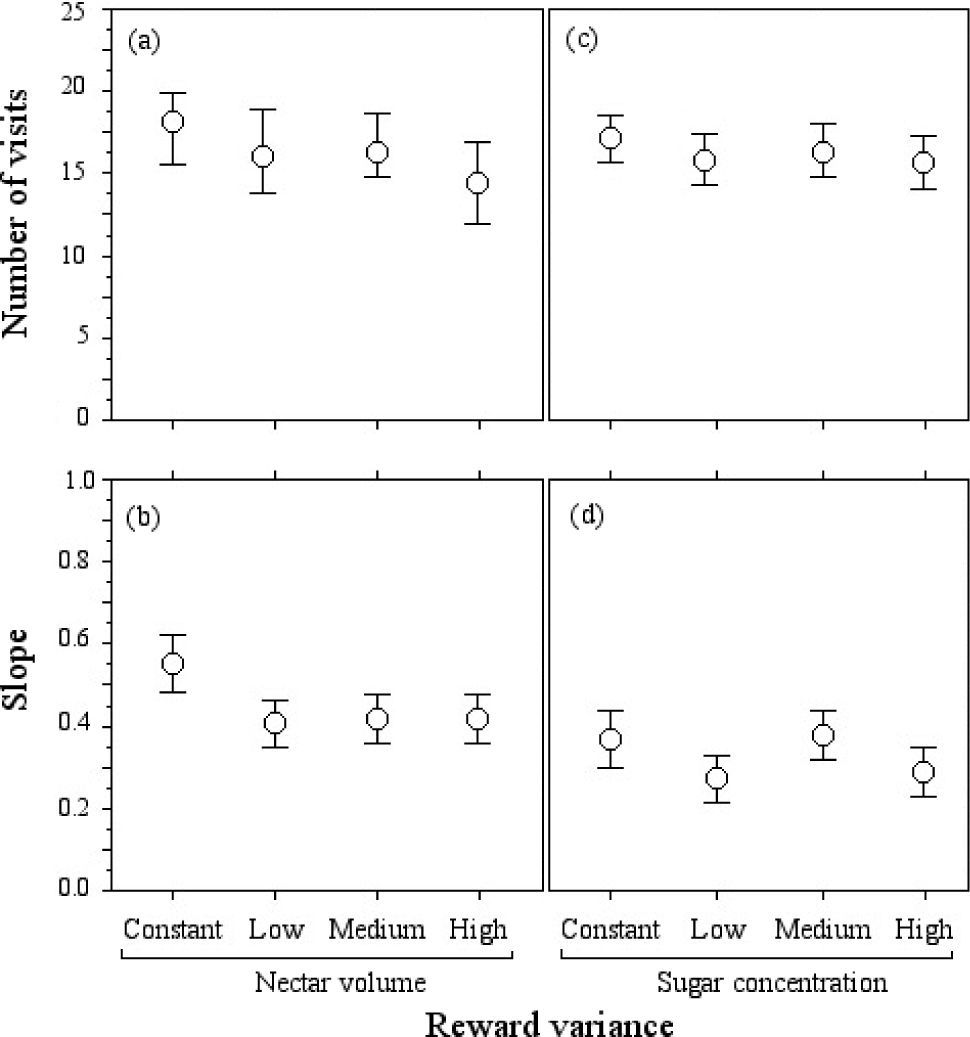
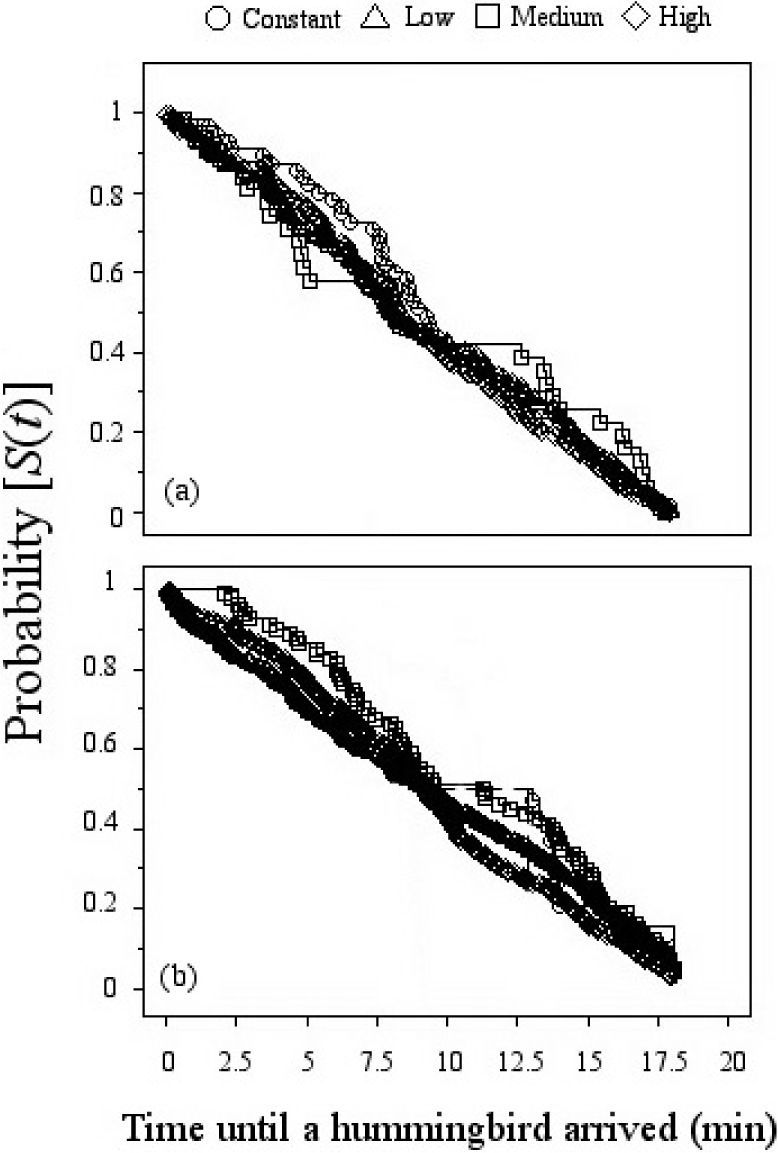
ResultsResponse to volume variation. Examination of choices across all bouts revealed that none of the hummingbirds showed a preference for a specific floral array (Fig. 1a). Our results revealed a non-significant effect of the CV values offered to the hummingbirds on the number of visits to the flowers in the arrays (Table 1a, Fig. 1b). Thus, there was no effect across subjects in the order and extent of preferences, giving further indication that birds showed indifference to volume variation. We found no differences in arrival times to flowers in the 4 arrays (Mantel-Cox Logrank: χ2=0.988, d.f.=3, p=0.804). These results indicate that the flowers of all the arrays had the same probability of being visited throughout the experiment, regardless of the variation of the nectar volume contained in the flowers (Fig. 2a).
 Mean + SE of the number of visits to flowers of each type. (b, d) Mean slope of cumulative visits (n=40 subjects).")
Repeated-measures Anova for hummingbird visits to floral arrays where the coefficient of variation (CV) of (a) nectar volume and (b) sugar concentration was manipulated
| Source | df | SS | F | p |
|---|---|---|---|---|
| (a) Nectar volume between-subjects | ||||
| Treatment | 3 | 0.158 | 0.053 | 0.728 |
| Error | 76 | 9.225 | ||
| Within-subjects | ||||
| Stage | 1 | 0.079 | 1.114 | 0.294 |
| Stage x treatment | 3 | 0.309 | 1.343 | 0.265 |
| Error | 76 | 5.391 | ||
| (b) Sugar concentration between-subjects | ||||
| Treatment | 3 | 0.020 | 0.615 | 0.979 |
| Error | 76 | 8.291 | ||
| Within-subjects | ||||
| Stage | 1 | 0.159 | 2.421 | 0.123 |
| Stage x treatment | 3 | 0.329 | 1.672 | 0.180 |
| Error | 76 | 4.984 |
 nectar volume and (b) sugar concentration, showing probability of being visited by hummingbirds during a 20 minute observation period. The abbreviation S (t) on the y axis is the probability that a hummingbird has not yet visited a flower in an array. “Time until a hummingbird arrived” refers to the time elapsed since the start of the observation period.")
Comparison using survival analysis between artificial flower arrays with different variance in (a) nectar volume and (b) sugar concentration, showing probability of being visited by hummingbirds during a 20 minute observation period. The abbreviation S (t) on the y axis is the probability that a hummingbird has not yet visited a flower in an array. “Time until a hummingbird arrived” refers to the time elapsed since the start of the observation period.
Response to sugar concentration variation. The subjects indifferently visited the flowers in all the arrays (Fig. 1c). There was no preference within or between birds for constant over any variable (low, medium, or high) reward (Table 1b; Fig. 1d). As in experiment 1, the analysis of the probability of visit to the flowers of any array showed that the arrival times were not statistically different (Logrank-Mantel Cox: χ2=1.453, d.f.=3, p=0.061, Fig. 2b).
DiscussionThe results of our study indicate that white-eared hummingbirds are not risk-sensitive foragers under experimental conditions of 4 foraging alternatives. Thus, individual birds make equal numbers of visits to constant, low, medium, and high variance flower arrays. Furthermore, the risk indifference in the birds is the same regardless of whether the nectar's variability is in volume (Experiment 1) or in concentration (Experiment 2). However, the nearly significant difference obtained (p=0.061) between the probability curves of hummingbirds visiting arrays where the concentration was manipulated does not exclude the possibility of risk-sensitivity to this nectar characteristic. Also, because we standardize volume to manipulate sugar concentration, we altered the amount of energy offered by an array and this can have a direct influence on the hummingbird arrival times since drinking less of a more concentrated solution may in fact yield a higher energy reward than drinking more of a lower concentration solution (Brown et al., 2010a, 2010b). Other factors not assessed in this study should be considered in interpreting the results obtained. For example, we do not know the minimal volume a hummingbird needs to ingest to be able to detect the sugar concentration, and therefore it is difficult to determine if a hummingbird was able to differentiate among sugar concentrations when we offer them very small amounts of sugar solutions. Likewise, risk indifference could be caused by hummingbirds being hungry. If they need to obtain energy, and have very small amounts of nectar available, they could be just ingesting all the nectar in the 4 arrays, and this could cause the patterns found here. Further studies should consider all these possibilities.
Our results of consistent risk-indifference are not supported by traditional foraging models such as the variance discounting model (Real, 1980), which predicts constant risk aversion. Nevertheless, other studies have also found tendencies to moderate risk sensitivity in hummingbirds (Waser and McRobert, 1998; Hurly and Oseen, 1999; Hurly, 2003), conventionally interpreted as irrational choice behavior (Bateson et al., 2002). In this sense, the explanation of these findings suggests the necessity to incorporate alternative information into the foraging models such as starvation, fat reserves, use of torpor, and reproductive energetic thresholds (Hurly, 2003).
Here, we have shown that white-eared hummingbirds confronted with 4 foraging options displayed a stochastic choice consistent with no model of risk sensitivity. The risk indifferent results of our experiments agree with other studies of invertebrate pollinators such as honeybees and bumblebees, which also found indifference to variability (Banschbach and Waddington, 1994; Waddington, 1995; Cartar and Abrahams, 1996; Perez and Waddington, 1996; Fulop and Menzel, 2000). Energetic reasons can be invoked to explain these results. Thus, we can suggest that these animals are maximizing their expected short-term rate of net energy gain. The findings in the studies mentioned above suggest that risk indifferent foragers, given the linear relationship between nectar volume/ concentration and rate of gain, may obtain no advantage by preferring either low or high variance flowers under any conditions. Therefore, there can be no selection for sensitivity to variation in nectar rewards based on differences in rate of gain between the flower types, and individuals should visit all foraging options (Perez and Waddington, 1996). Likewise, as suggested by several risk-sensitivity studies (Shafir, 2000;Weber et al., 2004), the apparent discrepancies among studies that report risk-sensitivity and risk-indifference can be due to the lack of control for the magnitude of the CV when testing risk-sensitivity under different conditions. Thus, a consensus is necessary on the ideal magnitude of the CV used in risk-sensitivity studies.
We believe that our reward conditions represent the natural situation present in the “flower market”. If this is true, lack of sensitivity to variance in white-eared hummingbirds is not a failure performance, or limitation of memory processing, but rather an adaptation to cope with the unreliability of scattered food sources such as flowers (Fulop and Menzel, 2000). For example, in our study site both male and female white-eared hummingbirds display a consistent set of foraging strategies, ranging from territorial behaviour on dense flower patches of Penstemon roseus (Lamiaceae) and Bouvardia ternifolia (Rubiaceae) to inconstant foraging events and traplining behaviour in scattered and low-reward flowers such as Castilleja tenuiflora (Scrophulariaceae; Lara, 2006). In both cases, hummingbirds sample multiple patches while foraging on these plant species. Also, it has been shown for these plant species that the variance in rewards (nectar volume and sugar concentration) differs from one patch of flowers to another (Lara, 2006; Lara and Ornelas, 2008). All of these suggest that white-eared hummingbirds naturally experience a great variation of the rewards offered by their plants, and that they would not exhibit risk-sensitivity in natural patches, such as has been previously showed for broad-tailed hummingbirds (Selasphorus platycercus) and Rufous hummingbirds (S. rufus) visiting flowers of scarlet gilia (Ipomopsis aggregate) in the Rocky Mountains of Colorado, USA (Waser and McRobert, 1998). In this way, hummingbirds and other nectarivore foragers behaving as risk-prone should take risks differently and act differently than those being risk adverse. A hummingbird should change its foraging behavior depending on this, and changes in foraging behavior should affect its energy budget. Therefore, when considering that most hummingbird species live on the limit of a negative energy balance, we could explain why the hummingbirds are risk indifferent. Thus, if a bird has no energy, and has high metabolic costs, it should take risks, and (as in our study) that involves going to all the arrays the same number of times.
Our results do not preclude the possibility that hummingbirds respond to nectar variability at other levels of the spatial scale (e.g., individual plants, groups of plants, vegetation patches, or landscape units; Cotton, 2007; Ortiz-Pulido and Vargas-Licona, 2008). Future investigations of risk-sensitivity in hummingbirds including spatial arrangement of flowers, the sensory cues, and the handling requirements will prove revealing.
We gratefully acknowledge Vanessa Martínez, Jessyca González, and María Cristina Tello for field assistance, and the staff of the Estación Científica “La Malinche”, Tlaxcala for access to their facilities. All hummingbirds were studied after receiving ethical approval from the relevant local authorities (SEMARNAT). We thank Kurt Hoffman and two anonymous reviewers for their helpful comments and grammar revision. This work was supported by a scholarship from the Consejo Nacional de Ciencia y Tecnología, México (No. 201851) to L. Gómez as part of thesis work to obtain master's degree in Maestría en Ciencias Biológicas, Universidad Autónoma de Tlaxcala.