








En este trabajo se ha comparado la organización de los lóbulos antenales (LAs) y su representación topográfica en los cuerpos fungiformes (CFs) en soldados de dos especies de hormigas, Atta mexicana y Camponotus ocreatus. La comparación morfológica sugiere que los LAs de A. mexicana podrían ser más especializados que los de C. ocreatus a expensas de capacidades más generales. En ambas especies, el LA está organizado en seis grupos de glomérulos, cada uno inervado por su propio tracto. La inervación de las neuronas de proyección (NP), las cuales conectan los LAs con los CFs a través de los tractos antenoprotocerebrales (TAP), divide el LA en dos regiones, anterior y posterior. Los cálices de los CFs tienen varias capas que reciben información olfativa segregada: NP que inervan los grupos anteriores de glomérulos envían axones a través del TAP-lateral hacia el cuerno lateral (CL) y después hacia la capa interna del cáliz, mientras que las NP que inervan los grupos posteriores de glomérulos envían axones a través del TAP-medial a la capa externa del cáliz y después al CL. Estos resultados muestran que la organización del LA está representada topográficamente en los CFs formando un mapa odotópico. Las dos vías que conectan los LAs con los CFs podrían representar dos canales para el procesamiento de diferentes clases de olores o diferentes propiedades de los estímulos olfativos. Los resultados neuroanatómicos sugieren una organización funcional conservada en los himenópteros sociales.
In this work, the organization of the antennal lobes (ALs) and its topographic representation in the mushroom bodies (MBs) in soldiers of two species of ants has been compared. Morphologic comparison between these species suggests that ALs of A. mexicana could be more specialized than that of C. ocreatus at the expense of general capabilities. In both species the AL is organized in six groups of glomeruli, each of which innervated by its own tract. Glomeruli clusters are grouped in two regions, anterior and posterior, by the innervation of projection neurons (PNs) that connect AL with MBs via antennoprotocerebral tracts (APT). Calyces of MBs are layered and receive segregated olfactory information: PNs innervating anterior glomeruli clusters send axons through lateral-APT to the lateral horn (LH) and then to inner layer of the calyx, while PNs innervating posterior glomeruli clusters send axons through medial-APT to inner layers of the calyx and then to the LH. These results show that AL organization is topographically represented in the MB forming a coarse odotopic map. The two pathways connecting AL with MB could represent two channels for processing different kinds of odors or different properties of olfactory stimuli. The neuroanatomical results suggest a conserved functional organization in social hymenopterans.
Las hormigas son insectos olfativos: su organización social depende de la emisión y detección de señales químicas llamadas feromonas. Además, alimento, presas, enemigos, compañeros de nido y estímulos químicos del ambiente involucrados en la orientación también son detectados por órganos quimiosensoriales, llamados sensilas, que se ubican en las antenas1. Esta plétora de estímulos químicos genera un flujo constante de información que es procesada en diferentes centros del cerebro. El sistema olfativo de las hormigas debe ser capaz de detectar y discriminar las complejas mezclas de feromonas sociales y señales químicas del ambiente para generar el comportamiento adecuado de acuerdo al contexto.
Se conocen diversos tipos de sensilas por su morfología y respuesta a los odorantes2,3. El número y distribución de sensilas es estereotipado, varía con la especie, el sexo, y la casta en insectos sociales4–8. Dentro de las diferentes clases de sensilas se encuentran combinaciones, también estereotipadas, de diferentes tipos de neuronas receptoras olfativas (NRO)3,6,7, que expresan sólo uno de las decenas de genes receptores que puede presentar cada especie8–10.
Las NRO que expresan los mismos receptores envían sus axones a través del nervio antenal (NA) a uno de los numerosos glomérulos (estructuras esferoidales de neuropilo sináptico) del lóbulo antenal (LA), en donde la información es procesada por primera vez en el cerebro8,11. Cuando el NA entra al LA, se divide en varios tractos que inervan diferentes grupos de glomérulos. La organización de estos grupos de glomérulos forma un mapa topográfico de la distribución periférica de los receptores10,12. En este mapa quimiotópico6, la información olfativa se codifica en patrones espaciales y temporales de actividad glomerular en el LA11,13–16.
En los glomérulos, los axones de las NRO hacen contacto sináptico, ya sea con las dendritas de neuronas locales (NL) que conectan glomérulos entre sí, o bien con neuronas de proyección (NP) que conectan el LA con los cuerpos fungiformes (CFs) y el cuerno lateral (CL)11,13. Los CFs son un par de centros cerebrales superiores, particularmente grandes en los himenópteros (principalmente en los grupos sociales17), que controlan el comportamiento y la plasticidad17–21, y en donde ocurre la convergencia sensorial22. Los CFs de los himenópteros están compuestos por miles de neuronas intrínsecas, llamadas células Kenyon, cuyas dendritas forman una región en forma de copa llamada cáliz que recibe aferencias sensoriales17. Los cálices reciben aferencias de las NP olfativas que se originan en los LAs ipsilaterales22,23. En los himenópteros, los cálices también reciben aferencias de los lóbulos ópticos en ambos lados del cerebro24–27. En las abejas, hormigas y avispas, el cáliz se encuentra concéntricamente subdividido en tres compartimientos: el labio que recibe aferencias del LA, el collar que recibe aferencias visuales, y el anillo basal que está dividido en dos capas, una recibe aferencias visuales y la otra antenales22,27,28.
Varios tractos conectan los LAs con los CFs, pero los más prominentes son los tractos antenoprotocerebrales medial (TAP-m) y lateral (TAP-l), compuestos por axones de NP que hacen sinapsis uniglomerulares. Hay otros tractos más pequeños compuestos por axones de NP multiglomerulares que inervan el protocerebro pero no los cálices11,13,29–30. Se ha descrito que las eferencias de los lóbulos antenales, constituidas por axones de NP uniglomerulares, dividen al LA en dos regiones, una anterior y una posterior, descrito primero para hormigas por López-Riquelme, et al. (2004)31, Gronenberg & López-Riquelme (2004)22 y López-Riquelme (2008)32 y después confirmados para otros himenópteros por otros autores33–35. Los axones de estas NP se separan formando los dos tractos principales, el TAP-l y el TAP-m, que terminan en regiones también segregadas de los CFs, en donde forman un mapa odotópico22,31,32, y del cuerno lateral (CL)8.
No obstante que se han realizado trabajos importantes sobre las vías olfativas, y que recientemente hay un interés creciente por el estudio del sistema olfativo en diversas especies en las que se ha encontrado una gran convergencia36,37, es necesario, para entender la codificación olfativa, así como los mecanismos que subyacen al reconocimeinto de olores y al comportamiento guiado por el olfato, dilucidar la organización neuroanatómica de las vías olfativas. En el presente trabajo se presenta un estudio comparativo de la organización del LA, de sus vías de conexión con el protocerebro, y de su representación odotópica en los CFs, en dos especies de hormigas: Camponotus ocreatus y Atta mexicana.
Materiales y MétodosInsectosLas hormigas carpinteras (Camponotus ocreatus) fueron colectadas en Coronado National Forest en las montañas de Catalina, Tucson Az, EUA, mientras que las hormigas arrieras (Atta mexicana) se colectaron en Yecapixtla, Morelos, México. Los insectos fueron mantenidos en laboratorio bajo un ciclo luz-oscuridad 12:12, a 25°C. Las hormigas carpinteras fueron alimentadas con grillos en trozos y agua con miel; y a las hormigas arrieras se les suministró sustrato vegetal para el cultivo de su hongo.
Trazado neuralPara determinar la estructura del LA y hacer visibles los glomérulos, se realizó un trazado masivo desde el nervio antenal (NA) de acuerdo con Gronenberg & López-Riquelme (2004)22 y López-Riquelme (2008)32. Las preparaciones resultantes se visualizaron y digitalizaron mediante microscopía confocal. Brevemente, las hormigas y sus antenas fueron inmovilizadas en cera y el NA fue expuesto para colocar cristales de biocitina sobre él. Después de la migración del trazador (12h), los cerebros fueron extraídos y fijados con formaldehído al 4%, lavados en PBS (pH 6.9) y deshidratados en alcoholes graduales. Después de esto, los cerebros fueron rehidratados y lavados en PBS y pasados a una solución digestiva (1mg/mL colagenasa, 1 mg/mL hialuronidasa y 0.1 Triton en PBS) por 30min. Posteriormente, los cerebros fueron lavados con PBS y transferidos a una solución de avidina conjugada con rojo Texas y 0.5% Triton-X en PBS por 12h. Finalmente, los cerebros se lavaron en PBS, se deshidrataron con dimetoxipropano y se montaron como cerebros completos en portaobjetos excavados con metilsalicilato. Se obtuvieron imágenes de estas preparaciones mediante microscopía confocal láser (Nikon PCM 2000 con Ar y láser HeNe y una combinación apropiada de filtros) realizando secciones ópticas cada 2μm, las cuales se procesaron con el programa Adobe Photoshop 5.0. De cada imagen se trazó un dibujo directamente sobre el monitor.
Para marcar las conexiones desde los LAs hasta los CFs, se inyectaron dextranes acoplados Rojo Texas o Fluoresceina (MW 3000, Molecular Probes) en diferentes regiones de los LAs. Después de la difusión del trazador (4–12h), los cerebros fueron extraídos y fijados en formaldehído, deshidratados, incluidos en Spurr's y seccionados con un grosor de entre 15 y 20μm22,32. Las secciones fueron observadas con microscopía de epifluorescencia. Se ajustó el brillo y contraste de estas imágenes empleando el programa Adobe Photoshop 5.0.
ResultadosOrganización aferente del lóbulo antenalLa distribución de los tractos, así como el patrón de inervación de los grupos de glomérulos es idéntica en ambas especies de hormigas. A partir de las micrografías confocal, se observó que el LA tiene un centro sin glomérulos por donde pasan los tractos provenientes del NA. Rodeando esta región, en las mismas micrografías confocal, se observaron los glomérulos organizados en grupos. Además, pudieron distinguirse diferentes grupos de somas de neuronas locales. Se identificaron cinco principales grupos de somas en el LA (Figura 1): 1) Grupo de Neuronas Centrales Medial-posterior; 2) Grupo de Neuronas Centrales lateral; 3) Grupo de Neuronas Centrales medial; 4) Grupo de Neuronas Centrales dorsal (el más grande), y 5) Grupo de Neuronas Centrales anterior (el más pequeño).
 que rodean a los glomérulos de soldados de C. ocreatus y A. mexicana. (a) Micrografías de confocal de C. ocreatus tomadas a 60x con los detalles de los diferentes grupos de somas de neuronas del LA. Nótese cómo los grupos de somas neuronales rodean la zona de glomérulos del LA. Barra de escala: 50 μm. (b y c) Micrografías de confocal del LA de C. ocreatus (b) y A. mexicana (c) con la localización de los somas de los grupos de neuronas centrales indicados por los contornos de líneas blancas. Debido a la magnificación de la imagen (20x) no pueden observarse los somas tan claramente como en las micrografías de (a). Los números en la esquina izquierda de cada micrografía corresponden al número de la sección óptica. Barra de escala: 100 μm. Trazado neuronal con neurobiotina y avidina conjugada con rojo Texas (ver Materiales y Métodos). NA, nervio antenal; d, dorsal; v, ventral; l, lateral; m, medial.")
Micrografías del LA tomadas mediante microscopía confocal en las que se muestra la localización de los grupos de neuronas centrales (GNC) que rodean a los glomérulos de soldados de C. ocreatus y A. mexicana. (a) Micrografías de confocal de C. ocreatus tomadas a 60x con los detalles de los diferentes grupos de somas de neuronas del LA. Nótese cómo los grupos de somas neuronales rodean la zona de glomérulos del LA. Barra de escala: 50 μm. (b y c) Micrografías de confocal del LA de C. ocreatus (b) y A. mexicana (c) con la localización de los somas de los grupos de neuronas centrales indicados por los contornos de líneas blancas. Debido a la magnificación de la imagen (20x) no pueden observarse los somas tan claramente como en las micrografías de (a). Los números en la esquina izquierda de cada micrografía corresponden al número de la sección óptica. Barra de escala: 100 μm. Trazado neuronal con neurobiotina y avidina conjugada con rojo Texas (ver Materiales y Métodos). NA, nervio antenal; d, dorsal; v, ventral; l, lateral; m, medial.
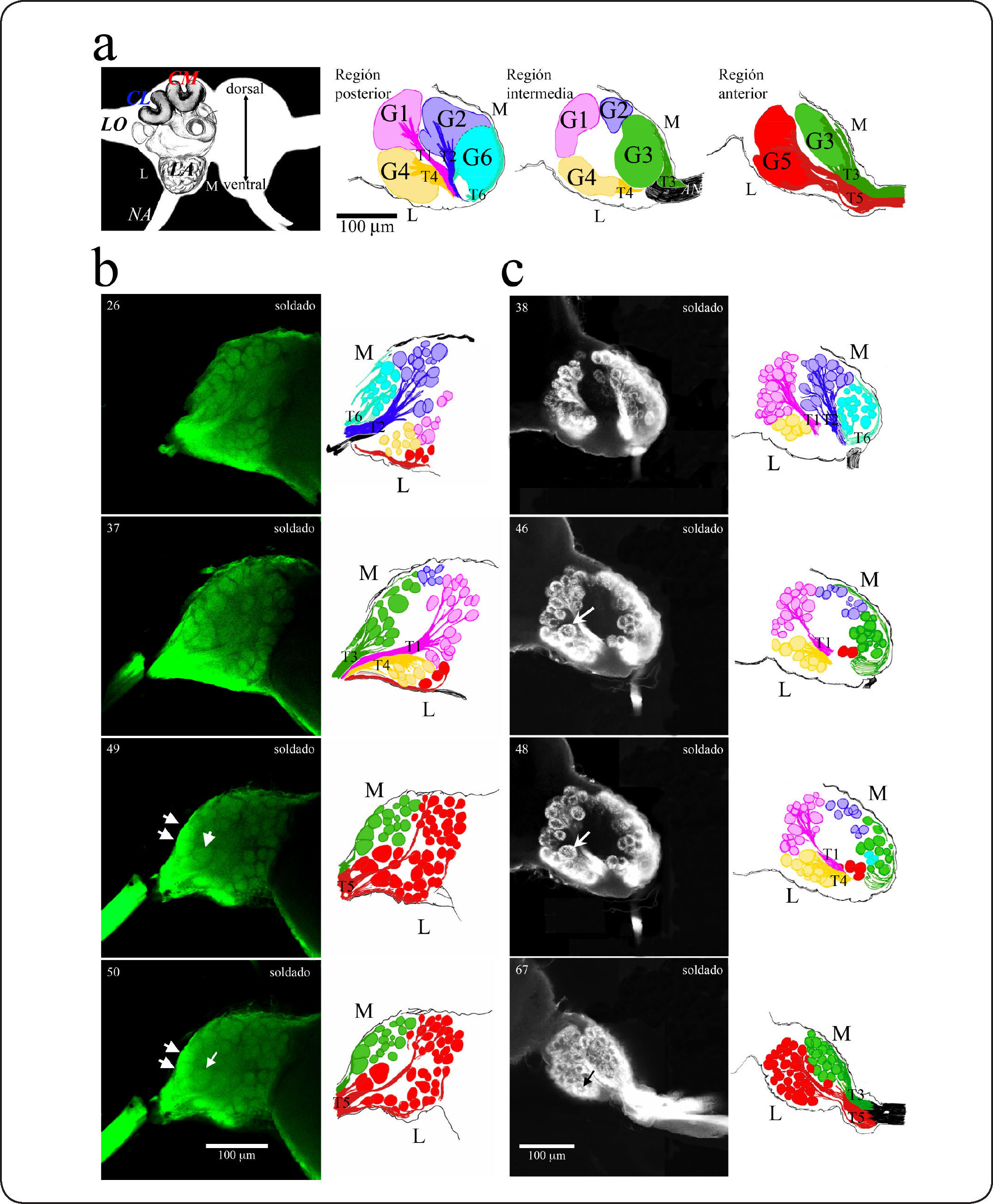
Los axones de las NRO, los cuales salen de cada sensila, permanecen juntos en el NA hasta que este último entra al LA en donde se divide en 8 tractos principales, dos de los cuales inervan el centro motor y mecanosensorial antenal (CMMA), ubicado en la región posterior del LA, mientras que los otros 6 tractos (T1-T6) inervan seis grupos (G1-G6) de glomérulos.
La posición de los grupos glomerulares y la trayectoria de los tractos fue determinada por criterios morfológicos y topográficos (Figura 2). Los grupos de glomérulos y sus tractos se numeraron de acuerdo a la nomenclatura descrita en abejas29,38,39. Sin embargo, en las abejas sólo se han descrito 4 grupos de glomérulos y 4 tractos, y en las hormigas estudiadas en este trabajo se encontraron 6 grupos (G1-G6) y 6 tractos (T1-T6). Los tractos T1-T4 de las abejas pueden reconocerse en las hormigas aproximadamente en las mismas posiciones relativas. En las hormigas se encontraron dos tractos y dos grupos de glomérulos más: T5-G5 y T6-G6. A continuación se describen los grupos de glomérulos y sus tractos (Figura 2):
- 1)
Tracto T1 y grupo G1. T1 es un tracto ancho y largo que pasa diagonalmente a través del centro del LA en su región intermedia. Inicia en el área ventral y lateral del NA y corre a través del LA hasta alcanzar el grupo G1 de glomérulos, el cual es el grupo más dorsal y más grande. En su región más ventral y frontal, T1 se une al tracto T4, formando parte de un tracto grueso que proviene del NA cuando éste entra al LA.
- 2)
Tracto T2 y grupo G2. T2 se encuentra en la región intermedia, pero posterior a T1. También es un tracto ancho y largo que atraviesa diagonalmente el centro del LA desde su origen hasta el grupo G2 que está localizado en la región dorsal y medial.
- 3)
Tracto T3 y grupo G3. T3 se origina en la región anterior medial a partir del NA cuando ingresa al LA. Aunque su porción distal está unida al tracto T5, se separa inmediatamente y se dirige hacia la región medial del LA por la periferia rodeando e inervando al grupo G3 (no atraviesa el centro del LA), el cual es un pequeño grupo situado en la región frontal y medial del LA.
- 4)
Tracto T4 y grupo G4. T4 se encuentra unido distalmente en su base con el T1 justo cuando el NA entra al LA en la región intermedia. Es un tracto corto pero grueso claramente distinguible del T1 que se separa rápidamente de este último y que recorre una corta distancia por el centro del LA en su región intermedia lateral distribuyendo axones a los glomérulos del grupo G4, el cual es un grupo grande que se ubica en la región intermedia lateral, junto al grupo de neuronas centrales lateral (GNC lateral).
- 5)
Tracto T5 y grupo G5. Tanto el tracto T5 como el grupo G5 no se presentan en abejas, por lo que es probable que sea exclusivo de Formicidae. En su porción distal, T5 está unido a T3, y al separarse continúa en línea recta y se separa en dos pequeños tractos; uno de ellos recorre la parte central del LA. La otra porción del T5 se dirige hacia la periferia de la región lateral del LA. Las dos porciones del T5 inervan al grupo G5 de glomérulos, el más frontal de los otros grupos de glomérulos, localizado en la región anterior y ventral del LA.
- 6)
Tracto T6 y grupo G6. Tampoco se presentan en abejas. Es un tracto delgado que se forma a partir del NA y se dirige hacia la región más posterior por la periferia del LA en su región medial. Inerva al pequeño grupo G6 que se localiza en la región ventral y posterior, y hacia la región medial, muy cerca del CMMA.
 Panel izquierdo. Dibujo del contorno de un cerebro de hormiga que muestra las regiones medial-lateral, anterior-posterior, y ventral-dorsal, así como las estructuras principales. LO: lóbulo óptico; CFs: cuerpos fungiformes; LA: lóbulo antenal; NA: nervio antenal. L: región lateral del cerebro; M: región medial; CM: cáliz medial; CL: cáliz lateral. Panel derecho. Dibujos esquemáticos del LA realizados a partir de micrografías confocal con la localización topográfica de los grupos de glomérulos y sus tractos. Los grupos de glomérulos fueron definidos tanto por su posición como por el tracto que los inerva: cada grupo de glomérulos tiene su propio tracto derivado del NA. Se han descrito 6 grupos principales de glomérulos en las especies A. mexicana y C. ocreatus esquematizados en los dibujos en color. (b) Columna izquierda. Micrografías de confocal del LA izquierdo de soldados de A. mexicana teñidas con neurobiotina y avidina conjugada con rojo Texas. Columna derecha. Dibujos realizados a partir de cada micrografía de confocal. Los grupos de glomérulos se indican con diferentes colores. La posición relativa de los grupos de glomérulos se mantiene constante entre los soldados. De arriba hacia abajo: Sección 26: en esta figura se observan los grupos G2 y G3 y algunos glomérulos del grupo G1, G4 y G5. Sección 37: en esta figura se muestra el grupo glomerular G4. Puede apreciarse que, como en C. ocreatus, en su base hacia la parte anterior, el tracto T3, en su parte más ventral, se une al tracto T1, y al T4 que inerva al grupo G4. Sección 49: en esta figura se muestra la región inicial del último grupo de glomérulos, el grupo G5, con su tracto correspondiente, el tracto T5. Este tracto y el tracto T3, se separan inmediatamente cuando el NA entra al LA y se dirigen más hacia la parte anterior, inervando grupos de glomérulos diferentes. El tracto T5 inerva a un grupo anterior de glomérulos que pueden verse en la parte central del LA. El tracto T3 se divide para dar lugar a una parte que se dirige hacia la región medial e inerva al grupo G3. Las flechas indican los macroglomérulos. Sección 50: se observan los grupos G3 y G5. Comparando el tamaño aparente de la mayoría de los glomérulos en A. mexicana, puede apreciarse que algunos glomérulos son más grandes (flechas) en el grupo G3 y G5. (c) Columna izquierda. Micrografías de confocal de los LAs de soldados de C. ocreatus teñidas con neurobiotina y avidina conjugada con rojo Texas. Columna derecha. Dibujos realizados a partir de cada micrografía confocal. De arriba hacia abajo: Sección 38: micrografía confocal y dibujo donde pueden apreciarse los grupos glomerulares G1, G2, G6 con sus respectivos tractos T1, T2 y T6. Además, algunos glomérulos del grupo G4 cuyo tracto T4 no se distingue en este nivel. Los glomérulos del G2 que se muestran en el dibujo y que no se aprecian claramente en la micrografía, se trazaron incrementando el brillo de la micrografía. Sección 46: en esta figura se ve el grupo G3 y algunos glomérulos del grupo G5. El grupo G3 es inervado por un tracto que se separa del resto de las fibras del NA inmediatamente después de haber entrado al LA. La base del tracto T1 está unida al tracto T4, que es corto, grueso y brillante. Se observan los glomérulos del grupo G4 y un glomérulo más grande (flecha). Sección 48: en esta figura se muestran los grupos G1 y G4, y la unión de los tractos T1 y T4. En el grupo G4 se encuentra un gran glomérulo (flecha) cerca del tracto T1 y del grupo G1. Sección 67: Grupo G5, con su tracto correspondiente, el tracto T5; este tracto y el tracto T3, se separan inmediatamente al entrar al LA. El tracto T5 inerva a un grupo anterior de glomérulos que puede verse en la parte central del LA. El tracto T3 se dirige hacia la región medial e inerva al grupo G3. El grupo 5 muestra un glomérulo más grande hacia la parte lateral (flecha). Los números en la esquina izquierda de cada micrografía en (b) y (c) corresponden al número de la sección óptica de microscopía confocal.")
Organización glomerular del lóbulo antenal en soldados de A. mexicana y C. ocreatus.
Descripción de la Figura 2. (a) Panel izquierdo. Dibujo del contorno de un cerebro de hormiga que muestra las regiones medial-lateral, anterior-posterior, y ventral-dorsal, así como las estructuras principales. LO: lóbulo óptico; CFs: cuerpos fungiformes; LA: lóbulo antenal; NA: nervio antenal. L: región lateral del cerebro; M: región medial; CM: cáliz medial; CL: cáliz lateral. Panel derecho. Dibujos esquemáticos del LA realizados a partir de micrografías confocal con la localización topográfica de los grupos de glomérulos y sus tractos. Los grupos de glomérulos fueron definidos tanto por su posición como por el tracto que los inerva: cada grupo de glomérulos tiene su propio tracto derivado del NA. Se han descrito 6 grupos principales de glomérulos en las especies A. mexicana y C. ocreatus esquematizados en los dibujos en color. (b) Columna izquierda. Micrografías de confocal del LA izquierdo de soldados de A. mexicana teñidas con neurobiotina y avidina conjugada con rojo Texas. Columna derecha. Dibujos realizados a partir de cada micrografía de confocal. Los grupos de glomérulos se indican con diferentes colores. La posición relativa de los grupos de glomérulos se mantiene constante entre los soldados. De arriba hacia abajo: Sección 26: en esta figura se observan los grupos G2 y G3 y algunos glomérulos del grupo G1, G4 y G5. Sección 37: en esta figura se muestra el grupo glomerular G4. Puede apreciarse que, como en C. ocreatus, en su base hacia la parte anterior, el tracto T3, en su parte más ventral, se une al tracto T1, y al T4 que inerva al grupo G4. Sección 49: en esta figura se muestra la región inicial del último grupo de glomérulos, el grupo G5, con su tracto correspondiente, el tracto T5. Este tracto y el tracto T3, se separan inmediatamente cuando el NA entra al LA y se dirigen más hacia la parte anterior, inervando grupos de glomérulos diferentes. El tracto T5 inerva a un grupo anterior de glomérulos que pueden verse en la parte central del LA. El tracto T3 se divide para dar lugar a una parte que se dirige hacia la región medial e inerva al grupo G3. Las flechas indican los macroglomérulos. Sección 50: se observan los grupos G3 y G5. Comparando el tamaño aparente de la mayoría de los glomérulos en A. mexicana, puede apreciarse que algunos glomérulos son más grandes (flechas) en el grupo G3 y G5. (c) Columna izquierda. Micrografías de confocal de los LAs de soldados de C. ocreatus teñidas con neurobiotina y avidina conjugada con rojo Texas. Columna derecha. Dibujos realizados a partir de cada micrografía confocal. De arriba hacia abajo: Sección 38: micrografía confocal y dibujo donde pueden apreciarse los grupos glomerulares G1, G2, G6 con sus respectivos tractos T1, T2 y T6. Además, algunos glomérulos del grupo G4 cuyo tracto T4 no se distingue en este nivel. Los glomérulos del G2 que se muestran en el dibujo y que no se aprecian claramente en la micrografía, se trazaron incrementando el brillo de la micrografía. Sección 46: en esta figura se ve el grupo G3 y algunos glomérulos del grupo G5. El grupo G3 es inervado por un tracto que se separa del resto de las fibras del NA inmediatamente después de haber entrado al LA. La base del tracto T1 está unida al tracto T4, que es corto, grueso y brillante. Se observan los glomérulos del grupo G4 y un glomérulo más grande (flecha). Sección 48: en esta figura se muestran los grupos G1 y G4, y la unión de los tractos T1 y T4. En el grupo G4 se encuentra un gran glomérulo (flecha) cerca del tracto T1 y del grupo G1. Sección 67: Grupo G5, con su tracto correspondiente, el tracto T5; este tracto y el tracto T3, se separan inmediatamente al entrar al LA. El tracto T5 inerva a un grupo anterior de glomérulos que puede verse en la parte central del LA. El tracto T3 se dirige hacia la región medial e inerva al grupo G3. El grupo 5 muestra un glomérulo más grande hacia la parte lateral (flecha). Los números en la esquina izquierda de cada micrografía en (b) y (c) corresponden al número de la sección óptica de microscopía confocal.
Los grupos de glomérulos y los tractos que los inervan pueden clasificarse en dos: 1) Grupos de glomérulos inervados por tractos que atraviesan el LA por el centro de neuropilo: G1- T1, G2-T2 y G4-T4; y 2) Grupos de glomérulos inervados por tractos que rodean el centro de neuropilo y a los glomérulos: G3-T3, G5-T5 y G6-T6.
Aunque la estructura de los LAs en A. mexicana y C. ocreatus es muy similar, existen diferencias importantes. Los soldados de A. mexicana pueden presentar de 305 a 313 glomérulos y en promedio 305±4 glomérulos (n=3), mientras que los soldados de C. ocreatus de 376 a 385, y en promedio 380±4 (n=3). Además, en los soldados de A. mexicana se observaron glomérulos pequeños y grandes, así como la presencia de 3 macroglomérulos en los grupos G3 y G5 (Figura 2). En contraste, los glomérulos de C. ocreatus se observan más uniformes en tamaño y forma y los tractos más gruesos. En esta especie hay dos macroglomérulos, uno en el grupo glomerular G4 y otro en el G5 (Figura 2).
Tractos de los LAs hacia los CFsLa mayoría de las vías eferentes del LA terminan en el protocerebro ipsilateral, y aunque se han descrito tractos que proyectan hacia el LA contralateral29, no se encontraron en este trabajo. A partir del trazado neuronal desde el LA, se encontró que, en ambas especies, los axones que salen del LA se dirigen hacia el protocerebro ipsilateral a través de tres tractos principales: 1) tracto antenoprotocerebral medial (TAP-m); 2) tracto antenoprotocerebral lateral (TAP-l), y 3) tracto antenoprotocerebral medial-lateral (TAP-ml). De éstos, los dos primeros son los más prominentes. Se observó que las eferencias del LA hacia los CFs se segregan y terminan en diferentes regiones particulares del labio de los cálices en los CFs. Pudieron distinguirse al menos tres capas en el labio del cáliz, y dos rutas por las que los axones entran al cáliz y se dirigen hacia el labio y al cuerno lateral (CL).
En la Figura 3 se muestra una sección frontal del cerebro de una hormiga soldado C. ocreatus en microscopía de fluorescencia con los tractos que conectan el LA con el protocerebro: el TAP-m hacia la parte medial de la sección, y el TAP-l hacia la parte lateral y un delgado TAP-ml hacia el CL. Los axones de las NP corren ya sea por el TAP-l o por el TAP-m dirigiéndose hacia el labio del cáliz (que recibe aferencias exclusivamente olfativas) pasando por el anillo basal (que recibe aferencias tanto olfativas como visuales). Cuando lo hacen por el TAP-m, los axones se dirigen directamente hacia el labio del cáliz de los CFs y de ahí envían colaterales hacia el CL; mientras que cuando lo hacen por el TAP-l, se dirigen primero hacia el CL y, después de hacer un relevo ahí, van hacia el labio del cáliz. Además, en las Figuras 3 y 4 también se muestran las tres principales capas que componen al labio que reciben aferencias segregadas desde el LA (ver más adelante): 1) capa externa; 2) capa intermedia (no pudo identificarse su origen), y 3) capa interna.
 Micrografía de epifluorescencia de una sección que muestra la mitad derecha del cerebro de un soldado C. ocreatus. Se ha trazado anterógradamente la vía sensorial desde los LAs (en donde se ha inyectado el trazador) hasta el labio del cáliz de los CFs con dextranes acoplados con Rojo-Texas (ver Materiales y Métodos). Pueden verse los tractos antenoprotocerebrales que conectan los LAs con los CFs: TAP-l, TAP-m y TAP-ml. Se ve el TAP-m que recorre la parte medial, dividiéndose en dos ramas, una de ellas se dirige hacia el CL por la parte ventral y formando el TAP-ml, mientras que la otra va hacia los CFs. Hacia la parte lateral, se observa el TAP-l que va primero hacia el CL y de ahí, hacia los cálices de los CFs. En la imagen pueden apreciarse también las tres capas en las que se divide el labio, recibiendo aferencias segregadas desde los LAs. Hacia abajo de la micrografía, está el lóbulo antenal (LA). La región más brillante del LA corresponde a la zona en la que se ha inyectado el fluorocromo. Hacia la derecha y hacia abajo del LA, están los glomérulos con su característica forma esferoidal. Puede apreciarse también, que todos los axones de las NP se unen formando una sola vía de salida que, tras abandonar el LA, inmediatamente se divide en dos gruesos tractos (el TAP-m y el TAP-l) que se dirigen al protocerebro. (b-e) Micrografías de epifluorescencia de los cálices de los CFs marcados anterógradamente desde los LAs. En (b) pueden apreciarse tres distintas capas (numeradas sobre cada una) en el labio y que constituyen las aferencias desde los LAs. De las tres capas, la más gruesa es la interna (e), seguida por la capa externa (c), mientras que la más delgada es la capa intermedia (d). La capa intermedia es difícil de marcar por separado, lo cual sugiere que los grupos de glomérulos que envían axones a estas dos capas (la interna y la intermedia) del labio se encuentran ubicados muy próximos o que los glomérulos que la inervan pertenecen al mismo grupo glomerular. Por el contrario, la capa interna es fácil de separar de la capa externa. Esto indica que las NP que conectan las dos regiones del LA con el cáliz están separadas formando las dos regiones eferentes del LA.")
(a) Micrografía de epifluorescencia de una sección que muestra la mitad derecha del cerebro de un soldado C. ocreatus. Se ha trazado anterógradamente la vía sensorial desde los LAs (en donde se ha inyectado el trazador) hasta el labio del cáliz de los CFs con dextranes acoplados con Rojo-Texas (ver Materiales y Métodos). Pueden verse los tractos antenoprotocerebrales que conectan los LAs con los CFs: TAP-l, TAP-m y TAP-ml. Se ve el TAP-m que recorre la parte medial, dividiéndose en dos ramas, una de ellas se dirige hacia el CL por la parte ventral y formando el TAP-ml, mientras que la otra va hacia los CFs. Hacia la parte lateral, se observa el TAP-l que va primero hacia el CL y de ahí, hacia los cálices de los CFs. En la imagen pueden apreciarse también las tres capas en las que se divide el labio, recibiendo aferencias segregadas desde los LAs. Hacia abajo de la micrografía, está el lóbulo antenal (LA). La región más brillante del LA corresponde a la zona en la que se ha inyectado el fluorocromo. Hacia la derecha y hacia abajo del LA, están los glomérulos con su característica forma esferoidal. Puede apreciarse también, que todos los axones de las NP se unen formando una sola vía de salida que, tras abandonar el LA, inmediatamente se divide en dos gruesos tractos (el TAP-m y el TAP-l) que se dirigen al protocerebro. (b-e) Micrografías de epifluorescencia de los cálices de los CFs marcados anterógradamente desde los LAs. En (b) pueden apreciarse tres distintas capas (numeradas sobre cada una) en el labio y que constituyen las aferencias desde los LAs. De las tres capas, la más gruesa es la interna (e), seguida por la capa externa (c), mientras que la más delgada es la capa intermedia (d). La capa intermedia es difícil de marcar por separado, lo cual sugiere que los grupos de glomérulos que envían axones a estas dos capas (la interna y la intermedia) del labio se encuentran ubicados muy próximos o que los glomérulos que la inervan pertenecen al mismo grupo glomerular. Por el contrario, la capa interna es fácil de separar de la capa externa. Esto indica que las NP que conectan las dos regiones del LA con el cáliz están separadas formando las dos regiones eferentes del LA.
 Micrografías de epifluorescencia de varias secciones de cerebro de soldados de C. ocreatus marcadas con trazadores (dextran acoplado con Rojo Texas) desde el LA. Pueden verse las diferentes capas de los CFs en donde terminan los tractos antenoprotocerebrales. (a) Detalle del cáliz de los CFs con la capa interna marcada con fluorescencia. (b) Sección de la mitad derecha del cerebro de una hormiga C. ocreatus. Se observa la vía, en este caso el TAP-l, que siguen los axones de las NP que conectan el LA con la capa interna. (c-e) Micrografías en las que se muestra el cáliz de los CFs con la capa externa marcada con fluorescencia.")
(a-e) Micrografías de epifluorescencia de varias secciones de cerebro de soldados de C. ocreatus marcadas con trazadores (dextran acoplado con Rojo Texas) desde el LA. Pueden verse las diferentes capas de los CFs en donde terminan los tractos antenoprotocerebrales. (a) Detalle del cáliz de los CFs con la capa interna marcada con fluorescencia. (b) Sección de la mitad derecha del cerebro de una hormiga C. ocreatus. Se observa la vía, en este caso el TAP-l, que siguen los axones de las NP que conectan el LA con la capa interna. (c-e) Micrografías en las que se muestra el cáliz de los CFs con la capa externa marcada con fluorescencia.
La organización del LA en seis grupos de glomérulos corresponde con la organización de las aferencias, en seis tractos, provenientes de las NRO de las antenas. Sin embargo, el LA también presenta una organización eferente de acuerdo con las vías que siguen los axones de las NP hacia el protocerebro, ya sea el TAP-l o el TAP-m, de manera que pueden distinguirse dos regiones del LA, en realidad dos mitades, de acuerdo con dichas eferencias (Figuras 5–7):
- 1.
Región posterior. Es la región más caudal, y puede dividirse en:
- a)
Sub-Región posterior y medial. Comprende el tracto T6 y al grupo G6.
- b)
Sub-Región posterior intermedio-medial. Comprende el tracto T2 y el grupo G2.
- a)
- 2.
Región anterior. Está compuesta por dos subregiones:
- a)
Sub-Región intermedio-dorsal y central. Contiene los tractos T1 y T4, y los grupos glomerulares G1 (situado dorsalmente) y G4 (se encuentra lateralmente).
- b)
Sub-Región anterior y ventral. Ésta es la región más anterior, donde se encuentran ubicados los tractos T3 y T5, así como los grupos glomerulares G3 y G5.
- a)
 del LA a la capa externa del cáliz de los CFs en A. mexicana y C. ocreatus. En las micrografías del cerebro de A. mexicana (a) y C. ocreatus (b), se muestra la región posterior (grupos G2 y G6), el TAP-m y la capa externa del labio y del anillo basal de los CFs marcada con fluorescencia. Debajo de cada figura, se ha colocado una serie de secciones del LA al cual se le ha inyectado el fluorocromo para trazar las vías hacia los CFs. (a) Panel superior. Micrografías de confocal del cerebro de un soldado de A. mexicana en la que se observa el TAP-m saliendo del LA hacia los CFs, inervando primero la capa externa del labio y del anillo basal del cáliz medial, después del cáliz lateral, y finalmente inerva el CL, en donde termina. Panel inferior. Serie de micrografías de confocal del LA del cerebro mostrado en el panel superior. Hacia la izquierda, y con una numeración menor, están las de la región anterior del LA, mientras que hacia la derecha, y con una numeración mayor, están las de la región posterior. Los grupos glomerulares G2 y G6 de la región posterior están marcados con fluorescencia, mientras que los grupos G1, G3, G4 y G5 de la región anterior no lo están. (b) Panel superior. A la izquierda se muestra una micrografía de epifluorescencia de corte del cerebro de un soldado de C. ocreatus. En la imagen pueden apreciarse las NP que inervan a los glomérulos de los grupos de la región posterior (G2 y G6), así como los axones que se unen para formar el TAP-m que se dirige primero hacia el cáliz medial y después hacia el cáliz lateral de los CFs. Ahí, las colaterales inervan la capa externa del labio y del anillo basal. El TAP-m continúa su camino para terminar en el CL. A la derecha, en el recuadro amarillo se presenta una imagen aumentada de la región del cáliz del cerebro mostrado a la izquierda del panel superior con la capa externa del labio y del anillo basal teñida con fluorescencia. También se observan los axones de las NP que pasan entre los cálices lateral y medial. Panel inferior. Serie de micrografías de epifluorescencia del LA del cerebro mostrado en el panel superior. Al igual que en (a), hacia la izquierda se encuentran las secciones de la región anterior del LA, y hacia la derecha están las de la región posterior del LA. Nótese que, del mismo modo que para A. mexicana, los glomérulos más anteriores (de los grupos G1, G3, G4 y G5) no están teñidos, así como tampoco los grupos de neuronas centrales GNC dorsal, GNC anterior y GNC medial. Por el contrario, los grupos glomerulares G2 y G6, de la región posterior, se encuentran claramente marcados con fluorescencia.")
Vía olfativa medial (TAP-m) del LA a la capa externa del cáliz de los CFs en A. mexicana y C. ocreatus. En las micrografías del cerebro de A. mexicana (a) y C. ocreatus (b), se muestra la región posterior (grupos G2 y G6), el TAP-m y la capa externa del labio y del anillo basal de los CFs marcada con fluorescencia. Debajo de cada figura, se ha colocado una serie de secciones del LA al cual se le ha inyectado el fluorocromo para trazar las vías hacia los CFs. (a) Panel superior. Micrografías de confocal del cerebro de un soldado de A. mexicana en la que se observa el TAP-m saliendo del LA hacia los CFs, inervando primero la capa externa del labio y del anillo basal del cáliz medial, después del cáliz lateral, y finalmente inerva el CL, en donde termina. Panel inferior. Serie de micrografías de confocal del LA del cerebro mostrado en el panel superior. Hacia la izquierda, y con una numeración menor, están las de la región anterior del LA, mientras que hacia la derecha, y con una numeración mayor, están las de la región posterior. Los grupos glomerulares G2 y G6 de la región posterior están marcados con fluorescencia, mientras que los grupos G1, G3, G4 y G5 de la región anterior no lo están. (b) Panel superior. A la izquierda se muestra una micrografía de epifluorescencia de corte del cerebro de un soldado de C. ocreatus. En la imagen pueden apreciarse las NP que inervan a los glomérulos de los grupos de la región posterior (G2 y G6), así como los axones que se unen para formar el TAP-m que se dirige primero hacia el cáliz medial y después hacia el cáliz lateral de los CFs. Ahí, las colaterales inervan la capa externa del labio y del anillo basal. El TAP-m continúa su camino para terminar en el CL. A la derecha, en el recuadro amarillo se presenta una imagen aumentada de la región del cáliz del cerebro mostrado a la izquierda del panel superior con la capa externa del labio y del anillo basal teñida con fluorescencia. También se observan los axones de las NP que pasan entre los cálices lateral y medial. Panel inferior. Serie de micrografías de epifluorescencia del LA del cerebro mostrado en el panel superior. Al igual que en (a), hacia la izquierda se encuentran las secciones de la región anterior del LA, y hacia la derecha están las de la región posterior del LA. Nótese que, del mismo modo que para A. mexicana, los glomérulos más anteriores (de los grupos G1, G3, G4 y G5) no están teñidos, así como tampoco los grupos de neuronas centrales GNC dorsal, GNC anterior y GNC medial. Por el contrario, los grupos glomerulares G2 y G6, de la región posterior, se encuentran claramente marcados con fluorescencia.
 del LA a la capa interna del cáliz de los CFs en A. mexicana y C. ocreatus. Descripción de Figura 6. (a) Panel superior. Micrografía de confocal del LA y CFs de A. mexicana. Arriba. Se observa el TAP-l, el CL, la capa intermedia y la interna del labio de los CFs marcadas. El TAP-l se forma inmediatamente que sale del LA y se dirige primero hacia el CL en donde deja colaterales y de ahí hacia la capa interna de los CFs (primero hacia el cáliz lateral y después hacia el medial), para terminar en las capas internas del labio y del anillo basal. Panel inferior. Serie de micrografías de confocal del LA del cerebro mostrado en el panel superior. Se ha inyectado el fluorocromo para trazar la vía hacia los CFs. De igual manera que en la figura 5, hacia la izquierda se encuentran las secciones de la región anterior del LA y hacia la derecha aquellas de la región posterior. Contrario a la Figura 5, los grupos glomerulares G2 y G6, de la región posterior, no se encuentran marcados con fluorescencia, mientras que los grupos G1, G3, G4 y G5 de la región anterior están teñidos. (b) Panel superior. Micrografía de epifluorescencia de corte del cerebro de un soldado de C. ocreatus. Los cálices se han desprendido un poco debido al seccionado en el microtomo. Se observa el TAP-l que se dirige, del LA, primero hacia el CL inervándolo; y de ahí hacia el cáliz lateral y después hacia el cáliz medial en donde, como en A. mexicana, inerva la capa interna del labio y del anillo basal. Se ve la capa interna gruesa y claramente marcada, mientras que la capa externa carece por completo de terminaciones fluorescentes. Esta capa interna consiste, en realidad, de las dos capas, interna y medial, marcadas con fluorescencia, pero que no se lograron teñir por separado, de manera que se han considerado como una sola capa que ha sido designada como capa interna. Nótese que, tanto en los cerebros de A. mexicana como en C. ocreatus, el TAP-m no se encuentra marcado con fluorescencia, indicando dos vías separadas de procesamiento olfativo (véase texto). Panel inferior. Serie de micrografías de epifluorescencia del LA del cerebro mostrado en el panel superior en donde se aprecia la tinción diferencial de los grupos de glomérulos y su relación con las eferencias hacia los CFs en el panel superior. Es importante notar que los grupos de glomérulos de la región anterior G1, G3, G4 y G5 están marcados, pero no los grupos G2 y G6 de la región posterior.")
Vía olfativa lateral (TAP-l) del LA a la capa interna del cáliz de los CFs en A. mexicana y C. ocreatus.
Descripción de Figura 6. (a) Panel superior. Micrografía de confocal del LA y CFs de A. mexicana. Arriba. Se observa el TAP-l, el CL, la capa intermedia y la interna del labio de los CFs marcadas. El TAP-l se forma inmediatamente que sale del LA y se dirige primero hacia el CL en donde deja colaterales y de ahí hacia la capa interna de los CFs (primero hacia el cáliz lateral y después hacia el medial), para terminar en las capas internas del labio y del anillo basal. Panel inferior. Serie de micrografías de confocal del LA del cerebro mostrado en el panel superior. Se ha inyectado el fluorocromo para trazar la vía hacia los CFs. De igual manera que en la figura 5, hacia la izquierda se encuentran las secciones de la región anterior del LA y hacia la derecha aquellas de la región posterior. Contrario a la Figura 5, los grupos glomerulares G2 y G6, de la región posterior, no se encuentran marcados con fluorescencia, mientras que los grupos G1, G3, G4 y G5 de la región anterior están teñidos. (b) Panel superior. Micrografía de epifluorescencia de corte del cerebro de un soldado de C. ocreatus. Los cálices se han desprendido un poco debido al seccionado en el microtomo. Se observa el TAP-l que se dirige, del LA, primero hacia el CL inervándolo; y de ahí hacia el cáliz lateral y después hacia el cáliz medial en donde, como en A. mexicana, inerva la capa interna del labio y del anillo basal. Se ve la capa interna gruesa y claramente marcada, mientras que la capa externa carece por completo de terminaciones fluorescentes. Esta capa interna consiste, en realidad, de las dos capas, interna y medial, marcadas con fluorescencia, pero que no se lograron teñir por separado, de manera que se han considerado como una sola capa que ha sido designada como capa interna. Nótese que, tanto en los cerebros de A. mexicana como en C. ocreatus, el TAP-m no se encuentra marcado con fluorescencia, indicando dos vías separadas de procesamiento olfativo (véase texto). Panel inferior. Serie de micrografías de epifluorescencia del LA del cerebro mostrado en el panel superior en donde se aprecia la tinción diferencial de los grupos de glomérulos y su relación con las eferencias hacia los CFs en el panel superior. Es importante notar que los grupos de glomérulos de la región anterior G1, G3, G4 y G5 están marcados, pero no los grupos G2 y G6 de la región posterior.
 al conectar los grupos de glomérulos (los glomérulos están representados por las estructuras esferoidales grandes en el LA en las que se han esquematizado terminaciones dendríticas de las NP) de la región anterior (G1, G3, G4 y G5) con la capa interna del labio y del anillo basal de los cálices de los CFs a través del TAP-l. En verde se muestra la vía que, a través del TAP-m, conecta los grupos de glomérulos de la región posterior (G2 y G6) con la capa externa del labio y del anillo basal de los cálices de los CFs. Se han esquematizado también las conexiones con el CL. Las aferencias del LA en los CFs forman un mapa odotópico que podría representar dos canales de procesamiento de diferentes familias de odorantes con significado biológico (véase texto). ab: anillo basal; lo: lóbulo; me: médula; CL: cuerno lateral; TAP-l: tracto antenoprotocerebral lateral; TAP-m: tracto antenoprotocerebral medial; L: lateral; M: medial.")
Mapa odotópico. Dibujo esquemático de las dos vías de conexión dede el LA hasta los CFs en el cerebro de las hormigas descritas en este trabajo. En rojo se muestra la vía que forman las NP (pequeñas esferas marcadas con s-NP) al conectar los grupos de glomérulos (los glomérulos están representados por las estructuras esferoidales grandes en el LA en las que se han esquematizado terminaciones dendríticas de las NP) de la región anterior (G1, G3, G4 y G5) con la capa interna del labio y del anillo basal de los cálices de los CFs a través del TAP-l. En verde se muestra la vía que, a través del TAP-m, conecta los grupos de glomérulos de la región posterior (G2 y G6) con la capa externa del labio y del anillo basal de los cálices de los CFs. Se han esquematizado también las conexiones con el CL. Las aferencias del LA en los CFs forman un mapa odotópico que podría representar dos canales de procesamiento de diferentes familias de odorantes con significado biológico (véase texto). ab: anillo basal; lo: lóbulo; me: médula; CL: cuerno lateral; TAP-l: tracto antenoprotocerebral lateral; TAP-m: tracto antenoprotocerebral medial; L: lateral; M: medial.
Las vías eferentes del LA hacia los CFs se encuentran separadas, de tal manera que cada capa labial recibe axones, por una de las dos vías de tractos antenoprotocerebrales, de las NP de una de las dos regiones en las que se organizan las conexiones dendríticas de dichas NP en los grupos de glomérulos del LA. Esto significa que las regiones anterior y posterior están representadas topográficamente en las capas del labio de los CFs estableciendo un mapa odotópico del LA en el protocerebro (Figura 7):
- a)
Región posterior. Los grupos G2 y G6 están inervados por NP de los grupos de neuronas centrales GNC-lateral y GNC M-posterior que envían axones por el TAP-m hacia la capa externa del labio y del anillo basal de los CFs, y de ahí hacia el CL.
- b)
Región anterior. Los grupos G1, G3, G4 y G5 están inervados por NP de los grupos de neuronas centrales GNC-lateral, GNC medial, GNC dorsal y GNC anterior que envían axones por el TAP-l hacia el CL y de ahí hacia la capa interna del labio y del anillo basal de los CFs.
En este trabajo se determinó y comparó la organización del LA de las hormigas A. mexicana y C. ocreatus [descrita por primera vez para hormigas en Gronenberg & López -Riquelme22, López- Riquelme et al.31, López-Riquelme32, y después en Mysore et al.40, Nakanishi et al.41]. La organización espacial gruesa del LA descrita en este trabajo es muy similar a la descrita para las abejas29,38,39, otros himenópteros y la pollilla Manduca sexta13, consistiendo de glomérulos rodeando una zona aglomerular de neuropilo, con grupos de somas neuronales en la periferia del LA (Figura 1). En otras especies de insectos, como en las cucarachas Periplaneta americana, los glomérulos se encuentran distribuidos por todo el LA13. Los LAs de las hormigas son más parecidos a los de las abejas, en las cuales se han descrito cuatro grupos de glomérulos inervados por cuatro tractos29,38,39.
En los soldados de A. mexicana y de C. ocreatus, se encontraron 6 grupos de glomérulos (G1-G6), cada uno inervado por un tracto (T1-T6) derivado del NA (Figura 2). Dos tractos (TCMMA1 y TCMMA2), aparte de los 6 que inervan los grupos de glomérulos, se dirigen hacia el CMMA, por lo que en total, en hormigas se encontraron 8 tractos, 6 de ellos inervan glomérulos olfatorios (T1-T6), mientras que dos se dirigen hacia el CMMA. Algunos tractos, como el T2 que inerva el G2, se subdividen en pequeñas ramificaciones que pueden ser consideradas como tractos independientes 41 generando variaciones en el número de tractos descritos en el LA de las hormigas. Los tractos descritos en este trabajo fueron aquellos derivados del NA al entrar al LA y que inervan grandes grupos de glomérulos.
Se han denominado T5 y T6 a los otros dos tractos que llegan al LA provenientes del NA, por lo que no deben confundirse con los tractos que, en las abejas, se dirigen hacia el CMMA y se han nombrado así. Sin embargo, de acuerdo con características morfológicas y topográficas, tanto los tractos T1-T4, como los grupos de glomérulos G1-G4, podrían ser homólogos en abejas y hormigas. En las abejas se han descrito tres grupos de somas de neuronas centrales13,42, mientras que en las hormigas estudiadas en este trabajo pudieron observarse cinco grupos de somas neuronales. Es muy probable que si existe mayor número de glomérulos en los LAs de las hormigas también haya más neuronas centrales que establezcan conexiones entre glomérulos y entre éstos y el protocerebro. Las abejas son insectos “visuales”, ya que su orientación depende de la visión, mientras que las hormigas son insectos “más olfativos” (no obstante que la organización social de ambos está basada en la comunicación química). Mientras que en las hormigas Atta los LAs pueden llegar a representar hasta el 12 % del total del cerebro y en Camponotus hasta el 10%; en las abejas, los LAs apenas alcanzan un 3% y están compuestos por entre 156 y 166 glomérulos43. La homología entre los LAs de Atta mexicana y Camponotus ocreatus, especies pertenecientes a subfamilias distintas sugiere que la estructura general de los LAs se ha conservado, por lo que debe tener alguna importancia en la vida social basada en el olfato, ya que otros insectos no-sociales pero olfativos13,44 no presentan una organización glomerular como la de los himenópteros. Además, la similitud entre los LAs de abejas y hormigas, sugiere una homología estructural en los himenópteros aculeados. En los himenópteros sociales, los estímulos químicos son los más importantes, ya que la comunicación ocurre dentro de los oscuros nidos y está basada en la emisión y recepción de mezclas complejas de feromonas que regulan el comportamiento del superorganismo. Es posible que esta organización conservada tenga un significado funcional relacionado con la generación del código olfativo y la vida social.
En este trabajo, se encontró que los LAs de los soldados de A. mexicana pueden llegar a tener entre 305 y 313 glomérulos, mientras que los de C. ocreatus entre 376 y 385. Posiblemente, la importancia y número de las aferencias sensoriales provenientes de tipos particulares de NRO esté representada en el tamaño de los glomérulos, mientras que el número de glomérulos por grupo glomerular esté relacionado con las proporciones de los tipos de NRO en las antenas, como en el caso de los macroglomérulos encontrados en los soldados de A. mexicana, o los encontrados en C. ocreatus (Figura 2). Por lo tanto, la organización del LA contendría una representación funcional de las diferentes poblaciones de NRO sobre la antena y de los grupos de olores con significado biológico. Sin embargo, es necesario realizar el trazado desde diferentes sensilas y NRO con los perfiles de respuesta a diferentes olores, tanto de las NRO como de los glomérulos a los que inervan, para determinar el significado funcional de la organización del LA en grupos de glomérulos como se ha hecho para otros insectos10.
Comparación de los LAs entre Atta mexicana y Camponotus ocreatusEn las dos especies de hormigas estudiadas, tanto los tractos como los grupos de glomérulos mantienen la misma distribución topográfica (Figura 2), lo cual sugiere que la estructura general de los LAs se ha conservado y que debe tener alguna importancia en la vida social basada en el olfato, ya que otros insectos no-sociales pero olfativos13,44 no presentan una organización glomerular como la de los himenópteros. Sin embargo, A. mexicana tiene unos 80 glomérulos menos que C. ocreatus (cuyo LA tiene unos 380). Esto sugiere una mayor especialización de los soldados en el sistema de castas de A. mexicana en comparación con C. ocreatus. Es posible que el sistema de castas de A. mexicana produzca individuos más especializados y adaptados a funciones más específicas dentro de la sociedad. Esto es posible tomando en cuenta los conteos realizados para otras especies de hormigas de la tribu Attini como Acromyrmex octospinosus que presenta 120 glomérulos42; obreras minor de Atta vollenweideri que presentan 178, mientras que las obreras mayor presentan 243; obreras minor de Atta sexdens que presentan 223, y obreras mayor que presentan 18945. Por el contrario, los conteos para especies del género Camponotus indican en general un mayor número de glomérulos. Por ejemplo en Camponotus vagus hay 200 glomérulos42, en C. floridanus los soldados presentan un promedio de 45234, los soldados de C. sericeus 344, los de C. compresus 47640, en C. japonicus las obreras tienen 480, las reinas 470 y los machos 29041, mientras que las reinas de C. ocreatus tienen 400 y los machos 240 glomérulos32. Es posible que la especialización morfofisiológica del sistema de castas de Atta se refleje en las estructuras neurales que generan y controlan el comportamiento, como se aprecia en el reducido número de glomérulos en el LA. De hecho, las diferentes castas de hormigas Atta presentan diferentes umbrales de respuesta a olores particulares46, lo cual probablemente esté relacionado con el número y tipo de sensilas y de NRO presentes en las antenas, y, por lo tanto, con la estructura del LA y otras regiones del cerebro.
Debe considerarse que Atta mexicana y Camponotus ocreatus, son especies con formas de vida muy diferentes, ya que en la primera, el sistema de castas está basado en el polimorfismo como una adaptación al cultivo de hongos a partir de un sustrato vegetal, mientras que el género Camponotus, aunque presenta obreras mayor con enormes cabezas que son llamadas soldados y obreras de diferentes tamaños, es considerado generalista y dimórfico, más que polimórfico. Las hormigas Camponotus presentan división del trabajo basada en el polietismo temporal con individuos con desarrolladas capacidades de aprendizaje y memoria1,47 y que utilizan más la visión que las hormigas Atta, consideradas hormigas olfativas. Lo anterior no quiere decir que las hormigas Camponotus tengan atrofiado su sistema olfativo, sino que las Atta tendrían atrofiado su sistema visual. A primera vista esto parece contradecir los resultados de este trabajo ya que C. ocreatus presenta más glomérulos en el LA que A. mexicana. Sin embargo, es al contrario ya que cada casta de Atta estaría especializada en la realización de una o unas pocas tareas dentro del nido. Además, la clasificación de “hormigas visuales” versus “hormigas olfativas”, se refiere a la relación que existe entre las regiones del cerebro que procesan señales visuales u olfativas. De esta manera, en Camponotus, los lóbulos ópticos y la región del collar (que recibe aferencias visuales) de los CFs son grandes, mientras que en Atta, estas regiones están reducidas, pero los LAs y la región del labio (que recibe aferencias olfativas de los LAs) son relativamente más grandes43. Además, la población de glomérulos de los LAs de los soldados de A. mexicana es heterogénea, con la mayoría de los glomérulos pequeños comparados con los 3 macroglomérulos de los grupos G3 y G5 (Figura 2). En contraste, en los soldados de C. ocreatus existe una relativa uniformidad en los glomérulos del LA y sólo dos estructuras glomerulares más grandes.
Debido a que el neuropilo de los glomérulos está compuesto por las aferencias de las NRO (además de neuritas y axones de las neuronas centrales), es posible que el aparente tamaño reducido de la mayoría de los glomérulos se deba a la relativa baja densidad de aferencias de las NRO que responden a ciertos olores, es decir, estos glomérulos presentarían un grado de atrofia. Por el contrario, los macroglomérulos podrían presentar una mayor cantidad de aferencias provenientes de una población alta de NRO sensibles a olores casta-específicos presentes sobre la antena. Lo anterior puede estar relacionado con las diferentes proporciones de sensilas sobre las antenas en diferentes castas de hormigas4,5. Esto significaría que los macroglomérulos de las obreras mayor de A. mexicana podrían ser glomérulos especializados en el procesamiento de olores particulares, probablemente feromonas de alarma u otros olores importantes para los soldados48. De hecho, se ha encontrado un glomérulo agrandado en forrajeras de Atta vollenweideri, que participa en la detección de la feromona de rastro45. Por otro lado, en Camponotus obscuripes se ha encontrado un pequeño subgrupo de glomérulos en los LAs especializados en el procesamiento de feromonas de alarma49. Un sistema olfativo especializado y atrofiado en algunas castas, con la presencia de glomérulos relativamente grandes explicaría la presencia de “macroglomérulos” en la casta obrera de las hormigas. En Camponotus ocreatus también se han descrito macroglomérulos relacionados con el dimorfismo sexual. Los machos presentan una gran estructura macroglomerular en el G5, el cual podría estar especializado en el procesamiento de feromonas sexuales para el encuentro de las hembras durante el apareamiento31,32. Por su parte, las reinas también presentan un glomérulo agrandado, aunque no un macroglomérulo, en el G531,32. En C. japonicus también se ha descrito la presencia de macroglomérulos en los machos41.
Es posible que la vida social imponga restricciones y requerimientos a la maquinaria neural y al procesamiento de feromonas sociales. Las variaciones en la estructura de los LAs estarían relacionadas con las formas particulares de vida de ambas especies. Así, las hormigas Atta representan un extremo del gradiente de complejidad social con individuos que tienen sistemas olfativos desarrollados y especializados43,50. Las especies del género Atta constituyen sociedades altamente complejas en las que ciertas capacidades individuales podrían estar reducidas o atrofiadas, quizá como una consecuencia del extremo polimorfismo, en donde los individuos están especializados morfológica y fisiológicamente para construir una sociedad compleja a expensas de las capacidades generales de los individuos o pérdida de la individualidad51. De esta manera, la diferenciación neural de las castas de obreras no es uniforme, por lo que la presencia de estos macroglomérulos indicaría un desarrollo alométrico diferencial en el LA de las castas de A. mexicana, quizá regulado por genes involucrados en el crecimiento y sensibles a las condiciones sociales. Así pues, es muy posible que en la sociogénesis de cada especie haya evolucionado un patrón particular de diferenciación alométrica y especialización neural entre sus castas de obreras50.
Un mapa odotópico en los cuerpos fungiformesEn este trabajo se han descrito dos grupos principales de NP que dividen el LA en dos regiones, una anterior y una posterior. Los grupos de NP están segregados inervando grupos de glomérulos por separado y enviando axones por vías separadas, ya sea a través del TAP-m o del TAP-l, y conectándose con dos capas en el cáliz de los CFs. En estas capas está representada topográficamente la organización de las dos regiones eferentes del lóbulo antenal, estableciendo un mapa odotópico del LA en la región del labio de los CFs (Figuras 4–7), como ha sido descrito para Drosophila52,53,54. No obstante que se encontraron dos vías principales que conectan los LAs con dos capas principales en los CFs, podría haber aún una segregación más fina, como lo sugiere el número de capas encontradas en este mismo trabajo y en otros estudios22,28.
Este mapa odotópico caracterizado por la convergencia espacial, permitiría la integración de las señales olfativas. Las dos vías paralelas que conectan los LAs con los CFs (Figura 7) podrían representar dos canales de procesamiento de información olfativa proveniente de las NRO de las antenas, ya sea procesando diferentes clases de olores (procesamiento diferencial) o diferentes propiedades (procesamiento paralelo) de los mismos olores35. De hecho, se ha demostrado que los glomérulos que procesan olores de alarma están conectados por axones de NP uniglomerulares49 que van a través del TAP-m hacia la capa externa del cáliz, y por axones de NP multiglomerulares que se dirigen hacia el CL por el TAP-ml (por lo que provienen de los grupos de glomérulos de la región posterior del LA). De hecho, se ha reportado que la mayoría de las NP uniglomerulares que son sensibles a la feromona de alarma no responden a olores no-feromonales, mientras que muchas de las NP multiglomerulares sensibles a la feromona de alarma responden a olores no-feromonales49. Por ello, el procesamiento diferencial de olores no necesariamente excluye el procesamiento paralelo, ya que el primero podría llevarse a cabo en los CFs (pues la segregación separa grupos de glomérulos completos inervados por NP uniglomerulares, no glomérulos individuales), mientras que el segundo en el CL (que recibe axones multiglomerulares). No obstante, esto debe estudiarse en detalle con experimentos y registros de la actividad de las dos regiones eferentes del LA, de los dos tractos y de las capas del cáliz de los CFs.
Es posible que los olores sociales se procesen de manera separada por uno de estos canales, mientras que otro tipo de olores no-sociales sean procesados por el otro. De hecho, se ha sugerido que ésta podría ser una de las funciones principales de la vía olfativa dual en las hormigas55. Probablemente los dos canales procesen de manera separada amplias clases de olores y atributos de los estímulos relevantes conductualmente, y su posterior procesamiento convergente sea intergrado junto con otras modalidades sensoriales que proporcionarían modulación o información importante como el contexto o aspectos aprendidos. Así, esta organización funcional implicaría una alta especialización del cerebro de las hormigas a la vida social, tal y como lo sugiere la hipótesis de los cerebros sociales56. Las eferencias desde los LAs hacia los CFs no estarían segregadas solamente por modalidad sensorial, sino también en diferentes canales conductualmente relevantes de submodalidades que permitirían la funcion asociativa que se les ha adscrito a los CFs, tales como vincular modalidades sensoriales como un color con un olor22, o una feromona en un contexto específico. Esta segregación de las vías olfativas en paralelo en diversos grupos de insectos35 podría representar una adaptación a ambientes y formas de vida con demandas olfativas complejas, como la vida en sociedad que está basada en la detección y procesamiento de feromonas.
Es probable que las subpoblaciones de células Kenyon57,58 reciban aferencias de grupos particulares de glomérulos, y, por lo tanto, procesen diferente información olfativa, como ha sido descrito en Drosophila melanogaster y en Apis mellifera59,60,61, la cual sería posteriormente integrada junto con otras modalidades sensoriales. Si la codificación de los olores en el LA se basa en el patrón de actividad de diferentes glomérulos, el labio podría codificar de manera convergente y abstracta este patrón en actividad diferencial de las capas que lo constituyen. Esto podría jugar un papel importante en el aprendizaje y la memoria y en el procesamiento de información multimodal. Además, puesto que existen diferencias morfológicas entre especies en número y tamaño de glomérulos en los LAs, es posible que también existan diferencias especie-específicas tanto morfométricas, de arquitectura neural así como funcionales en la representación del LA en los CFs. Es posible que, en una especie, también existan diferencias casta-específicas en dicha representación, similares a las que se han descrito en las castas temporales de Pheidole dentata62. Estas diferencias en la organización del LA y su representación odotópica en los CFs estarían relacionadas con los estímulos que son importantes en el ambiente y organización social de cada especie. Por lo tanto, ya sea entre especies o entre castas de una misma sociedad, podrían existir diferencias identificables en la estructura y función de sus cerebros, ya que la evolución de los CFs en los himenópteros ha estado marcada por la expansión, el desarrollo y la especialización de los subcompartimientos de los cálices63.
No obstante que las similitudes interespecíficas en la vía olfativa encontradas en este trabajo sugieren una organización funcional altamente conservada en los himenópteros sociales, el significado de este mapa odotópico sólo puede ser explicado con futuros experimentos neuroanatómicos, morfométricos, funcionales y conductuales apropiados.
Este trabajo es parte de la tesis de doctorado del autor y fue financiada por CONACYT (158200). Agradezco al Dr. Wulfila Gronenberg por las facilidades otorgadas para el desarrollo de este trabajo durante la estancia doctoral realizada de octubre del 2001 a febrero de 2002 en la Universidad de Arizona, en Tucson, AZ, EUA. Los resultados de este artículo se presentaron en el Society for Neuroscience 34th Annual Meeting, San Diego California, USA, en el 2004.