



Tension cost, the ratio of myosin ATPase activity to tension, reflects the economy of tension development in the myocardium. To evaluate the mechanical advantage represented by the tension cost, we studied papillary muscle contractility and the activity of myosin ATPase in the left ventricles in normal and pathophysiological conditions.
METHODS:Experimental protocols were performed using rat left ventricles from: (1) streptozotocin-induced diabetic and control Wistar rats; (2) N-nitro-L-arginine methyl ester (L-NAME) hypertensive and untreated Wistar rats; (3) deoxycorticosterone acetate (DOCA) salt-treated, nephrectomized and salt- and DOCA-treated rats; (4) spontaneous hypertensive rats (SHR) and Wistar Kyoto (WKY) rats; (5) rats with myocardial infarction and sham-operated rats. The isometric force, tetanic tension, and the activity of myosin ATPase were measured.
RESULTS:The results obtained from infarcted, diabetic, and deoxycorticosterone acetate-salt-treated rats showed reductions in twitch and tetanic tension compared to the control and sham-operated groups. Twitch and tetanic tension increased in the N-nitro-L-arginine methyl ester-treated rats compared with the Wistar rats. Myosin ATPase activity was depressed in the infarcted, diabetic, and deoxycorticosterone acetate salt-treated rats compared with control and sham-operated rats and was increased in N-nitro-L-arginine methyl ester-treated rats. These parameters did not differ between SHR and WKY rats. In the studied conditions (e.g., post-myocardial infarction, deoxycorticosterone acetate salt-induced hypertension, chronic N-nitro-L-arginine methyl ester treatment, and streptozotocin-induced diabetes), a positive correlation between force or plateau tetanic tension and myosin ATPase activity was observed.
CONCLUSION:Our results suggest that the myocardium adapts to force generation by increasing or reducing the tension cost to maintain myocardial contractility with a better mechanical advantage.
Myocardial contraction results from a cyclic interaction between actin and myosin. In the heart, this mechanism adapts to several pathophysiological conditions, including pressure and volume overload, myocardial infarction, and diabetes, to maintain adequate cardiac output. Although a significant amount of data support a positive correlation between mechanical parameters, muscle-shortening velocity or force generation, and myosin ATPase activity (1–8), controversies still exist.
Mechanical activity in different situations, such as myocardial infarction, diabetes, or hypertension with and without hypertrophy, might present different correlations among the variables of force, the myosin isoforms involved and myosin ATPase activity. This indicates that several adaptive mechanisms might act to maintain myocardial contractility under these conditions. Particular changes have already been described for different physiopathological conditions. For example, a previous report described that after 12 weeks of myocardial infarction, papillary muscle force and contraction kinetics were enhanced, but the level of myosin ATPase activity was reduced (6). However, results from Wang et al. (9) showed a correlation between reduced contractility and a decrease in myosin ATPase activity.
Myocardial hypertrophy has been reported to be associated with reduced contraction speed, but changes in myosin ATPase activity may or may not be present (4,10,11). Diabetes is another condition that affects contractility by reducing force magnitude, increasing contraction duration (3,12,13), and producing a shift from myosin V1 to V3 (12). In L-NAME-treated rats, the force of muscle contraction can be increased without affecting the myosin isoform composition of the cardiac muscle (7,14). Moreover, force generation can either increase or remain unchanged with myosin isoform shifts and changes in the level of myosin ATPase activity. These findings show the multiplicity of the adaptive mechanisms involved in myocardial contractility regulation, in both normal and pathophysiological conditions.
To minimize the cause of the aforementioned controversies, we used a previously suggested approach for the analysis of tension cost: the ratio of myosin ATPase activity to tension (15–17). This ratio reflects the economy of tension development and can be used to predict mechanical changes corresponding to a biochemical change. Therefore, experiments were performed in several models (e.g., myocardial infarction, deoxycorticosterone acetate (DOCA)-salt-induced hypertension, L-NAME-induced hypertension, experimental diabetes) and in SHRs under stabilized conditions. The level of myosin ATPase activity was correlated with the force generation capacity and the tension cost for each condition.
MATERIALS AND METHODSMale Wistar rats weighing 250 to 350 g, Wistar Kyoto (WKY) rats weighing 230 to 350 g, and spontaneous hypertensive rats (SHR) weighing 200 to 300 g were used in this study. All experiments were conducted in compliance with the guidelines for biomedical research as stated by the Brazilian Societies of Experimental Biology. All rats had free access to water and were fed rat chow ad libitum.
The rats were divided into 12 groups for the mechanical and biochemical studies (Table 1). For the mechanical studies, the groups included rats with myocardial infarction (n = 21) and sham-operated rats (n = 23); diabetic rats (n = 12) and controls (n = 12); DOCA-salt-treated (n = 10), nephrectomized rats (n = 12), salt-treated (n = 12) and DOCA-treated rats (n = 15); L-NAME-treated (n = 7) and untreated (n = 12) rats; and SHR rats (n = 22) and Wistar Kyoto rats (WKY, n = 10) as the controls (Table 1). For the biochemical studies, other groups of rats were used, including infarction (n = 7) and sham-operated (n = 7); diabetes (n = 7) and control (n = 6); DOCA-salt-treated (n = 6), salt-treated (n = 6), DOCA-treated (n = 6) and nephrectomized (n = 6); L-NAME (n = 9) and control (n = 10); and SHR (n = 8) and WKY (n = 8).
Diabetes inductionDiabetes was induced with streptozotocin (50 mg/kg body weight diluted in 0.1 M citrate buffer solution, pH 4.5, i.v. single injection). Two main groups of rats were used: control rats and rats treated with streptozotocin for 30 days. Blood glucose was measured using a hemoglucotest (Advantage, Roche) before and after streptozotocin treatment. Animals that did not develop hyperglycemia were excluded. The blood pressure of each animal was measured in vivo using a tail cuff (IITC Model 29 Pulse Amplifier, IITC Inc., California).
L-NAME-induced hypertensionRats were divided in two main groups. To induce hypertension, one group was treated for 30 days with L-NAME (N-nitro-L-arginine methyl ester). L-NAME (50 mg/kg/day) was administered in the drinking water. Control rats received untreated water. The day before the experiments were conducted, the systolic blood pressure of the rats was measured using a tail cuff (IITC Model 29 Pulse Amplifier, IITC Inc., California).
DOCA-salt-induced hypertensionTo induce hypertension with DOCA-salt, male 45-day-old rats were uninephrectomized under halothane anesthesia, as previously described (18). Four days later, the rats were treated with either deoxycorticosterone acetate (DOCA, 8 mg/kg twice a week) or vehicle (soybean oil, 0.1 mL/animal). This treatment was repeated twice per week for 30 days. The uninephrectomized rats received drinking water as vehicle, and the DOCA-salt-treated rats received a saline solution (0.9% NaCl) as their drinking water during the treatment period. The day before the experiments were conducted, the systolic blood pressure of all rats was measured using a tail cuff (IITC Model 29 Pulse Amplifier, IITC Inc., California). Animals that did not develop hypertension were excluded.
Myocardial infarction modelMyocardial infarction was induced by ligation of the left coronary artery as previously described (19). Anesthesia was induced by halothane. After recovery, the rats were kept in collective cages at the animal facility. To avoid stress and because we had previous measurements of systolic blood pressure in a similar group of rats, systolic blood pressure was not measured in the tail.
Studies of myocardial contractilityThe rats received 500 units of heparin i.p., and after 10 min, they were anesthetized with 45 mg/kg of sodium pentobarbital i.p. The hearts were rapidly removed after thoracotomy and perfused with Krebs solution through the aortic stump to permit proper selection and dissection of the left ventricle papillary muscles. The papillary muscles were immersed in a 50-mL water-jacketed bath maintained at 29±1°C and gassed with (95% O2 and 5% CO2) Krebs bicarbonate buffer solution. The muscles were stretched to Lmax (muscle length at which active tension is maximal) and stimulated with isolated rectangular pulses (10 to 15 V, 12-ms duration) using a pair of platinum electrodes placed along the entire length of the muscle. The standard stimulation rate was 0.5 Hz (steady-state). Recording was begun after 60 minutes to allow for the beating preparation to adapt to the new environmental conditions.
The bathing solution was a modified Krebs solution with the following composition (in mM): 120 NaCl, 5.4 KCl, 1.2 MgCl2 6H2O, 1.25 CaCl2 2H2O, 2.0 NaH2PO4H2O, 1.2 Na2SO4, 27 NaHCO3, and 11 glucose, gassed with 95% O2 and 5% CO2, pH 7.4.
The developed force (F) was measured with an isometric force transducer (Nihon-Kohden, TB 612T, Tokyo), which was recorded on a chart recorder (Nihon-Kohden, RM-6200, Tokyo) and normalized to the muscle cross-sectional area (g/mm2). Considering the papillary muscle as a cylinder and the tissue density as 1, the cross-sectional area was calculated by dividing the muscle length at Lmax by its weight. To avoid the possibility of a hypoxic core, we performed the experiments at low temperatures (29±1°C) as previously described (20).
The influence on the contractile machinery was studied in another group of papillary muscles during tetanic stimulation. Tetanic tension was obtained after treatment with 5 mM caffeine (for 20 minutes, at a rate of 10 Hz, for 15 s) as previously described (20). The tetanic contractions developed a fast upstroke (tetanic peak force) followed by a slow decay (tetanic plateau force). Tension was measured at the peak and in the middle of the plateau. The muscles remained under steady-state stimulation for at least 10 min between stimuli. This protocol was repeated twice.
Studies of myosin ATPase activityTo evaluate myosin ATPase activity, the left ventricle was dissected and assayed as described previously (11). Briefly, myosin was prepared from minced and homogenized left ventricles, which were extracted briefly with KCl phosphate buffer (0.3 M KCl and 0.2 M phosphate buffer, pH 6.5).
Myosin ATPase activity was assayed by measuring Pi liberation from 1 mM ATP in the presence of 50 mM HEPES, pH 7, 0.6 M KCl, and 5 mM CaCl2 or 10 mM EGTA in a final volume of 200 µl. The level of myosin activity was measured without Mg2+ in the medium. There is no significant Ca2+-ATPase activity in the sarcoplasmic reticulum membrane, which requires high Mg2+ and low Ca2+ concentrations. Controls containing the enzyme preparation and trichloroacetic acid were used to correct for non-enzymatic hydrolysis of the substrate. All samples were prepared in duplicate. The level of enzyme activity was calculated as the difference between the levels observed in the presence of Ca2+ and in the presence of 10 mM EGTA. The level of inorganic phosphate was determined according to the method proposed by Chan et al. (21). Specific activity is reported as nmol Pi released per min per mg of protein unless otherwise stated. Protein was measured using the Coomassie Blue method according to Bradford (22) using bovine serum albumin as a standard.
Drugs usedPentobarbital sodium was obtained from Cristalia Produtos Químicos Farmacêuticos Ltda. (São Paulo, SP, Brazil). Heparin was obtained from Roche Q.F.S.A. (Rio de Janeiro, RJ, Brazil) and Sigma. Bovine serum albumin, Na2ATP, and anhydrous caffeine were purchased from Sigma Chemical Co. (St. Louis, MO, U.S.A.). All of the other reagents used were of analytical grade and from Sigma, E. Merck (Darmstadt, Germany), or Reagen (Rio de Janeiro, RJ, Brazil).
Statistical analysisThe results are presented as the means ± SEM with n indicating the number of observations. The values were analyzed using ANOVA (one- and two-way) and unpaired Student t-tests. When ANOVA revealed a significant difference, the Tukey test was applied. Linear regression was used for correlation analysis. p<0.05 was considered to be significant. Data analysis and figure preparation were performed using GraphPad PrismTM (version 2.0, GraphPad Software, San Diego, CA, USA) and GB-STAT (version 4.0, Dynamic Microsystem Inc., Silver Spring, MD).
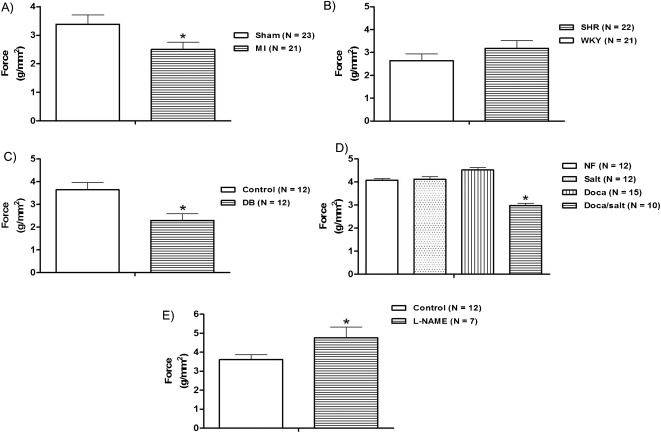
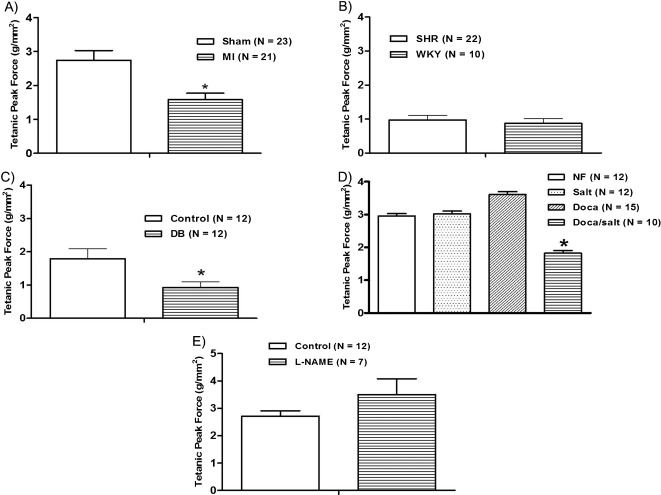
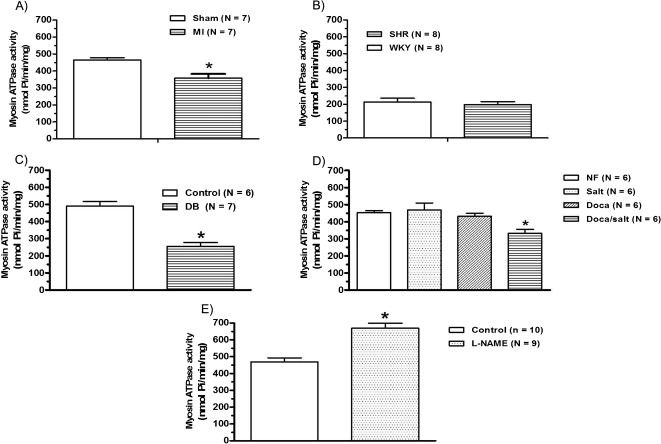
RESULTSFour weeks after myocardial infarction, the isometric force development (Figure 1A) and peak and plateau tetanic tension (Figure 2A), Table 3 were reduced. The level of myosin ATPase activity was also reduced compared to that observed in sham-operated rats (Figure 3A).
, hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on the isometric force (g/mm2) in the rat papillary muscle. The results represent the mean ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control, Sham, NF, Salt, and DOCA (t-test or one-way ANOVA, repeated measurements).")
Effects of myocardial infarction (A), hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on the isometric force (g/mm2) in the rat papillary muscle. The results represent the mean ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control, Sham, NF, Salt, and DOCA (t-test or one-way ANOVA, repeated measurements).
, hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on the tetanic peak force (g/mm2) in rat papillary muscle. The results represent the mean ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control, Sham, NF, Salt, and DOCA (t-test or one-way ANOVA, repeated measurements).")
Effects of myocardial infarction (A), hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on the tetanic peak force (g/mm2) in rat papillary muscle. The results represent the mean ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control, Sham, NF, Salt, and DOCA (t-test or one-way ANOVA, repeated measurements).
Changes in plateau tetanic tension (g/mm2) as observed in papillary muscles obtained from myocardial infarction (MI) and sham-operated (Sham) rats; spontaneously hypertensive rats (SHR) and Wistar Kyoto (WKY), diabetic; nephrectomized (Nefrec), salt-treated (salt), DOCA-treated, DOCA-salt-treated (DOCA), and L-NAME-treated rats.
| Groups | Plateau Tetanic Tension (g/mm2) |
|---|---|
| Sham (23)MI (21) | 1.77±0.220.80±0.10∗ |
| WKY (10)SHR (22) | 0.87±0.14∗0.97±0.13∗ |
| Control (12)Diabetic (12) | 1.79±0.300.92±0.17∗ |
| Nefrec (12)Salt (12)DOCA (15)DOCA-salt (10) | 1.97±0.142.08±0.232.20±0.251.06±0.25∗ |
| Control (12)L-NAME (7) | 1.71±0.182.30±0.42 |
Values represent the means ± SEM. ANOVA one-way and t-test: ∗p<0.05 vs. controls of respective groups. The number of animals is indicated in parentheses.
, hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on myosin ATPase activity (nmol/Pi/min/mg) in the rat left ventricle. The results represent the means ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control (t-test or one-way ANOVA, repeated measurements).")
Effects of myocardial infarction (A), hypertension (B), diabetes (C), DOCA-salt-induced hypertension (D), and chronic L-NAME treatment (E) on myosin ATPase activity (nmol/Pi/min/mg) in the rat left ventricle. The results represent the means ± SEM, and N indicates the number of experiments. ∗p<0.05 vs. control (t-test or one-way ANOVA, repeated measurements).
SHRs were hypertensive (systolic pressure; SHR = 213±7.5 mmHg vs. WKY = 124±1.3 mmHg; p<0.01, unpaired Student's t-test) and showed left ventricular hypertrophy (Table 2). Force development and peak and plateau tetanic tension were similar between SHRs and WKY rats (Figure 1B) and 2B). For SHRs and WKY rats, this similarity in terms of tetanic tension was paralleled by similar levels of myosin ATPase activity (Figure 3B).
Measurements of final body weight (g), left ventricular weight (mg), and the left ventricular weight/body weight (LVW/BW) ratio (mg/g) in myocardial infarction (MI) and sham-operated (Sham) rats; spontaneously hypertensive rats (SHR) and Wistar Kyoto (WKY), diabetic; nephrectomized (Nefrec), salt-treated (salt), DOCA-treated, DOCA-Salt (DOCA), and L-NAME-treated rats.
| Groups | Final BodyWeight (g) | Left VentricularWeight (mg) | LVW/BW ratio(mg/g) |
|---|---|---|---|
| Sham (23)MI (21) | 347±9.9331±7.9 | 660±20670±40 | 1.90±0.052.04±0.13 |
| WKY (10)SHR (22) | 296±7.6251±7.8∗) | 600±20700±30∗) | 2.05±0.042.82±0.09∗) |
| Control (12)Diabetic (12) | 289±4.2262±3.9∗) | 550±20552±10 | 1.91±0.061.99±0.03 |
| Nefrec (12)Salt (12)DOCA (15)DOCA-Salt (10) | 285±6.5288±8.7289±8.0249±7.7∗) | 540±10570±20600±20790±30∗) | 1.89±0.041.96±0.042.07±0.033.19±0.16∗) |
| Control (12)L-NAME (7) | 340±7.8319±12.7 | 670±20660±20 | 1.97±0.052.08±0.05 |
Values represent the means ± SEM. ANOVA one-way and t-test:
Thirty days after streptozotocin administration, the diabetic group showed an increased blood glucose level (DB = 488±20.5 mg/dL and Control = 82±1.8 mg/dL, p<0.01, unpaired Student's t-test) and a reduction in body weight (Table 2) when compared to baseline (Control: 270±3.16 vs Diabetic: 280±3.63, p>0.05). The papillary muscles from diabetic rats developed less isometric tension than those from controls (Figure 1C); the same trend was observed for peak and plateau tetanic tension (Figure 2C), Table 3). The level of myosin ATPase activity was reduced (Figure 3C). In the DOCA-salt group, only the DOCA salt-treated rats were hypertensive. The other three subgroups (nephrectomized, salt treated, and DOCA treated) were normotensive (DOCA salt = 201±3.4 mmHg vs. Nefrec = 119±1.1 mmHg; Salt = 120±2.1 mmHg and DOCA = 135±2.0 mmHg: p<0.01, one-way randomized ANOVA). DOCA-salt rats also presented reduced body weight compared to the other three subgroups and displayed significant LV hypertrophy at the end of the treatment (Table 2). Isometric tension (Figure 1D) and peak and plateau tetanic tension (Figure 2D), Table 3 were reduced in the DOCA-salt group compared with the other groups. Myosin ATPase activity was also reduced in the DOCA-salt group (Figure 3D).
We used L-NAME treatment for 30 days to create a model with hypertension but without LV hypertrophy. Arterial systolic pressure increased (Control = 119±0.8 mmHg and L-NAME = 196±4.4 mmHg; p<0.01, unpaired Student t-test), but no hypertrophy developed (Table 2). L-NAME-treated rats developed increased isometric force (Figure 1E), but peak and plateau tetanic tension presented no significant differences (Figure 2E), Table 3. Myosin ATPase activity increased in the L-NAME group (Figure 3E).
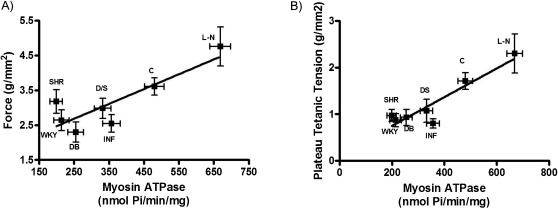
Because our results were consistent with the previously reported controversies, we attempted to identify a correlation between mechanical changes and biochemical changes. According to Rundell et al. (17), the relationship between myosin ATPase activity and developed force provides information about the energetic cost of tension. As shown in Figure 4), in which force is plotted against myosin ATPase activity (tension cost) observed in the L-NAME, DOCA-salt, diabetic, Wistar control, SHR and WKY groups, there was a positive correlation among these variables under both normal conditions (Figure 4A) and with the use of tetanic plateau force (Figure 4B).
 the isometric force (g/mm2) or (B) the plateau tetanic tension and the myosin ATPase activity (nmol/Pi/min/mg) of the left ventricle in control, myocardial infarction, diabetic, DOCA-salt, L-NAME-treated, SHR, and WKY rats. The results represent the means ± SEM.")
Relationship between (A) the isometric force (g/mm2) or (B) the plateau tetanic tension and the myosin ATPase activity (nmol/Pi/min/mg) of the left ventricle in control, myocardial infarction, diabetic, DOCA-salt, L-NAME-treated, SHR, and WKY rats. The results represent the means ± SEM.
Our results demonstrate that tension cost, the ratio of myosin ATPase activity to force, correlates mechanical and corresponding biochemical changes. The ratio shows that the contractile state of the myocardium in several conditions, such as post-myocardial infarction, DOCA-salt-induced hypertension, chronic L-NAME treatment, and streptozotocin-induced diabetes, is represented by a positive correlation between tension developed and myosin ATPase activity. This characterizes the energetic cost of tension as an attempt to obtain a better mechanical advantage.
The relationship between myosin ATPase activity and contraction velocity is well established (1). In the rat myocardium, however, the activity and expression of myosin ATPase has been suggested to reflect myocardial force development (2–7). The three myosin isoforms are denoted as V1, V2, and V3. The V1 isoform (αα) possesses the highest ATP hydrolysis velocity of the three and composes the heavy chains. The isoform displaying the lowest velocity is V3 (ββ); V2 (αβ) is a mixed isoform with intermediate velocity (5,8,10). However, during adaptation to several pathophysiological conditions, Ca2+-handling mechanisms might prevent mechanical deficiencies before stable conditions are achieved. These various adaptive mechanisms explain why correlations between force development and the biochemical activity in cardiac muscle are still a matter of debate. We then performed experiments with several models that exhibit changes in myosin ATPase activity and force when conditions stabilize.
Myocardial infarction is associated with adaptive changes that enable the heart to maintain an effective systolic performance (23). The infarcted chamber increases its volume and the end-diastolic pressure to maintain cardiac output. These changes are limited by the contractile capacity of the remaining myocardium and the energetic cost of the mechanical activity. Our results obtained 30 days after myocardial infarction show a reduction in isometric and tetanic tension. These findings and those regarding the area of infarction are similar to previous results from our laboratory (23–25). However, the mechanisms underlying this force reduction remain to be determined. Min et al. (26) reported reductions in force and SERCA2 levels. Hyposensitivity to positive inotropic stimulation with calcium and isoproterenol was also reported (19). These findings are paralleled by the reduction in myosin ATPase activity reported here. Other authors have reported similar findings (27,28), although without the shift of V1 to V3 MHC (28). Wang et al. (9) investigated the time course of myosin ATPase activity after myocardial infarction and found that the level of left ventricular myosin ATPase activity began to decrease at four weeks. These different adaptive changes might result from a phenomenon recently described in rats such that 30-day myocardial infarctions with similar scar areas can behave normally or, alternately, lead to the development of heart failure (25).
SHRs develop hypertension and myocardial hypertrophy. These rats have a large period of stable hypertrophy with preserved or enhanced contractile function (23,29). In fact, we did not find differences in force development in these rats compared to WKY rats. Previous reports suggest an increase in contractile force resulting from either increased Ca2+ entry through L-type Ca2+ channels and release from the sarcoplasmic reticulum (29) or enhanced contractility without changes in the levels of contractile proteins but with large calcium spikes (30). However, other reports described reductions in force under steady-state or isoproterenol stimulation (31) resulting from increased calcium uptake by the sarcoplasmic reticulum and an increase in the level of phospholamban phosphorylation. Taken together, these results show that the development of force and the nature of calcium handling change during the evolution of hypertension in SHRs. Our results, however, showed no differences in either the development of force or the level of myosin ATPase activity, suggesting that we studied SHRs with stable contractility function, which explains why the SHRs displayed tension cost similar to that of WKY rats.
Regarding the development of tetanic tension, our results demonstrate that under tetanic stimulation, the isometric tension force was not modified in SHRs compared with WKY rats. In accordance with these results, the level of myosin ATPase activity in SHRs was similar to that in WKY rats. Other reports show a reduction in myosin ATPase activity but only after longer periods (32,33).
Diabetes is a chronic syndrome resulting from insulin deficiency, which also affects the cardiovascular system. Cardiac contractility dysfunction is described in streptozotocin-induced diabetes in rats. Notably, our results are in accordance with those showing isometric force reduction reported by Fein et al. (12) and Dhalla et al. (3). Other reports showed no alteration in force alterations (17,34) but differ from our study because myocardial hypertrophy was present in their preparations. Regarding evaluation of the contractile proteins, our results show a reduction in tetanic tension and a decrease in myosin ATPase activity. These findings are in accordance with previous reports (17,35).
The DOCA-salt rat exhibited cardiac hypertrophy after 30 days as previously reported (23). However, the reports regarding changes in contractility are controversial (36,37). Our findings show that DOCA-salt-treated papillary muscles developed less tension, but other studies show no change or even increased tension (36,37). However, all related studies report cardiac hypertrophy. Tetanic tension and myosin ATPase activity were also reduced, suggesting the involvement of contractile proteins.
L-NAME treatment induces severe hypertension but little or no hypertrophy. Our findings show no hypertrophy and are in accordance with previous reports (7,14). Meanwhile, other reports show hypertrophy (38) or hypotrophy (39). The increased isometric force development observed in the present study is in agreement with the results reported by Bartunek et al. (14), which described an increase in contractility indexes. However, we observed no changes in tetanic tension development, although the level of myosin ATPase activity was found to be increased.
Therefore, our results, in agreement with previous reports, demonstrate that tension cost is the best way to correlate mechanical and biochemical parameters (15–17),. We used this approach to maintain consistency, and our results are in accordance with those reports. Interestingly, as shown in Figure 4, once the sarcoplasmic reticulum activity is reduced, the tension cost (myosin ATPase activity vs. plateau tetanic tension) reveals a clear correlation, suggesting that any existing discrepancies may be due to the interference of sarcoplasmic reticulum adaptation mechanisms.
Our results suggest that higher myosin ATPase activity leads to a higher energy cost in association with the development of tension. When myosin ATPase activity in the heart is reduced, there is an accompanying reduction in tension cost. Based on these findings, we suggest that the myocardium increases or decreases tension cost to maintain myocardial contractility with a better mechanical advantage.
FUNDING
This work was supported by CNPq and FAPESP/FUNCITEC.
AUTHOR CONTRIBUTIONSMoreira CM performed the experimental protocols, statistical analyses, and wrote and reviewed the manuscript. Meira EF and Vestena L performed the experiments. Stefanon I, Vassallo DV and Padilha AS wrote and reviewed the manuscript. All the authors have read and approved the contents of the manuscript. Each author has made a valuable contribution to the preparation of this manuscript.
No potential conflict of interest was reported.