





INTRODUCCIÓN
Entre los numerosos datos que apoyan el concepto de que, efectivamente, las mitocondrias tienen un papel clave en el envejecimiento celular figuran los resultados obtenidos recientemente en dos especies de animales transgénicos. En efecto, según Fridell et al1, Drosophila adultas que expresan la proteína humana hUCP2 (causante de «uncoupling» mitocondrial en su sistema nervioso, con el consiguiente descenso en la producción de ROS) muestran un aumento de su longevidad con respecto a las moscas control. Por otra parte, Schriner et al2 han conseguido aumentar la longevidad media y máxima de ratones transgénicos que muestran altos valores de actividad de la catalasa humana en sus mitocondrias, mientras que los efectos en la longevidad no son tan marcados cuando el aumento de catalasa tiene lugar en el núcleo o los peroxisomas.
Concepto integrador del envejecimiento a los principales niveles de organización biológica
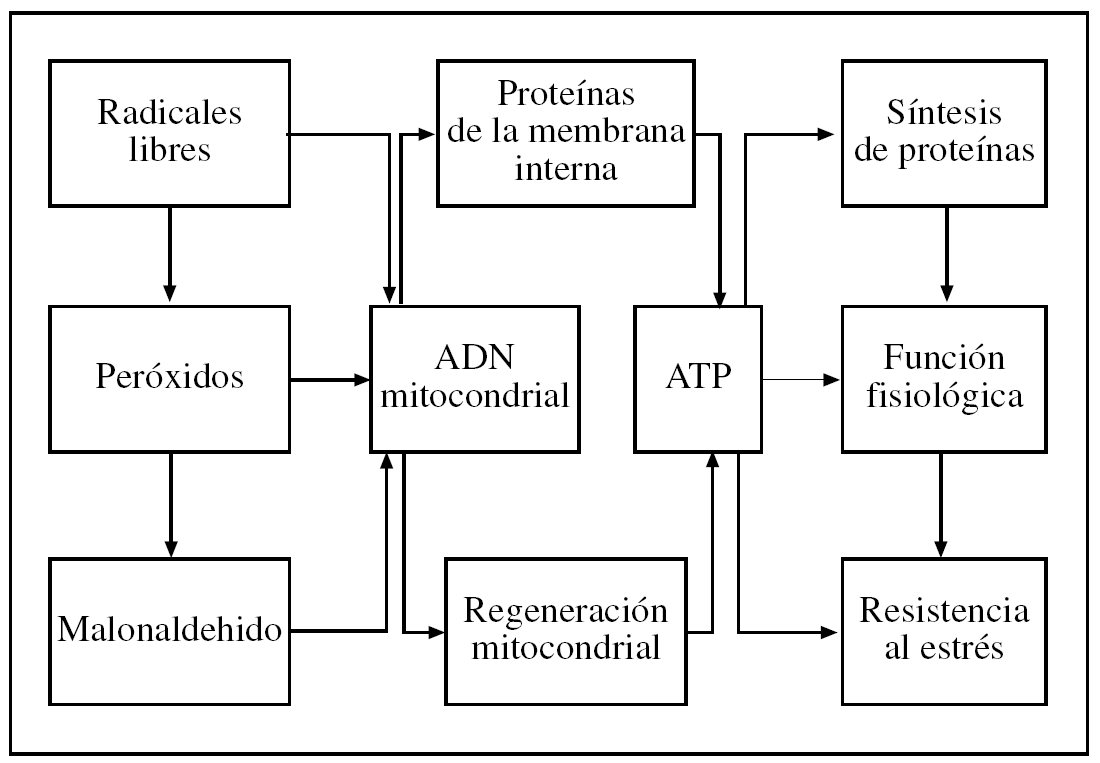
Según Dobzhanski3: «Algunos científicos se aferran a la fe reduccionista de que la manera de saberlo todo es concentrarse en la investigación del nivel (de organización) más bajo, al que consecuentemente se llama el nivel fundamental. Una estrategia más juiciosa y acertada de investigación científica es llegar a comprender los fenómenos (...) a todos los niveles de organización». Así, como ya hemos indicado, la aplicación de la metodología del análisis de sistemas4,5 puede ser útil para estudiar las relaciones de causa/efecto en los cambios que tienen lugar al envejecer en los diversos niveles de organización biológica (tabla 1), y así ayudar a integrar varias teorías complementarias que incluyen las más recientes, así como las mencionadas en la tabla 1 de la parte I de esta revisión. En la figura 1 se expone los dos conceptos clave de nuestra teoría integradora, es decir, el agente agresor es el estrés oxidativo intramitocondrial y la principal diana de la agresión es el ácido desoxirribonucleico (ADN) mitocondrial, con la consiguiente pérdida de mitocondrias funcionales y rendimiento bioenergético y fisiológico a los niveles celular y fisiológico.

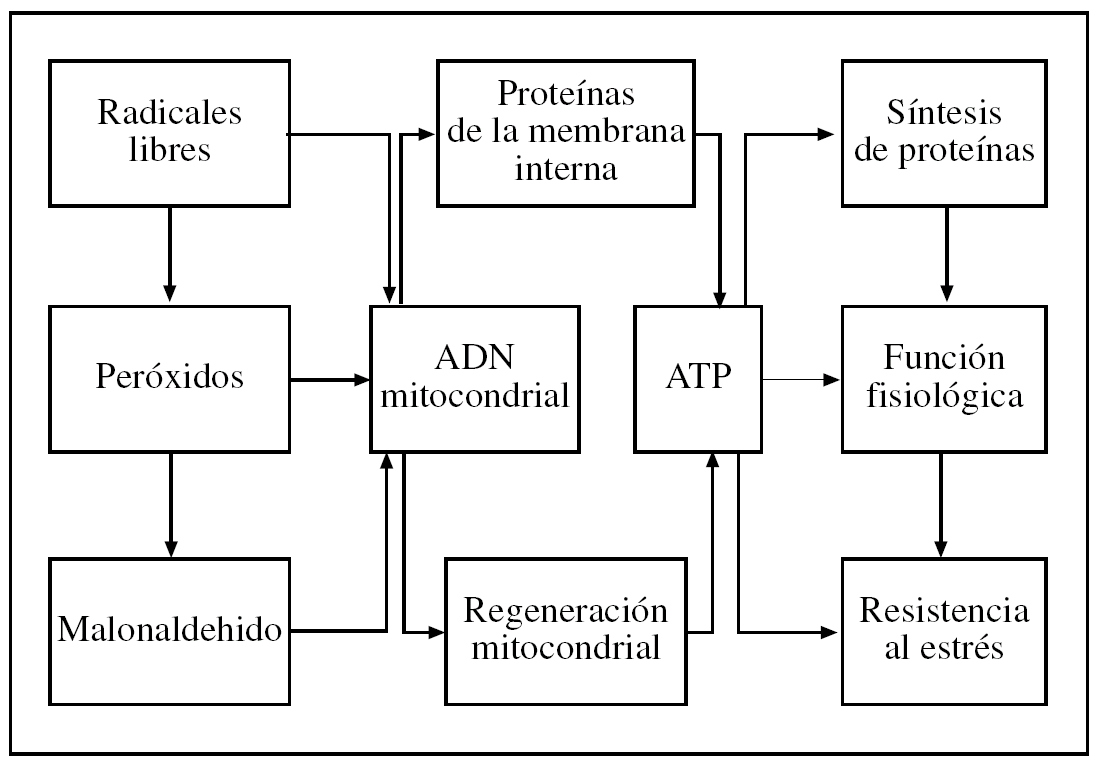
Figura 1.Este esquema propuesto por nuestro laboratorio (que incluye diversos mecanismos de senescencia ligados en una cadena de causas y efectos) es compatible con las teorías de los radicales libres, diferenciación celular, «desgaste» y «rate-of-living», así como con nuestra «teoría mitocondrial del envejecimiento de las células diferenciadas» y la teoría del «soma desechable». Modificado de Miquel y Fleming17. ADN: ácido desoxirribonucleico; ATP: adenosintrifosfato.
De acuerdo con lo anterior, los datos de numerosos estudios experimentales apoyan la teoría del rate-of-living de Pearl6, que define el proceso de envejecimiento como «desgaste» causado por el trabajo fisiológico, así como las teorías de Harman7 y Gerschman8, que asocian el envejecimiento con la acción de los radicales libres formados en el metabolismo y con la «toxicidad residual del oxígeno», respectivamente. Otras piezas clave, a nivel celular, de nuestra integración son el concepto de Weissmann9, que explica la pérdida de la supervivencia ilimitada de los metazoos como el resultado de la división del trabajo entre las células «inmortales» de los epitelios germinales y las células somáticas destinadas a envejecer, así como la afirmación de Minot10 de que el envejecimiento es «el precio pagado por la diferenciación celular». Estos conceptos también son compatibles con la teoría del soma desechable de Kirkwood y Holliday11, según la cual la longevidad de las distintas especies animales depende de un equilibrio entre su capacidad reproductora y la eficacia de sus mecanismos de mantenimiento y reparación de las células diferenciadas del soma, destinadas a «desecharse» una vez se ha asegurado la supervivencia de la especie mediante la reproducción (Kirkwood12). Y a los niveles subcelular y genético, la teoría mitocondrial del laboratorio del presente autor (Miquel et al13) puede contribuir a una integración más completa de las teorías del envejecimiento y señalar la importancia del genoma mitocondrial de las células diferenciadas como el probable «talón de Aquiles» del organismo multicelular.
Evolución biológica y origen del envejecimiento
En cuanto a la cuestión gerontológica clave, o sea las causas de la aparición del proceso de envejecimiento en la biosfera, este proceso probablemente deriva de una línea de evolución biológica que llevó a un predominio de la síntesis de adenosintrifosfato (ATP) mediante la vía aerobia, a expensas del más primitivo (y menos eficiente) metabolismo anaerobio. En una primera etapa de la evolución, en la que había abundante materia orgánica en la Tierra y la atmósfera contenía poco oxígeno, la glucólisis anaerobia bastaba para mantener la vida primitiva de las procariotas. Sin embargo, posteriormente los organismos tuvieron que hacer frente a una disminución en las fuentes orgánicas de energía y a un aumento simultáneo del contenido atmosférico de oxígeno. Esto condujo a la aparición del metabolismo aerobio y de células dotadas de mitocondrias, con un gran aumento en la producción de ATP. Sin embargo, un alto consumo de oxígeno es incompatible con una mitosis normal y por ello los organismos multicelulares deben mantener su respiración mitocondrial en valores relativamente bajos en sus líneas germinales. Así se preserva su capacidad mitótica y disminuye el riesgo de alteraciones en la información genética, cuya integridad es esencial para la reproducción de la especie. Por el contrario, las altas cuotas de síntesis de ATP (y respiración mitocondrial) que requieren las células somáticas para realizar su trabajo fisiológico especializado las lleva a su desorganización y envejecimiento a causa del estrés oxidativo mitocondrial. Según ya se ha comentado (Miquel14), el envejecimiento mitocondrial no supone un fallo de la evolución biológica. Ciertamente, la evolución podría haber suministrado mecanismos antioxidantes capaces de proteger al 100% contra la desorganización, que es el «efecto secundario» de los altos valores de síntesis de ATP y correspondiente consumo de oxígeno que requieren las células diferenciadas. En teoría, estas células podrían regenerar sus poblaciones mitocondriales mediante una mayor replicación de las mitocondrias supervivientes para compensar la pérdida de mitocondrias funcionales causadas por las ROS. Pero conviene recordar que las células diferenciadas tienen que distribuir la limitada cantidad de ATP que obtienen en las mitocondrias entre el mantenimiento de las propias células (incluidas sus mitocondrias) y las funciones que estas células realizan en apoyo del organismo del que forman parte. Por ello, para que una célula diferenciada, como la neurona, pudiera desviar más energía para reparar y mantener intacta su dotación de mitocondrias, tendría que sustraer parte del ATP que se destina al apoyo de las funciones especializadas que son esenciales para mantener al organismo en un estado de máxima capacidad funcional. Esto llevaría a una menor resistencia del organismo contra el estrés del hábitat (incluido el estrés oxidativo) y a un descenso de la capacidad reproductora, e incluso podría ser incompatible con la supervivencia de la especie. El gran número de especies de metazoos que existe prueba que la estrategia evolucionista que lleva al envejecimiento es adecuada, pues el desgaste de los tejidos especializados, aunque incompatible con la supervivencia del «soma desechable»15,16, suele ocurrir cuando la supervivencia de la especie se ha asegurado mediante la reproducción sexual.
Correspondencia: Dr. J. Miquel.
Departamento de Biotecnología. Universidad de Alicante.
Marqués de Campo, 66. 03700 Denia. Alicante. España.
Correo electrónico: kiri_miquel@yahoo.com
Recibido el 22-06-05; aceptado el 30-09-05.
