




La obesidad se caracteriza por un aumento de la masa adiposa secundaria a un balance energético positivo mantenido en el tiempo. El incremento en el volumen del tejido adiposo se acompaña de otros cambios en las características biológicas habituales de éste, que se vuelve disfuncional. El depósito visceral de la grasa, la hipertrofia y cambio del perfil secretor de los adipocitos, junto con la infiltración del tejido adiposo por células inflamatorias son algunas de las características que determinan una comunicación alterada del tejido adiposo con otros órganos. Se ha planteado que la disfunción del tejido adiposo, explicaría parte de la etiopatogenia de las enfermedades metabólicas y cardiovasculares asociadas a obesidad; sujetos obesos que mantienen un tejido adiposo funcional no presentan las alteraciones metabólicas propias de la malnutrición por exceso. La modulación de las características biológicas del tejido adiposo permitiría tener un menor riesgo cardiovascular asociado a obesidad.
Obesity is characterized by an increase in the adipose mass, which is due to the deposit of the fatty acids surplus in adipose cells, due to a positive energy balance maintained over the time. Along with the increase in the volume of adipose tissue, there are changes in its normal physiology, driving it toward a dysfunctional tissue. Such changes involve adipocyte hypertrophy and changes in its secretory profile, visceral deposit of fat, increased immune cell infiltration, among others. Adipose tissue dysfunction possibly plays a pivotal role in the pathophysiology of obesity-related metabolic and cardiovascular diseases. The understanding of these changes in adipose tissue function could provide a tool for modulating the metabolic and cardiovascular burden of obesity.
La obesidad es considerada en la actualidad una epidemia a nivel mundial. La Organización Mundial de la Salud (oms) estima que 1.500 millones de adultos presentaban sobrepeso u obesidad en el año 2008. Esto sucede tanto en países desarrollados como en vías de desarrollo. La obesidad es un problema en aumento no solo en términos de prevalencia; la mayor intensidad de presentación, así como el inicio a edades cada vez más tempranas, constituyen un incremento adicional del riesgo asociado. En Chile, las características de la transición epidemiológica permitieron que se cambiaran rápidamente los problemas de malnutrición por déficit por los problemas de malnutrición por exceso (1). Los datos nacionales derivados de la monitorización del estado nutricional en mujeres embarazadas y en niños que ingresan a primero básico permiten visualizar el aumento constante de la malnutrición por exceso en los últimos 30 años (2). Durante la última década se han generado datos representativos de la población chilena mayor de 15 años (3, 4). En esos estudios se refleja que más de 2/3 de la población tiene sobrepeso u obesidad, cifra que ha ido en aumento, a pesar de la cercanía temporal de las dos mediciones. En el caso de los adultos mayores (65 años y más) existe un 42,6% de sobrepeso y un 30,9% de obesidad.
La obesidad es un importante factor de riesgo para el desarrollo de múltiples patologías. Las comorbilidades más estudiadas de la obesidad son las metabólicas (resistencia insulínica, dislipidemia, diabetes mellitus tipo 2) y cardiovasculares (hipertensión, ateromatosis, hipertrofia ventricular izquierda). Sin embargo, muchas otras patologías se ven aumentadas en sujetos con sobrepeso y obesidad, como son diversos tipos de neoplasias (colon, mama, endometrio, próstata, vesícula), enfermedades del aparato digestivo (reflujo gastroesofágico, esteatosis hepática no alcohólica), enfermedades neurodegenerativas (Alzheimer, Parkinson), alteraciones del aparato locomotor (artrosis de rodilla y cadera), entre otras (5). A pesar que la asociación entre obesidad y otras patologías ha sido descrita ampliamente, el mecanismo fisiopatológico que asocia exceso nutricional con las enfermedades mencionadas no es bien conocido. El avance del conocimiento de los últimos años permite plantear que las características de la expansión del tejido adiposo tendrían un papel fundamental en determinar el potencial patológico de la obesidad (6).
La obesidad ha sido definida por la OMS como la acumulación anormal o excesiva de grasa que puede ser perjudicial para la salud. La expansión del volumen del tejido adiposo es secundaria a un balance energético positivo, mantenido en el tiempo. Los triglicéridos sintetizados como consecuencia del exceso de energía, se depositan en el citoplasma de una célula especializada para almacenar lípidos: el adipocito. El balance energético es la resultante de la diferencia entre la energía que ingresa al sistema y la energía utilizada por el organismo. El ingreso de energía está dado por la ingesta de alimentos. El gasto energético está determinado principalmente por la mantención de las funciones básicas del organismo, comprendidas en lo que se denomina el metabolismo basal. También se consume energía en la termogénesis inducida por los alimentos y la actividad física. Además, en situaciones especiales debe considerarse el gasto determinado por crecimiento, embarazo, lactancia o enfermedad. La distribución de este gasto dependerá, fundamentalmente, de la importancia relativa del componente de actividad física (7).
Cabe destacar que el balance energético no es el simple resultado de la ingesta y la actividad física, ya que diversos factores genéticos y ambientales influyen en determinar cuanto de lo ingerido ingresa realmente al organismo. Por ejemplo, la matriz alimentaria en que vienen los nutrientes, el horario de consumo, la flora bacteriana, entre otros componentes, influirán en la absorción de nutrientes. Así mismo, el gasto energético no solo depende de la actividad física realizada, puesto que el gasto que ésta genere está en directa asociación con el metabolismo basal, el que depende de la masa muscular. Además, existirían individuos más eficientes del punto de vista metabólico, que presentan menores pérdidas energéticas, por ejemplo por el concepto de generación de calor.
Tejido adiposoDurante muchas décadas se consideró al tejido adiposo como un depósito inerte de triglicéridos, reconociendo como única función del adipocito la de participar en la regulación de la utilización de estas reservas energéticas por el organismo, vía lipogénesis y lipólisis (formación y rompimiento de moléculas de triglicéridos, respectivamente). Sin embargo, a mediados de la década de los 90 se identificó la leptina, un factor proteico producido en el tejido adiposo, pero con acción en el sistema nervioso central (8, 9). Con esto se dio paso a la caracterización de una serie de factores secretados por este tejido: las adipoquinas. De esta manera, en la actualidad se considera al tejido adiposo como un tejido endocrino (6, 10, 11).
El estudio del tejido adiposo ha presentado grandes avances en la última década, de tal modo que actualmente se reconocen diferentes características biológicas que pueden interactuar condicionando ya sea un tejido inerte del punto de vista metabólico y cardiovascular, o un tejido que induce alteraciones del metabolismo de lípidos y de glucosa, además de alteraciones vasculares, entre otras patologías. En la actualidad,se piensa que gran parte de la asociación que existe entre obesidad y enfermedad metabólica y cardiovascular depende del buen o mal funcionamiento del tejido adiposo (12, 13).

Son múltiples las características biológicas que pueden alterarse en el tejido adiposo en condiciones de obesidad. Estas características están íntimamente asociadas entre sí, pero serán presentadas de forma aislada, a fin de permitir una mejor comprensión de la fisiología (o fisiopatología) del tejido adiposo en condiciones de obesidad. Dentro de las características que pueden determinar la nocividad del tejido adiposo, se encuentran aquellas relacionadas con su topografía, poder de lipólisis/ lipogénesis, perfil secretor, tipo de expansión (hiperplasia o hipertrofia), estado inflamatorio, niveles de oxígeno, entre otras.
Lipólisis/ lipogénesisEl adipocito es una célula que presenta la maquinaria adecuada para almacenar una gota lipídica en su citoplasma, sin que se produzca daño asociado. Este depósito no es pasivo, existe un sistema de regulación de la lipogénesis (entrada a la célula de ácidos grasos que se esterifican con una molécula de glicerol, originando los triglicéridos). También la lipólisis, salida de ácidos grasos desde la célula adiposa, es un fenómeno activo y regulado por diversas señales. Aunque no es el objetivo de esta revisión el entregar detalles de estas vías regulatorias, solo se señalará que la gota lipídica está cubierta de diferentes proteínas que le dan estabilidad y permiten la salida o entrada de ácidos grasos frente a determinadas señales. Por ejemplo, dentro de las señales que regulan la lipogénesis tenemos la de la insulina (que la estimula) y la de la leptina (que la inhibe); entre aquellas que modulan la lipólisis destacan las catecolaminas (prolipolíticas) y la insulina (antilipolítica). El exceso de ácidos grasos circulantes puede traer como consecuencia su depósito en células no adiposas, generando insulinoresistencia a través de los fenómenos de lipotoxicidad (14).
TopografíaEl principal depósito adiposo es el subcutáneao, que se encuentra en una importante superficie del cuerpo, pero principalmente en la zona proximal de las extremidades inferiores y en el abdomen. Adicionalmente, existen diferentes depósitos adiposos dentro de la cavidad torácica, abdominal y pélvica. Los individuos eutróficos tienen un desarrollo muy limitado de estos tejidos adiposos que rodean las vísceras; en condiciones de exceso nutricional, el depósito de triglicéridos ocurre tanto a nivel subcutáneo como visceral. Los principales depósitos adiposos viscerales son los que se desarrollan en el mesenterio y el epiplón mayor, pero también hay desarrollo de tejido adiposo a nivel perirrenal, pericardíaco y, según la magnitud de la obesidad, alrededor de otros órganos como páncreas, vesícula, vasos sanguíneos, próstata, entre otros. A pesar de que ambos tejidos adiposos están compuestos por los mismos tipos celulares, la evidencia sugiere que tienen una biología muy diferente entre sí, tanto en términos de lipólisis y lipogénesis como de las otras características biológicas mencionadas (15). No se ha identificado la razón por la cual algunos sujetos tendrían tendencia a la acumulación adiposa en uno u otro tipo de tejido adiposo. Se piensa que cuando el tejido adiposo subcutáneo no es capaz de hacerse cargo de depositar el exceso de triglicéridos, éstos se depositan de manera ectópica, aumentando el volumen de los tejidos adiposos viscerales (16). La lipodistrofia es un modelo que permite entender de mejor manera este fenómeno. Los sujetos lipodistróficos, tienen un desarrollo insuficiente del tejido adiposo subcutáneo, por lo que los triglicéridos se depositan en tejidos adiposos viscerales o de forma ectópica; estos sujetos son delgados, pero metabólicamente enfermos (17).
Ya a mediados del siglo pasado se describió que el tejido adiposo a nivel central (obesidad tipo manzana) sería más perjudicial que el que se deposita a nivel periférico (obesidad tipo pera). La nocividad del tejido adiposo intraabdominal ha sido atribuida a la posibilidad de que este depósito libere ácidos grasos directamente al hígado vía vena porta. Sin embargo, se ha evidenciado que la mayoría de los ácidos grasos que llegan al hígado provienen del tejido adiposo subcutáneo (por su mayor volumen) (18). Probablemente la vía portal pueda dar cuenta de un drenaje directo al hígado de otros productos de secreción del tejido adiposo, las adipoquinas. Esta posibilidad está avalada por los niveles de estas hormonas en la porta, los que son mayores a los de la circulación sistémica (19). Además, el tejido adiposo que se encuentra en la periferia de los órganos puede tener un efecto en la fisiología de éstos a través de un efecto paracrino de las adipoquinas, como se ha evidenciado para el tejido perivascular (20, 21).
CelularidadAntiguamente se pensaba que el tejido adiposo tenía una dotación determinada de células adiposas, definidas precozmente en el desarrollo, durante las etapas intrauterinas o los primeros años de vida. Sin embargo, en la última década se ha conocido la capacidad de algunas células (tipo fibroblastos) del tejido adiposo para diferenciarse a células adiposas maduras en cultivos celulares, frente a estímulos específicos. Esta capacidad adipogénica pareciera también estar presente in vivo, según lo descrito por Spalding y colaboradores (22). Estos investigadores, basándose en los niveles variables de carbono 14 en la atmósfera, han entregado evidencia sobre la generación de nuevas células adiposas en la vida adulta. No existe evidencia que permita estimar si la capacidad adipogénica es homogénea a lo largo de la vida o hay momentos de mayor plasticidad tisular. Tampoco hay certeza si esta capacidad varía en condiciones de obesidad. Como hemos visto, en condiciones de exceso nutricional, la alternativa más inocua es que los ácidos grasos se depositen a nivel del tejido adiposo subcutáneo. Por lo tanto, un tejido adiposo que tenga una buena capacidad adipogénica permitirá la expansión de la masa grasa tanto por hipertrofia (crecimiento del tamaño de las células) como por hiperplasia (incorporación de nuevas células). Éste será un tejido eficiente en el almacenaje del exceso energético. Sin embargo, aquellos tejidos que tienen una capacidad limitada de expandirse por hiperplasia, solo podrán aumentar acosta de hipertrofia celular. Aún cuando el adipocito puede aumentar su diámetro en 20 veces, esta capacidad no sería ilimitada por lo que, en caso de mantenerse el balance energético positivo en el tiempo, los ácidos grasos debería ir a depositarse de forma ectópica, ya sea en tejido adiposo visceral, o en el citoplasma de otras células que se ven afectadas en su fisiología. Esta es la llamada teoría del rebalse (16). Además, se piensa que los adipocitos hipertróficos tendrían un peor perfil secretor (23).
InflamaciónActualmente la obesidad se considera un estado inflamatorio crónico, de intensidad leve a moderada (24, 25). La evidencia de este estado inflamatorio exacerbado en condiciones de exceso se da a diferentes niveles. A nivel sistémico, se manifiesta por un aumento de los mediadores inflamatorios plasmáticos como la proteína C reactiva (PCR), el factor de necrosis tumoral alfa (TNFα) o de los leucocitos circulantes (cambios cuanti y cualitativos) (26, 27). Además, hay cambios a nivel intracelular en diferentes tipos de células asociadas al metabolismo, como son las células adiposas, hepáticas, músculoesqueléticas y beta pancreáticas. Estos cambios se manifiestan por una activación del principal modulador de la respuesta inflamatoria celular: el factor de transcripción NF-κB. Cuando este factor ingresa al núcleo, permite la expresión de diferentes moléculas proinflamatorias (28). La inflamación asociada a obesidad también se manifiesta a nivel de algunos órganos, los que se ven infiltrados de células inflamatorias. Es el caso del páncreas (29), endotelio (30) y el propio tejido adiposo (31). A pesar de que en condiciones normales el tejido adiposo contiene algunas células inflamatorias, el tejido de los individuos obesos muestra una proporción muy aumentada de éstas (31). En modelos murinos con deleciones génicas específicas, se ha visto que aquellos ratones obesos que no aumentan la infiltración de macrófagos no presentan las alteraciones metabólicas propias de la obesidad (31). De manera análoga, los ratones delgados que ven aumentada la cantidad de macrófagos en el tejido adiposo, presentan las alteraciones metabólicas propias de los obesos (32). De esta manera, la presencia de los macrófagos sería de vital importancia en la asociación de obesidad y enfermedad metabólica y cardiovascular.
Existen múltiples hipótesis sobre cuál sería la razón porque estas células inflamatorias infiltran el tejido adiposo, entre las que destaca la hipoxia tisular (33), hipertrofia adipocitaria (23), aumento en la expresión de quemoquinas (10, 11), alteraciones de la matriz extracelular (10), liberación de ácidos grasos (34), muerte adipocitaria (35), entre otras. También existe evidencia que los leucocitos circulantes mononucleares (entre los que están los monocitos, precursores circulantes de los macrófagos tisulares), se encuentran en un estado proinflamatorio en condiciones de obesidad (27). Esto condiciona una mayor susceptibilidad a reaccionar a las señales quimioatrayentes de los distintos tejidos, lo que explicaría la infiltración de otros órganos. Recientemente se ha descrito la presencia de linfocitos en el tejido adiposo, aún cuando no se comprende bien las consecuencias que su presencia tiene ahí (36). Estos hallazgos sugieren que es posible que la obesidad no solo esté activando la inmunidad innata, sino que también la inmunidad adaptativa. Hasta la fecha, no se ha identificado algún antígeno en particular que pudiera estar explicando este fenómeno.
Tensión de oxígenoEl tejido adiposo de los sujetos obesos presenta menores presiones de oxígeno que el de los sujetos normopeso (33). Esto sería secundario a un crecimiento insuficiente de la red vascular, a una hipertrofia celular excesiva (que dificultaría la difusión de oxígeno) o a la vasocontricción secundaria a las adipoquinas circulantes en el ambiente local (por ejemplo secundaria al aumento de angiotensina II). El tejido adiposo en condiciones de hipoxia aumenta la síntesis de mediadores inflamatorios, muy probablemente por la activación del factor de transcripción inducible por hipoxia (HIF1) que está en la zona reguladora de la mayoría de los genes de factores proinflamatorios (33, 37).
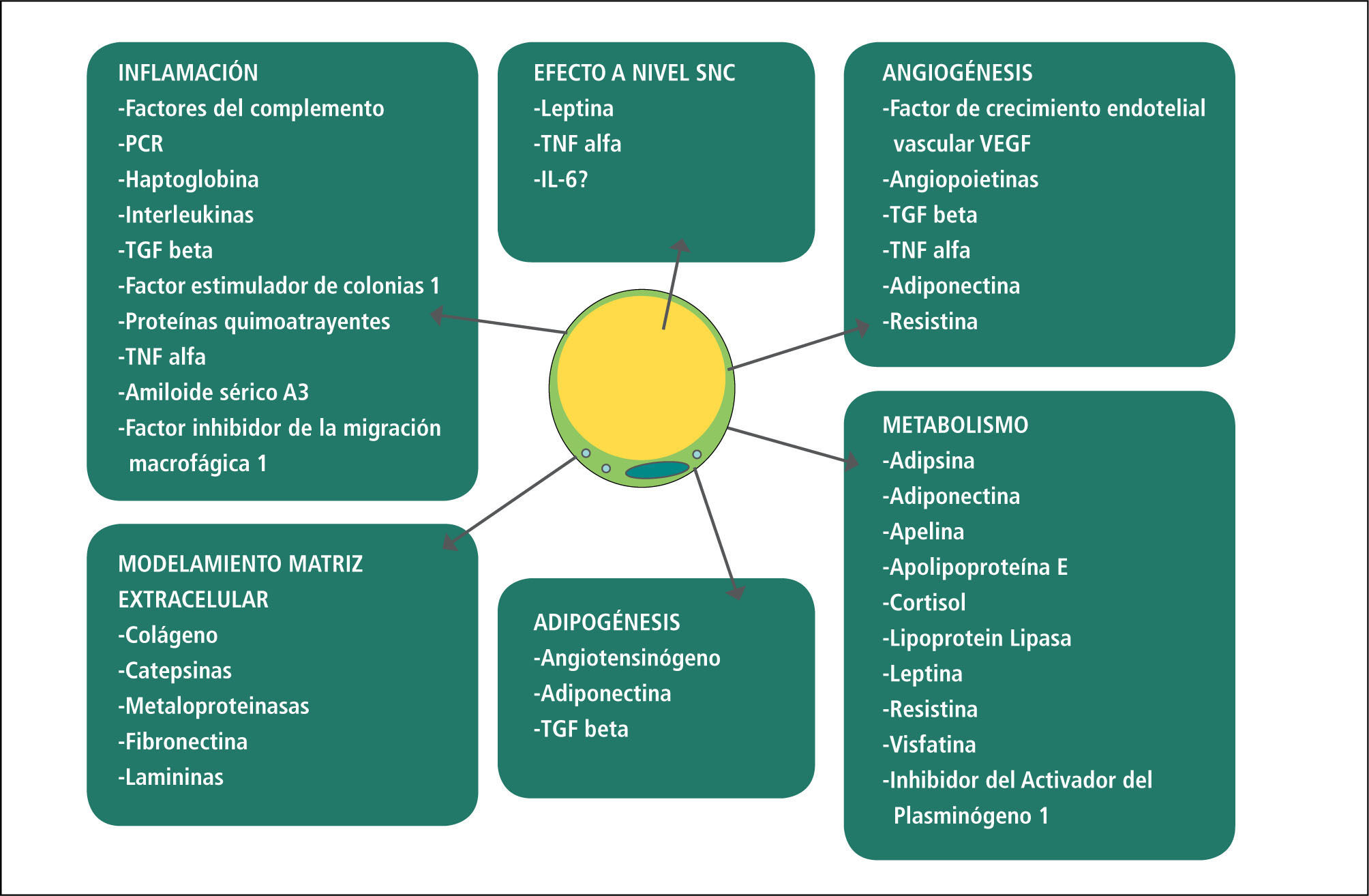
Perfil secretorTras la descripción inicial de la leptina, se han identificado múltiples productos de secreción del tejido adiposo. La lista de estos factores está en constante crecimiento, por lo que solo un porcentaje de ellos presenta una adecuada caracterización funcional. La mayoría de las adipoquinas pueden ser sintetizadas por cualquiera de los componentes celulares del tejido adiposo: adipocitos, preadipocitos, células del sistema inmune, células endoteliales y fibroblastos, aunque algunas son productos específicos de un solo tipo celular. Estos productos de secreción pueden tener efectos locales (actividad autocrina), influir en la fisiología de órganos adyacentes como son corazón, vasos sanguíneos u otros (efecto paracrino) o en órganos muy distantes, vía sanguínea (efecto endocrino). El perfil secretor en condiciones fisiológicas normales tendría un efecto de homeostasis metabólica. A través de estas señales, se coordina información sobre reservas energéticas, apetito, gasto energético, sensibilidad a hormonas claves del metabolismo como la insulina, entre otras. De esta manera, se integran funciones del tejido adiposo con otros órganos como páncreas, tubo digestivo, hígado y cerebro. Sin embargo, en condiciones de obesidad este perfil secretor se modifica, estableciendo una comunicación alterada entre los diferentes órganos (10-13, 20). Por ejemplo, en condiciones de expansión del tejido adiposo, las señales de exceso de depósitos no son censadas adecuadamente a nivel del sistema nervioso central, que no responde con la esperada disminución del apetito (8). Por otra parte, los productos de secreción del tejido adiposo condicionan una menor sensibilidad a la insulina, tanto a nivel local como sistémico (13, 20). Adicionalmente, las adipoquinas secretadas en condiciones de obesidad son proinflamatorias y protrombóticas, favoreciendo las condiciones fundamentales en la etiopatogenia de la ateromatosis. La formación de placas de ateromas (antes entendida como un simple depósito de colesterol en los vasos sanguíneos) es producto de la interacción entre un sistema inmune exacerbado, cuyas células migran de forma activa al espacio subendotelial, y las alteraciones metabólicas (aumento de LDL oxidadas). El proceso inflamatorio que ahí ocurre, junto con el estado procoagulante, desencadena el evento coronario agudo que constituye la principal morbimortalidad asociada a obesidad (30).

A continuación se revisarán las características de algunas de las adipoquinas mejor caracterizadas. El objetivo no es hacer una revisión de la literatura, puesto que existe bibliografía extensa para cada una de los productos de secreción del tejido adiposo. La idea es exponer, de manera simple, información que permita entender de mejor manera las interacciones que se establecen entre tejido adiposo y otros órganos, a través de su capacidad de síntesis y secreción de adipoquinas, con sus influencias locales y sistémicas. Es importante mencionar que existen múltiples interacciones entre las diferentes adipoquinas, por lo que generalmente se establece un círculo vicioso, en que el aumento en la secreción de una de ellas tiene en los niveles de expresión del resto, magnificando el efecto inicial (13).
LeptinaEl primer efecto descrito para la leptina fue en relación a la modulación del apetito y el gasto energético. A medida que disminuye la masa adiposa, también lo hacen los niveles circulantes de esta hormona. De esta manera, existen mecanismos para compensar el déficit nutricional, aumentando la ingesta y disminuyendo el gasto energético (8). Sin embargo, los mecanismos no son tan eficientes en el caso del exceso. Del punto de vista evolutivo, los mecanismos para disminuir el apetito o aumentar el gasto no están bien regulados (38). El fenómeno es aún más grave debido que los obesos desarrollan una resistencia a la leptina. Es por esto que la administración farmacológica de esta hormona ha mostrado un efecto benéfico solo en aquellos individuos que no la sintetizan por algún trastorno genético. Sin embargo, en los últimos años también se le han descrito otros efectos atribuibles a esta hormona, como la estimulación de la lipólisis en tejido adiposo (39), inhibición de la secreción de insulina por la célula beta pancreática (40), inhibición de la génesis de esteroides inducida por insulina en el ovario (41), entre otros. También se le ha identificado una acción inmunomoduladora, que explicaría en parte la respuesta inmunológica deficiente en sujetos desnutridos (42). Además de todos los efectos deletéreos señalados, esta hormona tendría un efecto nocivo directo para la salud cardiovascular, por su acción pro-trombótica y pro-oxidante (43). Por último, se ha sugerido que la leptina jugaría un papel importante en la asociación que existe entre obesidad y algunas neoplasias (44).
ResistinaLa resistina es otra de las adipoquinas que aumenta con la expansión de la masa adiposa. Si bien en un principio se le adjudicó un efecto en el deterioro de la sensibilidad insulínica, esto no está tan claro en la actualidad (45). Los adipocitos no son la única fuente de esta proteína, la que también es producida y secretada por monocitos, células ß pancreáticas, miocitos entre otras. Esta es una proteína proinflamatoria que estimula la síntesis de otros mediadores clásicos de la inflamación como interleuquinas y TNFα. También tiene un efecto nocivo a nivel endotelial, aumentando la expresión de factores de adhesión (11). En condiciones patológicas, se ha detectado su secreción aumentada en placas de ateroma, alteraciones articulares y enfermedad renal crónica (45).
AdiponectinaLa adiponectina es una de las pocas adipoquinas conocidas que presenta un efecto positivo en la salud metabólica y cardiovascular (46). La adiponectina circulante está disminuida en condiciones de obesidad. Este fenómeno ha sido implicado en la génesis de resistencia insulínica, disfunción endotelial, hígado graso no alcohólico, dislipidemia, enfermedad cardiovascular y diversas neoplasias. Gran parte de estas acciones podrían estar dadas por su importante efecto antiinflamatorio (11, 47), el que además se ve amplificado por el efecto de esta hormona en la síntesis de diferentes interleuquinas (48, 49). Además de los efectos periféricos, se ha descrito un efecto en los centros hipotalámicos de apetito, siendo reconocida como un factor anorexigénico (50).
AngiotensinaEl tejido adiposo puede sintetizar y secretar angiotensinógeno y los diversos factores que permiten su conversión a angiotensina II. Su producción está aumentada en condiciones de obesidad. Se estima que sus efectos serían principalmente a nivel local, sin paso importante a la circulación sistémica, por lo que no habría efecto deletéreo en los valores de presión arterial. A nivel del tejido adiposo, el principal efecto de estas hormonas tampoco estaría relacionado al tono vascular. Su acción fundamental sería la de disminuir la adipogénesis y estimular la hipertrofia adipocitaria (51). De esta manera, una mayor expresión de este eje tendría como consecuencia una expansión del tejido adiposo principalmente basada en la hiperplasia de los adipocitos, los que se vuelven hipertróficos y por ende disfuncionales. Además tendría un efecto proinflamatorio y de alteración del metabolismo lipídico y glucídico (51, 52).
Otras citoquinasTanto las células adiposas como preadiposas sintetizan diversas citoquinas, fundamentalmente del tipo inflamatorio. Las células preadiposas tendrían un perfil secretor más inflamatorio que las células maduras (53). Sin embargo, se estima que las células del sistema inmune que infiltran el tejido adiposo serían la principal fuente de estas citoquinas (54). Algunos mediadores inflamatorios secretados por el tejido adiposo son TNFα y las interleuquinas 1 y 6 (IL1 y IL6). Los principales efectos a nivel local son la disminución de la sensibilidad a la insulina y la inhibición de la adipogénesis (55, 56). Además, estas señales alcanzan al hígado, gatillando el aumento de las proteínas de fase aguda, lo que amplifica esta señal inflamatoria local y por ende su efecto deletéreo en la sensibilidad insulínica. Adicionalmente, se ha identificado la síntesis de interleuquinas antiinflamatorias, como es el caso de la IL10 (57).
QuimioatrayentesLas quimoquinas son un tipo especial de adipoquinas, cuya función es la de favorecer la interacción de los leucocitos circulantes con el endotelio de los vasos sanguíneos del tejido, induciendo su migración al lugar de la inflamación. El principal quimoatrayente secretado por el tejido adiposo en condiciones de obesidad es la proteína quimoatrayente de monocitos 1 (MCP1, por su sigla en inglés). Sin embargo, también hay un aumento de otras como, como CCL5 (también conocida como RANTES, por su sigla en inglés), la que atrae principalmente linfocitos.
Funcionalidad del tejido adiposoLas características biológicas descritas han sido presentadas de forma aislada solo con fines didácticos, puesto que la realidad in vivo es mucho más compleja: estas características están íntimamente asociadas entre sí y el mal funcionamiento de una de ellas influye en la alteración de las otras, generando un círculo vicioso. De esta manera, cuando la capacidad de adipogénesis está limitada, el crecimiento de la masa grasa es preferentemente por hipertrofia (16). Los adipocitos hipertróficos presentan un perfil secretor más inflamatorio, probablemente gatillado por la hipoxia relativa (23, 37). De esta manera, la llegada de monocitos (y su maduración a macrófagos) está aumentada. Las citoquinas producidas por los macrófagos tornan más inflamatoria las adipoquinas secretadas por el tejido adiposo en su conjunto (54). Además, la presencia de los macrófagos inflamatorios influye negativamente en los adipocitos, alterando su perfil secretor a uno más inflamatorio y aterogénico y aumentando la lipólisis (12). Los ácidos grasos, a su vez, inducen mayor inflamación en los macrófagos (12). Asimismo, los productos secretados por los macrófagos inhiben la adipogénesis (55), empeorando la situación inicial. El cambio relativo de los diferentes tipos celulares también puede alterar el perfil lipídico, se piensa que las células preadiposas tienen un perfil secretor más aterogénico que las células adiposas maduras (53), por lo que la limitada adipogénesis también tendría un efecto nocivo por la vía de acumular más células preadiposas. El tejido adiposo visceral tiene mayor infiltración de células inflamatorias, menor capacidad adipogénica y peor perfil secretor que el tejido adiposo subcutáneo (58). Así como la alteración de una de las características se potencia generando círculos viciosos de mal funcionamiento del tejido, esta interacción también se da en el sentido inverso, siendo el buen funcionamiento la posible partida de un círculo virtuoso. El reporte de Klöting y cols (59) apoya lo planteado. En este estudio se comparan sujetos con igual magnitud de obesidad, pero con diferentes sensibilidades a la insulina. Aquellos obesos con sensibilidad conservada a la hormona mostraron un me nor contenido de grasa visceral, un perfil plasmático de adipoquinas menos nocivo (alto en adiponectina), menor proporción de adipocitos hipertróficos, menor infiltración del tejido adiposo por macrófagos y mayor sensibilidad insulínica por parte de los adipocitos. Este reporte constituye una excelente evidencia a favor de la hipótesis de la funcionalidad del tejido adiposo como determinante del potencial patogénico de la obesidad.
Manejo de la disfunción del tejido adiposoEs evidente que el foco terapéutico de la obesidad debe ser el favorecer las condiciones que permitan un balance energético negativo, mantenido en el tiempo. De esta manera, las alteraciones determinadas por la expansión del tejido adiposo debieran ser revertidas. Se desconoce si las características con la que se expanda o revierta la expansión del tejido adiposo difieren según edad. De ser esto así, se podría plantear que la disminución de los depósitos no siempre logrará una completa resolución de la disfunción del tejido.
Se ha señalado que la realización de ejercicio tiene un efecto benéfico, adicional al asociado solamente a la disminución de la masa grasa a través de la contribución a un balance energético negativo. Se ha identificado una secreción transitoria de IL6 por parte del músculo esquelético, en la hora siguiente a la realización de ejercicio. Aún cuando esta interleuquina ha sido caracterizada como proinflamatoria, su aumento transitorio desencadenaría un aumento en la secreción de mediadores antiinflamatorios sistémicos, como cortisol, IL10 y el antagonista del receptor de IL1, el IL1RA (60). No se comprende cuales factores determinan este efecto paradojal de IL6. Por otra parte, el ejercicio disminuiría el estado inflamatorio de diferentes células circulantes del sistema inmune, lo que contrarrestaría la migración aumentada de estas células al tejido adiposo (60).
La modulación farmacológica de la disfunción adipocitaria también podría constituir una opción terapéutica. Sin embargo, no ha habido avances importantes en esta área. La inhibición del proceso inflamatorio podría traer consecuencias adversas en términos de la inmunidad sistémica, por lo que no es planteable con las herramientas farmacológicas actuales. Sin embargo, muchos de los fármacos que se utilizan de forma habitual para el manejo de las condiciones metabólicas y cardiovasculares asociadas a la obesidad presentan un efecto antiinflamatorio.
Este es el caso de los inhibidores del receptor de angiotensina II, las estatinas o los bloqueadores del receptor de la glicoproteína plaquetaria IIb/IIIa (61-63). Un caso especial está constituido por la tiazolinedionas, hipoglicemiantes agonistas del factor de transcripción PPARγ, implicado íntimamente en la adipogénesis y en las vías de señalización antiinflamatorias. Su uso aumenta la expansión del tejido adiposo subcutáneo y disminuye el estado inflamatorio, mejorando el metabolismo glicémico (64). Sin embargo, la mayoría de estos fármacos ha tenido que ser retirado del mercado, por el mayor riesgo cardiovascular asociado a su uso en estudios clínicos. Este fenómeno no ha sido bien comprendido, pero ha limitado de forma importante el uso de esta terapia que parecía tan promisoria (65-67).
Los recientes hallazgos sobre la presencia de tejido adiposo pardo en el tejido adiposo blanco de adultos, dan paso a nuevas posibilidades de modulación de la fisiología del tejido adiposo (68, 69). La principal característica del tejido adiposo pardo es la expresión de la proteína desacoplante 1 (UCP1, por su sigla en inglés). Esta proteína está encargada de reducir la gradiente electroquímica de protones en el espacio intermembrana de la mitocondria, disminuyendo la síntesis de adenosín trifosfato (ATP, por su sigla en inglés) y disipando la energía en forma de calor (70). La conversión de adipocitos blancos a adipocitos pardos no sería posible, ya que tendrían diferentes precursores celulares (70). Sin embargo, se ha demostrado que la presencia de grasa parda podría ser inducida, por ejemplo, por temperaturas frías (69). Hay muchas interrogantes en relación a la existencia de este tejido pardo; teóricamente, su presencia y funcionamiento podría ser estimulado, lo que determinaría un aumento del gasto calórico, disminuyendo el balance energético positivo.
ConclusiónEn síntesis, el exceso de grasas derivadas de un balance energético positivo mantenido en el tiempo, determina la expansión del volumen del tejido adiposo. Las características biológicas de este proceso determinarán las consecuencias metabólicas y cardiovasculares de la obesidad. La modulación de estas características se vislumbra como un potencial terapéutico para la disminución de la morbimortalidad asociada a la malnutrición por exceso. Sin embargo, se debe tener siempre en cuenta que el verdadero manejo de la obesidad es el revertir el balance energético positivo, a través de la modulación de los factores individuales y colectivos que favorezcan un metabolismo energético alterado.
La autora declara no tener conflictos de interés, en relación a este artículo.