




La investigación de los factores etiopatogénicos de la obesidad, sean estos ambientales o genéticos, ha influido fuertemente en el desarrollo de las estrategias para controlar la enfermedad. Se considera a la obesidad como una enfermedad en la cual confluyen factores genéticos y ambientales, pero no se han dilucidado cabalmente los mecanismos de regulación del balance energético, cuyas alteraciones favorecen el incremento de la adiposidad en humanos. Entre los factores ambientales, además de la alimentación alta en calorías y el sedentarismo, se describen factores predisponentes sociales, culturales y familiares, así como situaciones o etapas fisiológicas de la vida que pueden gatillar aumento de peso. Aunque los factores y patologías genéticas se pueden evidenciar en un bajo porcentaje de los sujetos obesos, la predisposición genética a la obesidad se puede expresar a través de algunas características metabólicas, como un bajo gasto energético de reposo, mala oxidación de grasas o un bajo nivel de actividad física espontánea.
The investigation of the etiological factors of obesity, whether genetic or environmental, has strongly influenced the development of strategies to control the disease. Obesity must be considered as a disease in which genetic and environmental factors converge. However, the mechanisms of regulation of the energy balance, which changes favoring increased adiposityin humans, are not fully elucidated. Among environmental factors, in addition to high-calorie diet and sedentary lifestyle, it has been described social, cultural and family predisposing factors, as well as physiological conditions or stages of life that can trigger weight gain. Although genetic factors and pathologies can be evidenced in a low percentage of obese subjects, genetic predisposition to obesity can be expressed through some metabolic characteristics such as low metabolic rate, poor oxidation of fats or a low level of spontaneous physical activity.
El conocimiento de la etiopatogenia de la obesidad debiera constituir una base sólida para el desarrollo de las estrategias para controlar la enfermedad. Entre las intervenciones destacan las medidas preventivas sobre factores ambientales, tales como la promoción de hábitos alimentarios saludables y el aumento en la actividad física, así como el descubrimiento de nuevas herramientas terapéuticas para enfrentar la obesidad asociada a una fuerte predisposición metabólica o genética.
La obesidad es una enfermedad en la cual confluyen distintos factores genéticos y ambientales, pero no se han dilucidado cabalmente los mecanismos de regulación del balance energético, cuyas alteraciones favorecen el incremento de la adiposidad en humanos. Entre los factores ambientales relacionados con la epidemia mundial de obesidad destacan la mayor disponibilidad de alimentos y el aumento progresivo en la conducta sedentaria (1), sobre la base de una carga genética que en el ser humano no se ha modificado por siglos. La teoría de los genes ahorradores (“Thrifty genes”) establece que durante la evolución del ser humano se fueron seleccionando, en forma natural, ciertas variantes genéticas que permitían lograr un mayor almacenamiento de energía en el tejido adiposo y un bajo gasto energético ante condiciones de vida caracterizadas por la escasez o dificultad en la obtención de los alimentos y/o exposición a largos periodos de hambruna (2). Esta carga genética enfrentada a un estilo de vida sedentario y alta ingesta de calorías constituye el principal factor de riesgo de obesidad, especialmente en las culturas occidentales.
Un interesante ejemplo acerca de la importancia relativa de factores géneticos y ambientales lo aporta el estudio realizado por Christakis y Fowler (3), sobre el rol de las redes sociales en la epidemia de obesidad. Tomando la información de 12.000 sujetos interrelacionados, seguidos durante 30 años en el Estudio del Corazón de Framingham, los autores encontraron que el riesgo de hacerse obeso durante el periodo analizado se elevaba hasta en 171% si uno de sus amigos se hacía obeso. También se encontró un efecto, aunque menor, cuando la relación era de esposo (37% mayor riesgo) o de parentesco directo (40% mayor riesgo). Aunque el estudio no permite establecer factores etiológicos precisos, permite estimar la gran relevancia que tiene el ambiente en el desarrollo de la obesidad.
Regulación de los depósitos de energía1Regulación del gasto energéticoAnte condiciones de sobrealimentación o hipoalimentación el organismo cuenta con mecanismos adaptativos que, aunque en forma insuficiente, modifican los distintos componentes del gasto energético y/o la ingesta alimentaria. Después de un periodo de balance energético positivo que conduce a un incremento de peso, aumenta el gasto energético tanto de reposo como post-ejercicio y post-alimentación. Esto se relaciona a un aumento en los niveles de T3 y del tono simpático, con incremento en los niveles de insulina y leptina. Por el contrario, luego de un periodo de balance energético negativo con disminución de peso, se reduce el gasto energético, el tono simpático y los niveles de T3, leptina e insulina, aumentando los niveles de ghrelina (4-6).
2Patrón de alimentación en la patogénesis de la obesidadLos factores ambientales son los principales condicionantes de la ingesta alimentaria, y entre ellos destacan los hábitos y tradiciones familiares, la presión social, la disponibilidad de alimentos, y los factores psicológicos. Existe clara evidencia de que los obesos consumen en forma habitual una dieta con mayor proporción de grasas que los sujetos con peso normal (7-10). Los individuos obesos y post-obesos manifiestan una preferencia por sabores provenientes de grasas. Esto podría precipitar la expresión de una predisposición genética como una baja tasa de oxidación de grasas (11). La dieta de grupos de obesos estudiados en Estados Unidos ha mostrado tener un 5 a 8% más calorías como grasas que la de grupos controles. Una mayor ingesta de grasas en la dieta puede inducir mayor ganancia de peso, porque estos macronutrientes tienen mayor densidad calórica (9kcal./g), se incorporan en alimentos más apetecibles, producen menos saciedad e inducen menor termogénesis post-prandial. Sin embargo, no es indispensable una sobreingesta de grasas para inducir obesidad, ya que esta puede aparecer por exceso de ingesta de otros aportadores de energía, como los carbohidratos y alcohol, cuya oxidación es proporcional a la ingesta y su capacidad de depósito es limitada (12).
3Actividad física y peso corporalLa menor actividad física como causa de obesidad es una hipótesis atractiva, dado que el aumento en la prevalencia de obesidad se acompaña de un aumento en el estilo de vida sedentario, y aquellos sujetos exitosos en la mantención del peso perdido practican hasta 80 minutos diarios de actividad física (13). Por otra parte, el aumento en el número de automóviles y de horas frente al televisor o computador puede tener un impacto en el nivel de actividad física (14). No obstante, el análisis del nivel de actividad física mediante agua doblemente marcada en poblaciones de países desarrollados y en desarrollo no muestra mayores diferencias. Más aún, al comparar el nivel de actividad física en las últimas cuatro décadas, se observa que la reducción del gasto energético por actividad física no explica el aumento de la obesidad (15). Resultados similares fueron obtenidos al calcular el flujo energético (ingesta v/s gasto energético) estimado a partir del gasto energético total (mediante agua doblemente marcada en 963 niños y 1.399 adultos) y los cambios en la ingesta energética en EE.UU. desde 1970 al 2000 (mediante hojas de balance ajustadas por pérdidas). Estos estudios concluyeron que virtualmente toda la ganancia de peso puede ser explicada por el aumento en la ingesta energética (16). Estos estudios resaltan el papel controversial de la actividad física en la actual prevalencia de obesidad y refuerzan la necesidad de estudios prospectivos para una mejor comprensión del fenómeno.
Aún así, un patrón de actividad física sedentario se ha asociado a mayor índice de masa corporal en estudios transversales (17-19), y un bajo nivel de actividad física recreativa en adultos predice mayor aumento de peso en estudios longitudinales (20). Un mayor nivel de actividad física habitual se ha asociado a un mayor gasto energético de reposo (21), aunque probablemente este efecto esté condicionado por una composición corporal con predominio de masa magra (22). Además, se ha demostrado que el entrenamiento físico aumenta la oxidación de lípidos, pudiendo así ser un factor protector ante una elevada ingesta de grasas (23).
4Conducta alimentaria y neurofisiología de la alimentaciónLa conducta alimentaria es un acto consciente sobre el cual influyen factores ambientales y estímulos endógenos, integrados a nivel de sistema nervioso central.
La capacidad de regular el apetito y la saciedad a nivel hipotalámico ha sido demostrada por la identificación de señales que incrementan o disminuyen el apetito, así como los mediadores químicos involucrados. Los núcleos arcuato y paraventricular en el hipotálamo ventromedial, son parte de un sistema que integra la composición corporal con la ingesta y el gasto energético. Los estímulos aferentes nerviosos (vagales y cate-colamínicos), y hormonales (insulina, leptina, ghrelina), son recibidos en el hipotálamo donde modulan la liberación de péptidos que afectan la ingesta de alimentos y las señales hacia el eje hipotálamo-hipófisis y hacia el sistema nervioso autónomo (6, 24).
5Alteraciones de la conducta alimentariaLas lesiones traumáticas, quirúrgicas o neoplásicas que afectan el área ventromedial del hipotálamo favorecen el incremento de peso a través de un aumento del apetito, pero constituyen causas infrecuentes de obesidad (25). Factores psicológicos pueden alterar gravemente la conducta alimentaria condicionando la manifestación de cuadros bien definidos y otros inespecíficos. Entre los primeros se describen el síndrome de los atracones (“binge eating disorder”) y el síndrome del comedor nocturno (“night eating disorder”), y entre aquellos menos definidos están los cuadros de hiperfagia y comer compulsivo, que generalmente se presentan asociados a trastornos por ansiedad (25, 26).
Determinantes metabólicos de ganancia de pesoEl estudio de los determinantes de la ganancia de peso requiere de estudios longitudinales (27). Una de las poblaciones más estudiadas por su alto riesgo de obesidad son los indios Pima, ubicados en el sur de Estados Unidos (28). A partir de este grupo proviene la mayoría del conocimiento disponible referido a los factores de riesgo de obesidad, como los que se describen a continuación.
Menor tasa metabólicaExiste una importante variabilidad en la tasa metabólica entre individuos, que persiste incluso al comparar sujetos con peso corporal, composición corporal, edad y sexo similares (29). En el estudio de Ravussin et al (30), se observó que aquellos individuos en el menor tercil de tasa metabólica de reposo tuvieron 8 veces más riesgo de ganar 10kg. de peso que los sujetos en el tercil superior, después de 4 años de seguimiento. En la misma línea, un meta-análisis concluyó que sujetos con historia de obesidad tienen una tasa metabólica de reposo 3 a 5% menor cuando se comparan con sujetos de peso corporal similar. Esta menor tasa metabólica podría contribuir a la reganancia de peso usualmente presente en estos individuos (31).
Menor actividad física espontáneaOtro componente del gasto energético de 24 horas es el atribuible a la actividad física espontánea, el cual da cuenta de un 8 a 15% del gasto energético diario. Este gasto corresponde a aquellas actividades realizadas durante la jornada laboral o durante el tiempo libre, e incluye la mantención de la postura corporal y actividades diversas como mover el cuerpo (o una parte) sin control aparente (ej. mover los pies mientras se permanece sentado) (32). Los estudios longitudinales muestran que la actividad física espontánea es un rasgo familiar, donde un bajo nivel de actividad física espontánea se asocia con mayor ganancia de peso en hombres (33). Aún más, la resistencia a la ganancia de peso inducida por sobrealimentación ha mostrado ser dependiente del aumento en la actividad física espontánea (34).
Menor oxidación de grasasLa composición nutricional de la dieta influye en la etiopatogenia de la obesidad, posiblemente por su efecto sobre el tipo de sustrato oxidado (35). Estudios longitudinales efectuados en indios Pima y en población caucásica muestran que el cuociente respiratorio (un indicador de la mezcla oxidativa de sustratos) de ayuno y de 24 horas es un predictor de ganancia de peso (11,36). En el estudio de Zurlo et al (11), aquellos sujetos en el percentil 90 de cuociente respiratorio (menor oxidación de grasas) tuvieron 2,5 veces mayor riesgo de ganar más de 5kg de peso corporal que aquellos en el percentil 10 (mayor oxidación de grasas). En este mismo sentido, los obesos que mantienen el peso perdido presentan un cuociente respiratorio menor, mientras que aquellos que recuperan peso muestran cuocientes respiratorios más elevados (37, 38).
Estos resultados han sido tradicionalmente considerados una evidencia del defecto en la oxidación de grasas en individuos con riesgo de obesidad. Sin embargo, también pueden sugerir una preferencia por la oxidación de carbohidratos por sobre los lípidos. Tal situación podría reducir el tamaño del depósito de carbohidratos, el cual ha sido relacionado de manera inversa al control de la ingesta energética en ratones (39). En humanos, estudios similares muestran una situación más controversial (40), existiendo estudios con efectos directos, inversos y neutros sobre el control de la ingesta alimentaria. Por otra parte, estudios prospectivos muestran que un mayor tamaño del depósito de carbohidratos previene la ganancia de peso (41).
Menor termogénesis inducidaUsualmente el aumento del peso corporal no es proporcional al balance energético positivo estimado. Frente a un mismo exceso energético la ganancia de peso es altamente variable entre individuos, lo cual sugiere que el exceso de alimentación induce diferentes niveles de termogénesis (42). A este fenómeno se le denomina termogénesis adaptativa y se refiere a los cambios en el gasto energético en respuesta al exceso de ingesta calórica, no atribuibles a cambios en el tamaño y composición corporal. Teóricamente el aumento del gasto energético prevendría una mayor ganancia de peso. Esta noción fue inicialmente sustentada en un estudio de sobrealimentación realizado en prisioneros, en los cuales se necesitó una ingesta energética 50% mayor a la predicha para mantener la nueva masa corporal alcanzada (43). Hallazgos similares fueron reportados por Bouchard et al. (44), al evaluar las diferencias en la ganancia de peso entre pares de gemelos monocigotos sometidos a 84 días de sobrealimentación con 1.000kcal/día. El cambio de masa corporal fue altamente variable, con un aumento promedio de 8,1kg. (4,3 a 13,3kg.). La variabilidad en la ganancia de peso grupal fue tres veces mayor a la observada dentro de cada par, lo cual sugiere que esta respuesta está bajo control genético. Existen otros aspectos que pueden dar cuenta de la variabilidad en la ganancia de peso, entre ellos la adherencia al régimen dietario, aunque estudios bien controlados han mostrado una variabilidad similar en la ganancia de peso (42). Por otra parte, nuestra dificultad para determinar apropiadamente el requerimiento energético para la mantención del peso corporal, y por lo tanto el exceso energético, juega un papel esencial (45). Lo anterior también es influenciado por diferencias en la digestión y absorción, lo que puede modificar la energía biodisponible.
Otro factor a considerar es la composición del tejido ganado (masa grasa v/s masa magra) dado que el costo energético implicado en el depósito de proteínas es mayor al del tejido adiposo (46). También la termogénesis adaptativa ha mostrado depender del contenido proteico de la dieta. Stock observó que dietas con bajo o alto contenido de proteínas se asocian a menor ganancia de peso respecto a dietas con un nivel proteico intermedio (42, 47). Las diferencias en la eficiencia mitocondrial también debieran ser un mecanismo que dé cuenta de la variabilidad en la ganancia de peso. Así, la mantención de una menor o mayor masa corporal, inducida por sub- o sobre-alimentación, resulta en mayor o menor eficiencia del trabajo muscular medido en una bicicleta ergométrica, respectivamente (48). Finalmente, estudios longitudinales de los cambios en el gasto energético y masa corporal en indios Pima muestran que la termogénesis adaptativa ocurre en respuesta a la ganancia de peso, no obstante, su magnitud es pequeña y muy variable entre los individuos (49).
Menor actividad del sistema nervioso simpáticoLa actividad del sistema nervioso simpático (SNS) se asocia directamente con la tasa metabólica basal, efecto térmico de los alimentos y actividad física espontánea (50). Además, se correlaciona de manera negativa con el cuociente respiratorio de 24 horas (51). Por lo tanto, la actividad del SNS pudiera ser uno de los mecanismos subyacentes que dan cuenta de la variabilidad en la ganancia de peso. Una menor actividad del SNS se asocia con menor pérdida de peso en obesos sometidos a dieta hipocalórica (52). Además, los indios Pima poseen menor actividad simpática muscular respecto a sujetos caucásicos de peso corporal similar (53). Esta evidencia se complementa con la observación que la excreción urinaria basal de norepinefrina (un índice global de la actividad del SNS) se asoció de manera inversa con la ganancia de peso en indios Pima, aunque solo en aquellos de sexo masculino (54).
Es importante destacar que varios de estos factores presentan agregación familiar y, por lo tanto, un posible carácter hereditario.
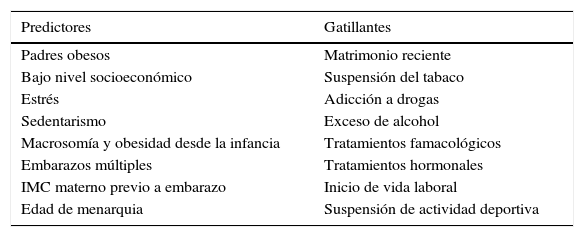
Predictores y gatillantes de ganancia de pesoVarios estudios poblacionales han identificado factores predisponentes de obesidad tales como: padres obesos, bajo nivel socioeconómico, estrés, sedentarismo, obesidad infantil, macrosomía fetal y embarazos múltiples (20,55). Entre los factores gatillantes de ganancia de peso se señalan la adicción a drogas, algunos tratamientos famacológicos y hormonales (25), el inicio de vida laboral, la suspensión de actividad deportiva y el matrimonio reciente, entre otros. La suspensión del tabaco (56,57) y la ingesta de alcohol (20, 58-61), también han sido asociados a ganancia de peso en estudios epidemiológicos (Tablas 1 y 2).
Factores predictores de obesidad no metabólicos y factores gatillantes de ganancia de peso
| Predictores | Gatillantes |
|---|---|
| Padres obesos | Matrimonio reciente |
| Bajo nivel socioeconómico | Suspensión del tabaco |
| Estrés | Adicción a drogas |
| Sedentarismo | Exceso de alcohol |
| Macrosomía y obesidad desde la infancia | Tratamientos famacológicos |
| Embarazos múltiples | Tratamientos hormonales |
| IMC materno previo a embarazo | Inicio de vida laboral |
| Edad de menarquia | Suspensión de actividad deportiva |
Fármacos asociados conaumento del peso corporal
| AcciÓn FarmacolÓgica | FÁrmaco |
|---|---|
| Antipsicóticos | Clorpromazina, tioridazina, haloperidol, clozapina, olanzapina |
| Antidepresivos | Triciclicos, trazodona, litio, mirtazapina |
| Antiepilépticos | Valproato, carbamazepina |
| Dolor neuropático | Gabapentino, pregabalina |
| Antagonistas adrenérgicos | Terazosina, doxazosina, propanolol |
| Antihistamínicos | Ciproheptadina, hidroxicina, clorfenamina |
| Antidiabéticos | Sulfonilureas, insulina, glitazonas |
Aunque las enfermedades endocrinológicas son causas infrecuentes de obesidad, éstas deben descartarse antes de plantear un plan de tratamiento (25). En el hipotiroidismo, especialmente en casos severos, puede observarse algún incremento en los depósitos de grasa corporal por una disminución en el gasto energético, siendo una causa infrecuente de obesidad. El síndrome de Cushing origina una obesidad de tipo central, comprometiendo cara, cuello (“lomo de búfalo”), tronco, abdomen y acumulación de grasa visceral (mesenterio y mediastino). En niños se observa una obesidad generalizada con disminución del crecimiento lineal. Este síndrome se asocia con un mayor riesgo de enfermedades cardiovasculares, hipertensión arterial y diabetes. El síndrome de ovario poliquístico se asocia a obesidad en cifras variables (10-80% de los casos) y se caracteriza por anormalidades menstruales, infertilidad, hiperandrogenismo e hirsutismo, anormalidades hipotálamo-hipofisiarias (aumento de LH y prolactina) y marcada resistencia a la insulina, entre otras características. La deficiencia de hormona de crecimiento en adultos se asocia a una disminución de masa magra y aumento en la masa grasa corporal total y visceral, alteraciones que se revierten con la terapia de reemplazo hormonal.
Factores genéticos asociados a obesidadSe ha estimado que los factores genéticos explican entre un 25 y 40% de la etiología de la obesidad (62). Entre estos factores se pueden distinguir las alteraciones en genes que determinan obesidad, de aquellas que condicionan una susceptibilidad que se manifiesta cuando confluyen factores metabólicos o ambientales.
En relación a mutaciones de genes aislados se han identificado en seres humanos mutaciones en 11 diferentes genes, que han dado origen a casos de obesidad severa en niños y adultos. Las mutaciones más frecuentes se han encontrado en el gen del receptor 4 de melanocortina (MC4R), gen de leptina, gen del receptor de leptina, gen de proopiomelanocortina (POMC), gen de la convertasa 1 de prohormona y en el gen de SIM1, entre otros (63-66). Las enfermedades mendelianas corresponden a síndromes genéticos que dan origen a la obesidad, con características dismórficas especiales que facilitan su reconocimiento, y en su conjunto representan solo una pequeña proporción de niños obesos. Entre éstas se describen el síndrome de Prader-Willi, síndrome de Bardet-Biedl, síndrome de Alström, síndrome de Cohen, osteodistrofia hereditaria de Albright, síndrome de Bongiovanni, lipodistrofia familiar de Dunningan, lipodistrofia congénita de Berardinelli-Seip y síndromes de resistencia a la insulina, entre otros (65).
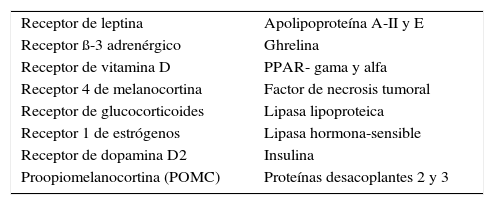
Mediante estudios de asociación se han identificado más de cien genes candidatos por su relación significativa con variables como peso, índice de masa corporal, porcentaje y distribución de masa grasa, niveles plasmáticos de leptina, gasto energético, lipólisis en adipocitos y variación de peso frente a distintas intervenciones. Las variaciones en la secuencia del ADN de hormonas, factores o receptores que se muestran en la Tabla 3 han sido asociadas con una o más de las variables señaladas.
Hormonas y receptores cuya variación en la secuencia de adn se asocia a obesidad o a factores de riesgo de ganancia de peso
| Receptor de leptina | Apolipoproteína A-II y E |
| Receptor ß-3 adrenérgico | Ghrelina |
| Receptor de vitamina D | PPAR- gama y alfa |
| Receptor 4 de melanocortina | Factor de necrosis tumoral |
| Receptor de glucocorticoides | Lipasa lipoproteica |
| Receptor 1 de estrógenos | Lipasa hormona-sensible |
| Receptor de dopamina D2 | Insulina |
| Proopiomelanocortina (POMC) | Proteínas desacoplantes 2 y 3 |
Los estudios de heredabilidad, usando estudios en familias de adoptados y en gemelos, han permitido estimar la importancia de los componentes genéticos en la etiología de la obesidad en humanos. La estimación de heredabilidad de fenotipos relacionados con obesidad fluctúa ente 6 y 85% en los diversos estudios: índice de masa corporal: 16-85%; circunferencia de cintura: 37-81%; relación cintura/cadera: 6-30%; porcentaje de masa grasa: 35-63% (65).
Junto con la culminación del Proyecto Genoma Humano y el desarrollo del Mapa Genético Humano (Internacional HapMap), se ha progresado en nuevas tecnologías que permiten analizar más de 1 millón de variantes genéticas en un solo ensayo. Uno de los hallazgos más importantes en los últimos años ha sido el descubrimiento del gen FTO (fat mass and obesity associated gene) y sus variantes alélicas (67). Es así como sujetos homocigotos para la variante alélica de riesgo, pesan 3kg. más que los homocigotos para la variante de bajo riesgo. Estas asociaciones han sido confirmadas en numerosos estudios en niños y adultos -caucásicos norteamericanos y europeos- y en población hispana, no solo con el índice de masa corporal y riesgo de obesidad, sino también con niveles plasmáticos de leptina, masa grasa total y subcutánea, y circunferencia de cintura. Aunque no se conoce con exactitud la función del gen FTO y su relación con el riesgo de obesidad, los estudios apuntan a que puede participar tanto en la regulación del apetito en el sistema nervioso central, como en la actividad lipolítica en el tejido adiposo (67). Otro importante hallazgo se originó en el mapeo del genoma completo de muestras de ADN obtenidas de los sujetos del Estudio del Corazón de Framingham y de otras importantes cohortes. En estos análisis, Herbert et al (68), encontraron una variante genética cercana al gen INSIG2 (insulin-induced gene 2), que estaba presente en cerca de 10% de los sujetos y que se asociaba a un exceso de peso de cerca de 2kg. por sobre los sujetos con alelos de bajo riesgo. La evidencia de estudios en animales muestra que la variante alélica de alto riesgo puede relacionarse con alteraciones en el metabolismo de triglicéridos.
Teoria infecciosa de la obesidad: adenovirus humano AD-36La presencia del AD-36 en el humano, en estado de portador sano, se ha asociado a una mayor masa grasa total y visceral, con menores niveles de colesterol y triglicéridos plasmáticos (69). En Estados Unidos la prevalencia de anticuerpos para AD-36 se observa en 30% de los obesos y en 11% de los no obesos. En otros países con baja prevalencia de obesidad, como Dinamarca y Bélgica, la presencia de AD-36 + es solo de 5% de la pobación adulta. Los mecanismos se postula que: promueve diferenciación de adipocitos y aumenta su replicación (Gen E4 de AD-36), aumenta la sensibilidad a la insulina, la captación de glucosa, reduce la expresión y secreción de leptina, induce la transcripción génica de enzimas lipogénicas (acción similar a glitazonas)
Microbiota intestinal y obesidadRatones obesos (ob/ob) tienen 50% menos Bacteroidetes y más Firmicutes que ratones delgados. La microbiota intestinal en ratones ob/ ob aumentaría la eficiencia en la absorción de calorías desde los alimentos. El transplante de microbiota intestinal desde ratones normales hacia ratones sin bacterias intestinales (germ-free) aumenta la masa grasa sin aumento en ingesta de alimentos, por lo que existe la posibilidad de que la composición de la microbiota intestinal afecte el monto de energía extraído desde la dieta (70-72). En humanos sometidos a dieta hipocalórica baja en carbohidratos o baja en grasas durante 1 año, se ha observado una cambio en la proporción de las distintas especies en la microbiota intestinal, aumentando la presencia de Bacteroidetes en correlación con la pérdida de peso (73). Sin embargo otros estudios han fallado en encontrar diferencias en la proporción de Bacteroidetes/Firmicutes entre sujetos delgados, con sobrepeso y obesos (74). Hasta la fecha no ha podido establecerse si en humanos existe una relación entre el patrón que adopta la microbiota intestinal y la predisposición hacia la obesidad.
SíntesisEl conocimiento de los factores de riesgo de obesidad es relevante, ya que permite elaborar estrategias preventivas en poblaciones e individuos suceptibles. Este conocimiento nos permite concordar y simpatizar, al menos parcialmente, con la visión de los pacientes de que su predisposición a ganar peso no es de su completo control. El tratamiento de pacientes obesos con predisposición a recuperar el peso reducido impone un mayor énfasis en los cambios conductuales para mejorar los resultados en el largo plazo.
Los autores declaran no tener conflictos de interés, en relación a este artículo.