




Las levaduras, durante el proceso de elaboración de cerveza, producen más de 500 compuestos químicos; estos pueden impactar tanto negativa como positivamente en las características organolépticas de la cerveza. En los últimos años, y en particular gracias al avance de la biología molecular y la genómica, se han logrado progresos notables en el conocimiento de las bases moleculares y celulares de la síntesis y regulación de muchos de estos compuestos que inciden en lo que se denomina flavor (aroma y sabor) de la cerveza. Este artículo está enfocado en los ésteres responsables del aroma y el sabor floral y frutado de la cerveza. La formación de estos ésteres depende de diversas enzimas y de factores como la concentración de nutrientes presente en el mosto, la cantidad de oxígeno y dióxido de carbono disuelto, la temperatura de fermentación y, principalmente, la genética de la levadura utilizada. En esta revisión se brinda información de cómo se originan los ésteres y cómo los diferentes parámetros fermentativos impactan en las concentraciones finales de estos compuestos y en la calidad del producto terminado.
During brewing process yeast produce more than 500 chemical compounds that can negatively and positively impact beer at the organoleptic level. In recent years, and particularly thanks to the advancement of molecular biology and genomics, there has been considerable progress in our understanding about the molecular and cellular basis of the synthesis and regulation of many of these flavor compounds. This article focuses on esters, responsible for the floral and fruity beer flavor. Its formation depends on various enzymes and factors such as the concentration of wort nutrients, the amount of dissolved oxygen and carbon dioxide, fermentation temperature and mainly the genetics of the yeast used. We provide information about how the esters originate and how is the impact of different fermentative parameters on the final concentrations of these compounds and the quality of the end product.
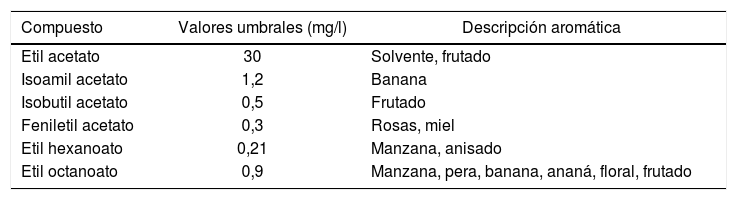
Durante la producción de cerveza, junto con la formación de dióxido de carbono y etanol, se genera una amplia variedad de compuestos de vital importancia para lo que comúnmente se conoce como flavor, es decir, el sabor y el aroma. Entre ellos, los ésteres conforman el grupo de compuestos que inciden de manera significativa en la calidad sensorial del producto final42,57,66. En cerveza se han detectado más de 100 ésteres diferentes14,45. Los más relevantes, generados por las levaduras a través de su metabolismo secundario, pueden dividirse en dos grupos principales: ésteres de acetato, tales como etil acetato, isoamil acetato y feniletil acetato, y ésteres de etilo, como etil hexanoato y etil octanoato. Ambos grupos en conjunto son deseables en la mayoría de las cervezas del tipo Ale (cervezas de fermentación alta), dado que aportan aromas y sabores florales/frutales y contribuyen a su característica complejidad de aroma y sabor. Mayormente, estos ésteres se encuentran en concentraciones cercanas al nivel umbral, lo cual implica que pequeños cambios en su concentración podrían afectar el aroma y sabor de la cerveza (tabla 1). Algunos ésteres en concentraciones muy altas aportan aromas y sabores indeseados (p.ej., acetato de etilo: solvente). Además, la presencia de diferentes ésteres puede tener un efecto sinérgico e influir en la percepción sensorial general, a pesar de que sus concentraciones individuales estén por debajo del umbral18,84. Es por esto que los niveles de estos compuestos deben mantenerse dentro de ciertos límites de concentración, a fin de evitar desbalances que den lugar a cervezas con características indeseables11,22,36. De aquí surge la necesidad de entender y controlar su formación, para brindar productos con características reproducibles que cumplan con las exigencias y preferencias de los consumidores.
Ésteres más relevantes en el flavor de la cerveza sintetizados por las levaduras
| Compuesto | Valores umbrales (mg/l) | Descripción aromática |
|---|---|---|
| Etil acetato | 30 | Solvente, frutado |
| Isoamil acetato | 1,2 | Banana |
| Isobutil acetato | 0,5 | Frutado |
| Feniletil acetato | 0,3 | Rosas, miel |
| Etil hexanoato | 0,21 | Manzana, anisado |
| Etil octanoato | 0,9 | Manzana, pera, banana, ananá, floral, frutado |
Fuente: Engan14.
Los ésteres generados por las levaduras se producen por reacción enzimática entre un ácido orgánico y un alcohol. Investigaciones en el campo de la genética y proteómica de levaduras han permitido conocer más en detalle las propiedades de los genes y las enzimas involucradas en la biosíntesis de ésteres, así como también su regulación frente a diferentes factores33,34,53,74,85. En el presente trabajo se realizará una revisión sobre el papel de cada una de estas enzimas en la síntesis de ésteres de acetato y de etilo durante la elaboración de cerveza. Asimismo, se describirá cómo varía la actividad de dichas enzimas y la concentración de estos compuestos frente a diferentes parámetros tecnológicos asociados a las condiciones de fermentación, la composición del mosto y las características de las levaduras. Se espera que la información presentada a continuación ayude a comprender en profundidad el proceso de formación de los ésteres por parte de las levaduras y sirva de guía, sobre todo para los productores de cerveza, al momento de aplicar estrategias de control y ajuste del sabor y aroma en sus cervezas.
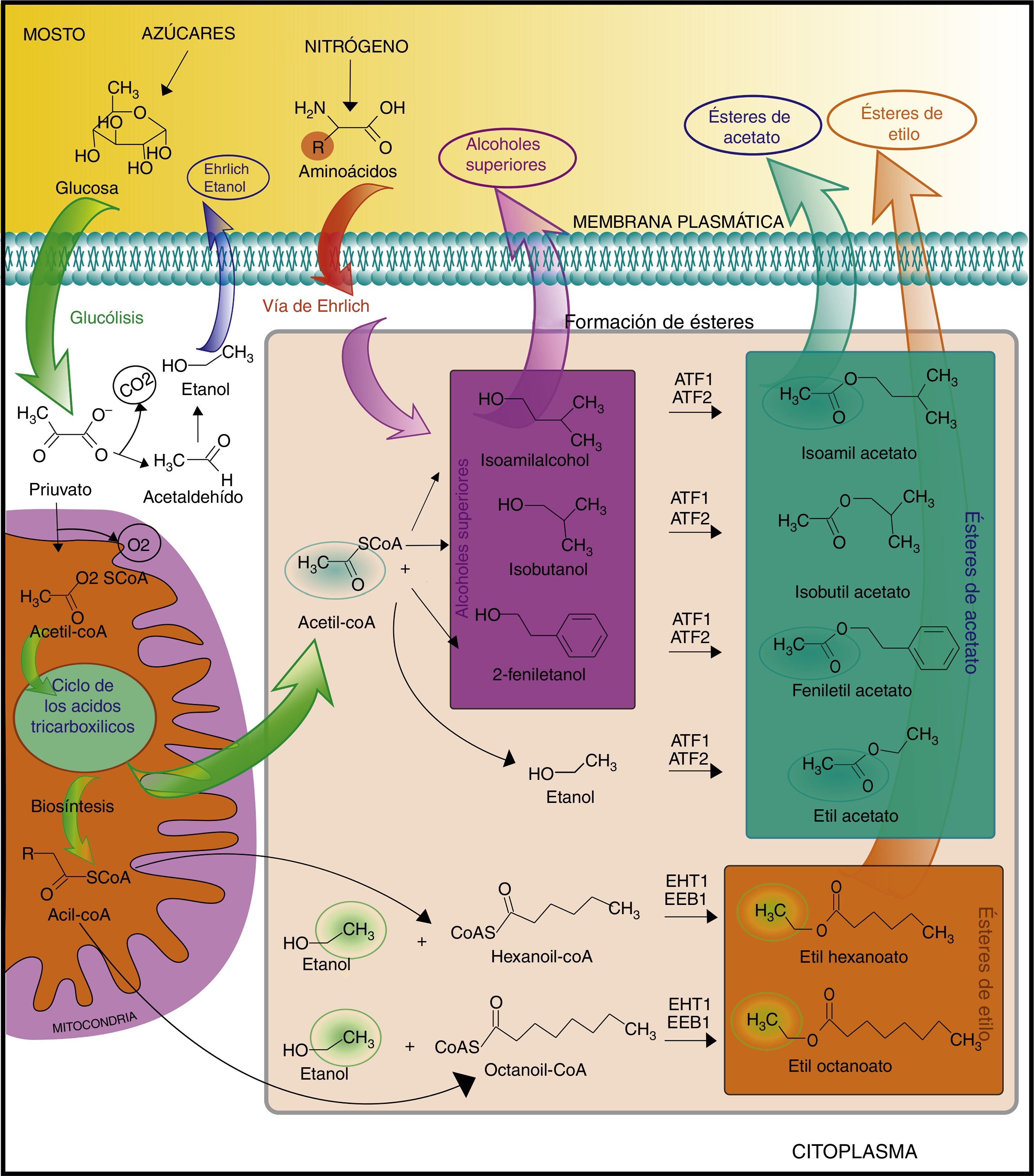
¿Cómo se generan los ésteres?La formación de ésteres se produce durante la fermentación primaria y se encuentra altamente asociada al metabolismo de lípidos y al crecimiento de las levaduras. Estos compuestos son sintetizados en el citoplasma a partir de reacciones catalizadas por enzimas acil transferasas (o éster sintasas), en las que se requiere acil-CoA como cosustrato42,64. La mayor parte del acil-CoA requerido es generado por descarboxilación oxidativa del piruvato, dando lugar a acetil-CoA, mientras que el resto del acil-CoA es formado por la acilación entre ácidos grasos y coenzimaA (CoA) libre60 (fig. 1).

En ausencia de oxígeno, la reacción entre acetil-CoA y un alcohol (etanol o alcoholes superiores) permite la formación de ésteres de acetato, como etil acetato, isoamil acetato, isobutil acetato y feniletil acetato, mientras que la combinación entre las largas cadenas de acil-CoA y etanol produce etil ésteres57,60. Entre estos últimos, el etil hexanoato y el etil octanoato son de gran importancia para la obtención de cervezas balanceadas en cuanto a perfiles aromáticos.
Una vez sintetizados dentro de la célula, por su carácter lipofílico, los ésteres pueden difundir a través de la membrana celular para ser liberados al medio de fermentación. A diferencia de los ésteres de acetato, que son excretados rápidamente y casi en su totalidad, la transferencia al medio de los ésteres de etilo disminuye a medida que aumenta el tamaño de esa molécula84. El etil hexanoato es liberado en un 100%, mientras que este porcentaje puede llegar hasta un 68 y un 17% para el etil octanoato y el etil decanoato, respectivamente.
Los primeros modelos sobre la síntesis de ésteres se basaron en la disponibilidad de sustratos como factor determinante para su formación9,78. Sin embargo, varios trabajos demostraron que las enzimas que catalizan la formación de estos compuestos también cumplen un rol central.
Enzimas y genes involucrados en la formación de ésteres de acetatoLas enzimas alcohol acetil transferasasI yII (AATasaI yII), codificadas por los genes atf1 y atf2, respectivamente, son las responsables de la formación de ésteres de acetato (fig. 1). Los genes atf1 y atf2 han sido identificados en el genoma de levaduras de tipo Ale (Saccharomycescerevisiae) y Lager (Saccharomyces.pastorianus)15,25,67,90. Las levaduras de tipo Lager son híbridos en los que una parte de su genoma proviene de S.cerevisiae y la otra de una especie denominada Saccharomyces.eubayanus, la cual se encuentra adaptada al frío4,38. Por estas características, las levaduras Lager pueden presentar dos genes por cada AATasa, dos correspondientes al subgenoma de S.cerevisiae (denominados genes atf1 y atf2) y el resto al subgenoma de S.eubayanus (genes Lgatf1 y Lgatf2).
Existe una fuerte correlación entre la expresión (proceso donde se transforma la información genética en proteínas y enzimas) de estos genes y los niveles de ésteres de acetato producidos65,85,91,96. Particularmente, en ensayos en los que se aumentó la expresión de genes atf1, atf2 y Lgatf1 en levaduras cerveceras hubo un incremento significativo en la producción de etil acetato e isoamil acetato, lo que sugiere que los genes atf son relevantes en la síntesis de estos compuestos85,96.
Verstrepen et al.85 también observaron que atf1 y atf2 participan, al menos parcialmente, en la generación de otros ésteres de acetato, entre los cuales se encuentran el isobutil acetato y el feniletil acetato. No obstante, el gen atf2, al igual que el gen Lgatf1, tendría un rol menor respecto de atf1. Por otro lado, las deleciones realizadas en los genes atf1 y atf2 prácticamente no permitieron la formación de isoamil acetato ni de feniletil acetato, lo que demuestra que las enzimas codificadas por ambos genes son responsables de la totalidad de la actividad alcohol acetil transferasa involucrada en la generación de ambos compuestos. Además, se ha demostrado que dichas deleciones solo reducen la producción de etil acetato, propil acetato e isobutil acetato hasta un 50%39,40,85, lo cual indica que en la formación de estos ésteres intervienen otras enzimas éster sintasas, aún desconocidas.
Pocos trabajos se encuentran orientados al análisis de la expresión de los genes ATTasa durante fermentaciones realizadas en mosto cervecero y a escala semipiloto utilizando levaduras comerciales sin modificar25,65. A pesar de ello, en dichos estudios se encontró que las mayores concentraciones de ésteres de acetato podrían explicarse por los niveles máximos de expresión de los genes atf1 y atf2 (niveles de Atf2p menores que Atf1p), en concordancia con los resultados obtenidos a partir de la modificación (sobreexpresión y deleción) de estos genes.
La concentración final de los ésteres de acetato también se encuentra influenciada por la presencia de esterasas, un grupo de enzimas hidrolasas que catalizan la ruptura de ésteres o evitan su formación. En particular, la enzima IAH1p cumple un rol relevante en la hidrólisis de isoamil acetato, etil acetato, isobutil acetato y feniletil acetato17,41,97. La cantidad de estos ésteres en sake puede aumentar hasta dos veces cuando se realizan fermentaciones con cepas de levadura comerciales deficientes en el gen iah117. Acorde con estos resultados, la sobreexpresión del gen iah1 en la levadura vínica VIN13 también permitió generar vinos con cantidades reducidas de etil acetato, feniletil acetato, isoamil acetato y hexil acetato41.
En diferentes cepas de levaduras utilizadas comúnmente en la producción de cervezas de trigo, la expresión de iah1 varía considerablemente, observándose niveles máximos de expresión luego de 36h de iniciada la fermentación primaria70. Asimismo, el aumento en la expresión de iah1 en dichas cepas se correlacionó con la disminución en la concentración de ésteres de acetato. Esto indica la importancia de Iah1p en la obtención de niveles balanceados de estos compuestos.
Ésteres de etiloEn cerveza, los ésteres de etilo se encuentran en menor proporción que los ésteres de acetato. Las enzimas que catalizan la formación de ésteres de etilo son las denominadas acil-CoA/etanol o-aciltransferasas. Malcorps y Dufour39 fueron los primeros en brindar evidencias bioquímicas sobre la presencia de enzimas etil alcohol acil transferasas en S.cerevisiae. Sin embargo, recién en 2006 fue posible estimar el papel que cumplen dos de ellas, codificadas por los genes eht1 y eeb168. Mediante ensayos de deleción de ambos genes utilizando una cepa de S.cerevisiae, Saerens et al.68 mostraron que estas enzimas contribuyen a la síntesis de etil butanoato, etil hexanoato y etil decanoato. Por otro lado, la presencia de actividad éster sintasa residual luego de la doble deleción de estos genes sugirió que otras etil éster sintasas —aún desconocidas— estarían involucradas en la formación de dichos compuestos.
En cuanto a la preferencia de sustratos utilizados por Eeb1p y Eht1p, y por ende a su capacidad de producir uno u otro éster de etilo, diferentes estudios han revelado resultados contradictorios. Proteínas recombinantes Eeb1p y Eht1p purificadas a partir de Escherichia coli mostraron preferencia por sustratos de cadena larga (C8) y cadena corta (C4), respectivamente68. Sin embargo, en el mismo trabajo esto no se correspondió con lo observado en fermentaciones, donde se utilizó una cepa de S.cerevisiae con deleciones en el gen eeb1. En este caso se produjo una reducción moderada de la concentración de etil octanoato (45%) respecto de la disminución en los niveles de etil hexanoato (88%). Asimismo, la deleción del gen eht1 dio lugar a niveles de etil butanoato similares a los generados en fermentaciones realizadas con la levadura control (sin modificar), y solo redujo las concentraciones de etil hexanoato y etil octanoato en un 36 y un 20%, respectivamente68. Recientemente se logró purificar y caracterizar la enzima EHT1p, pero utilizando una levadura como sistema de expresión. Su sustrato de preferencia fue octanoil-CoA (C8)31. No obstante, se necesitan más ensayos a fin de clarificar la especificidad tanto de Eht1p como de Eeb1p en la formación de diferentes ésteres de etilo relevantes para el aroma y el sabor de las cervezas.
Ensayos de sobreexpresión de eeb1 y eht1 en los que no se observaron incrementos en la producción de ésteres de etilo dieron indicios de que las enzimas codificadas por estos genes, además de poseer actividad sintasa, tendrían la capacidad de hidrolizar ésteres de etilo68. De hecho, ambas enzimas presentan un dominio α-β hidrolasa en su extremo C terminal31. Particularmente, en Eht1p se observó actividad tioesterasa con capacidad de hidrolizar acil-CoA de cadena media para generar ácidos grasos libres31. Ello refuerza una de las hipótesis previas acerca del principal rol metabólico de enzimas aciltransferasas: recuperar CoA libre a partir de acil-CoA de cadena media, compuestos que en condiciones anaeróbicas se acumulan en el interior de las levaduras67. A pesar de que en los últimos años se realizaron avances acerca de la actividad éster sintasa e hidrolasa presente en Eht1p y Eeb1p, aún existen interrogantes acerca de la regulación y el balance de ambas actividades durante la producción de ésteres de etilo.
Factores que afectan la formación de ésteresAzúcares del mostoUn mosto estándar contiene aproximadamente un 90% de hidratos de carbono, entre los cuales se encuentran dextrinas y azúcares como sacarosa, fructosa, glucosa, maltosa y maltotriosa7. La concentración y composición en mosto de estos hidratos de carbono, así como la manera en que son utilizados por las levaduras, tienen una influencia directa en su metabolismo y, por lo tanto, en el perfil aromático del producto final. Una práctica común en grandes cervecerías es la producción de cerveza a partir de mostos de alta densidad, es decir, con alta concentración de azúcares, a fin de aumentar la productividad en volumen de cerveza sin aumentar significativamente los gastos energéticos58. La desventaja de esta técnica es que el metabolismo de las levaduras se ve modificado de manera significativa. Varios estudios confirmaron la sobreproducción de ésteres de acetato en medios con alta concentración de hidratos de carbono, dando lugar a cervezas demasiado frutadas y con un notable aroma a solvente (como, por ejemplo, acetona) si no se mantiene un balance adecuado en la oxigenación y contenido de nitrógeno en el mosto5,35,43,95. El incremento en el contenido de hidratos de carbono en mostos utilizados en fermentaciones con levaduras de tipo Lager y Ale puede producir un 25 y un 45% de aumento en los niveles de etil acetato, respectivamente66.
Para generar mostos de alta densidad es frecuente adicionar jarabes, por lo que además se debe tener en consideración su composición de azúcares. Dependiendo del azúcar adicionado es posible generar más o menos compuestos del aroma y sabor. Younis y Stewart94 reportaron que al utilizar maltosa como única fuente de carbono en medio sintético, los niveles de ésteres como isoamil acetato y etil acetato generados por levaduras Ale y Lager son menores en comparación con medios que contienen únicamente fuentes de carbono más asimilables, como glucosa o fructosa. Posteriormente, los mismos autores demostraron que mostos de alta densidad con un 30% de jarabe de alta maltosa producen una cantidad de ésteres de acetato y alcoholes superiores significativamente menor en comparación con mostos de alta densidad de pura malta93. Estos resultados concuerdan con los obtenidos por Piddocke et al.56, quienes observaron que mostos de 21 y 24 °P (correspondientes a densidades iniciales de 1,0874 y 1,1011, respectivamente) suplementados con maltosa provocan una disminución de etil acetato e isoamil acetato respecto de mostos de la misma densidad en los que se adicionó glucosa. La adición de sacarosa, aunque posee un efecto estimulante en la expresión del gen atf1, genera menores concentraciones de ésteres de acetato en relación con el agregado de maltosa. Levaduras expuestas a altas concentraciones de sacarosa presentan mayor estrés osmótico que en mostos de otra composición. Es por esto que se aconseja evitar altos niveles de este azúcar con el fin de obtener mostos de alta densidad11.
Varias razones podrían explicar el efecto de los diferentes azúcares asimilables en la producción de ésteres. Una de ellas es que la fermentación con maltosa podría inhibir el transporte de compuestos volátiles hacia el exterior de la célula, posiblemente por alteración de la membrana plasmática94. Otra posibilidad es que, como resultado del metabolismo de la maltosa, se produzcan menores niveles de acetil-CoA y, por lo tanto, concentraciones menores de ésteres de acetato por falta de sustrato56. En el mismo sentido, Younis y Stewart94 sugirieron que altos niveles de glucosa incrementan la producción de acetil-CoA y de ésteres de acetato. Más aún, la expresión de los genes atf1 y atf2 es inducida por glucosa a través de una cadena de señales (vía AMPc/PKA) conectada a la proteína Sch9p82. Esta última proteína no solo responde a la presencia de glucosa, sino también a los niveles de nitrógeno y fósforo en el medio67,77,83, por lo que estos factores también influirían en la expresión de Atf1.
Efecto de la fuente de nitrógenoLa fuente de nitrógeno es de gran importancia para el desarrollo de las levaduras, dado que cumple un papel crucial en la formación de proteínas. El malteado y macerado permite, a partir del material nitrogenado presente en la cebada, liberar al mosto entre 700 y 800mg de nitrógeno por litro. Dependiendo de las condiciones de producción, la fracción soluble de nitrógeno en el mosto está formada por un 20% de proteínas, un 22% de polipéptidos y un 58% de péptidos y aminoácidos libres, aproximadamente6; estos últimos son la principal fuente asimilable de nitrógeno52,63. Generalmente se recomiendan concentraciones de aminoácidos libres del orden de los 150-200mg/l de mosto para lograr cervezas de buena calidad28.
En el mosto, la concentración de compuestos nitrogenados afecta tanto al crecimiento de las levaduras como a la generación de compuestos volátiles responsables del aroma y sabor durante la fermentación10,59,86. El aumento en las concentraciones de nitrógeno asimilable puede incrementar la expresión de genes atf136. La velocidad de formación y los niveles finales de ésteres de acetato también se ven incrementados al aumentar las cantidades de nitrógeno asimilable desde 115mg/l hasta 433mg/l, aunque solo cuando se utilizan mostos con densidades iniciales de 12 °P23,36. En mostos más concentrados (>16 °P), un aumento similar en la fuente de nitrógeno puede provocar una disminución en la concentración de compuestos del aroma y sabor, tales como etil acetato y feniletil acetato36,37. En mostos de alta densidad, las levaduras necesitan asimilar una cantidad extra de nitrógeno para hacer frente a las condiciones de estrés55. Asimismo, una relación alta de la fuente de carbono con respecto a la de nitrógeno (C:N) genera desbalances de nutrientes, que pueden impactar en la formación de compuestos aromáticos71,72. En cuanto a los ésteres de etilo, el aumento en los niveles de nitrógeno no modifica sus concentraciones, independientemente de la densidad del mosto36,37,66.
El perfil aromático de las cervezas no solo se ve influenciado por la cantidad de nitrógeno asimilable, sino también por el tipo de aminoácidos presentes en el mosto. La incorporación de aminoácidos como leucina, isoleucina o valina y su metabolismo a través de la vía de Ehrlich provocan, respectivamente, un aumento en la cantidad de isoamil alcohol, 2-metil butanol, isobutanol y sus correspondientes ésteres de acetato24,59,61,73. Específicamente, al utilizar la levadura Safale-S04, la adición de 0,750g/l de L-leucina conduce a un incremento de un 41% en la concentración total de ésteres de acetato comparado con el control sin suplemento26. En cambio, las concentraciones de ésteres de etilo pueden disminuir o no verse afectadas por el suplemento de aminoácidos, posiblemente por las modificaciones intracelulares en la concentración de acetil-CoA26,61.
La prolina y la histidina son otros aminoácidos de relevancia para el aroma y sabor de las cervezas34,59. La adición de prolina en medio sintético permite la producción de mayores concentraciones de etil acetato en levaduras de tipo Ale y Lager. Este aminoácido no puede ser metabolizado por la vía de Ehrlich para dar lugar a un alcohol superior que sirva de precursor para la formación de ésteres, con lo cual su aporte a la formación de aromas en cervezas podría explicarse a través de la síntesis de glutamato, metabolito que sí interviene en dicha vía19. Por otra parte, Lei et al.34 observaron que el suplemento de histidina en mostos de alta densidad activa genes codificantes de permeasas (Bap2, Lyp1, Hip1 y Agp1) específicas del transporte de aminoácidos durante la fase exponencial de levaduras Lager. En ese mismo trabajo se observó un incremento significativo en la producción de isobutanol, isoamil alcohol, feniletanol y etil acetato.
La temperatura de fermentaciónLa razón por la cual la síntesis de ésteres de acetato y de etilo depende de la temperatura aún se desconoce con exactitud. En general, se ha observado que la concentración final de ésteres en cervezas se ve incrementada al utilizar mayores temperaturas de fermentación, aunque algunas cepas de levaduras pueden mostrar otros comportamientos en este sentido54,65,79. Por ejemplo, en levaduras Lager, cuando se modifica la temperatura de fermentación de 12 a 15°C, las concentraciones de ésteres como el isoamil acetato y el feniletil acetato pueden llegar hasta 1,73mg/l (aumento aproximado del 40%) y 0,66mg/l (aumento del 12%), respectivamente65, mientras que el total de ésteres de etilo puede aumentar en un 15%. Sin embargo, en levaduras Ale se observó solo aumento en la concentración de feniletil acetato (17%) cuando se elevó la temperatura de 20 a 24°C.
Si bien un aumento de temperatura genera una mayor concentración de alcoholes superiores —compuestos necesarios para la síntesis de ésteres26,33,66—, varios estudios sugieren que la temperatura afecta principalmente la actividad de las enzimas que intervienen en su formación o la biosíntesis de aquellas. Saerens y colaboradores observaron que el aumento en la temperatura de fermentación incrementa los niveles de expresión de los genes atf1 y atf265, lo que confirma resultados anteriores42. En cepas industriales de tipo Ale y Lager, la expresión del gen eeb1 también se incrementa a temperaturas de fermentación mayores65. No obstante, la falta de correlación entre la expresión de eeb1 y la concentración de ésteres de etilo al realizar fermentaciones con cepas Ale sugiere que la temperatura y su efecto en la expresión de eeb1 no son factores primarios en la producción de estos compuestos65,66. Como se mencionó anteriormente, tanto las enzimas Eeb1p como Eht1p poseen actividad esterasa, por lo que pueden evitar incrementos en las concentraciones de ésteres de etilo. Por otro lado, la adición de ácidos grasos de cadena larga, que son precursores para la formación de ésteres de etilo, permite incrementar tanto los niveles de expresión de eeb1 como la producción de estos ésteres66,68. Esta observación indica que la concentración de sustratos puede ser un factor limitante en la síntesis de estos compuestos.
Si bien la temperatura de fermentación no es determinante en la síntesis de ésteres de etilo, algunos autores sostienen que la liberación de estos compuestos se vería facilitada con los aumentos de temperatura, por favorecer la eficiencia de excreción o la autolisis de las levaduras64,75.
Oxigenación y ácidos grasos insaturadosLa presencia de oxígeno y de ácidos grasos insaturados en el mosto causa un efecto negativo en la producción de ésteres13,51,81. Antes se sostenía que la reducción de ésteres de acetato en presencia de ácidos grasos insaturados se debía a la baja disponibilidad de acetil-CoA, dado que, en presencia de oxígeno y junto con los ácidos grasos, el acetil-CoA es requerido en la formación de fosfolípidos y triglicéridos, compuestos necesarios para el crecimiento de las levaduras78. Varios autores luego observaron que la formación de ésteres en presencia de oxígeno y ácidos grasos insaturados depende esencialmente de la enzima AATasa16,92. El gen codificante de esta enzima, al igual que un subgrupo de genes hipóxicos, posee una región promotora con un sitio de unión al complejo Rox1p-Tup1-Ssn6. En condiciones aeróbicas, la transcripción del gen Rox1 se activa y su producto es capaz de asociarse al complejo represor Tup1-Ssn6, lo que permite su unión al promotor atf1 y la inhibición de la transcripción de atf142. Sin embargo, Vasconcelles et al.80 revelaron posteriormente que dicho mecanismo juega un rol menor en la regulación de atf1 y que la principal respuesta frente al oxígeno depende del elemento de respuesta bajo en oxígeno (conocido por la sigla LORE [low oxygen response element]), ubicado en la secuencia promotora de atf1. La inducción mediada por LORE no se encuentra completamente caracterizada, pero se ha demostrado que este mecanismo depende del procesamiento de dos proteínas de membrana del retículo endoplasmático, Spt23p y Mga2p30. En la represión de atf1 generada por los ácidos grasos insaturados también interviene el LORE80, aunque aún se desconoce el mecanismo de represión.
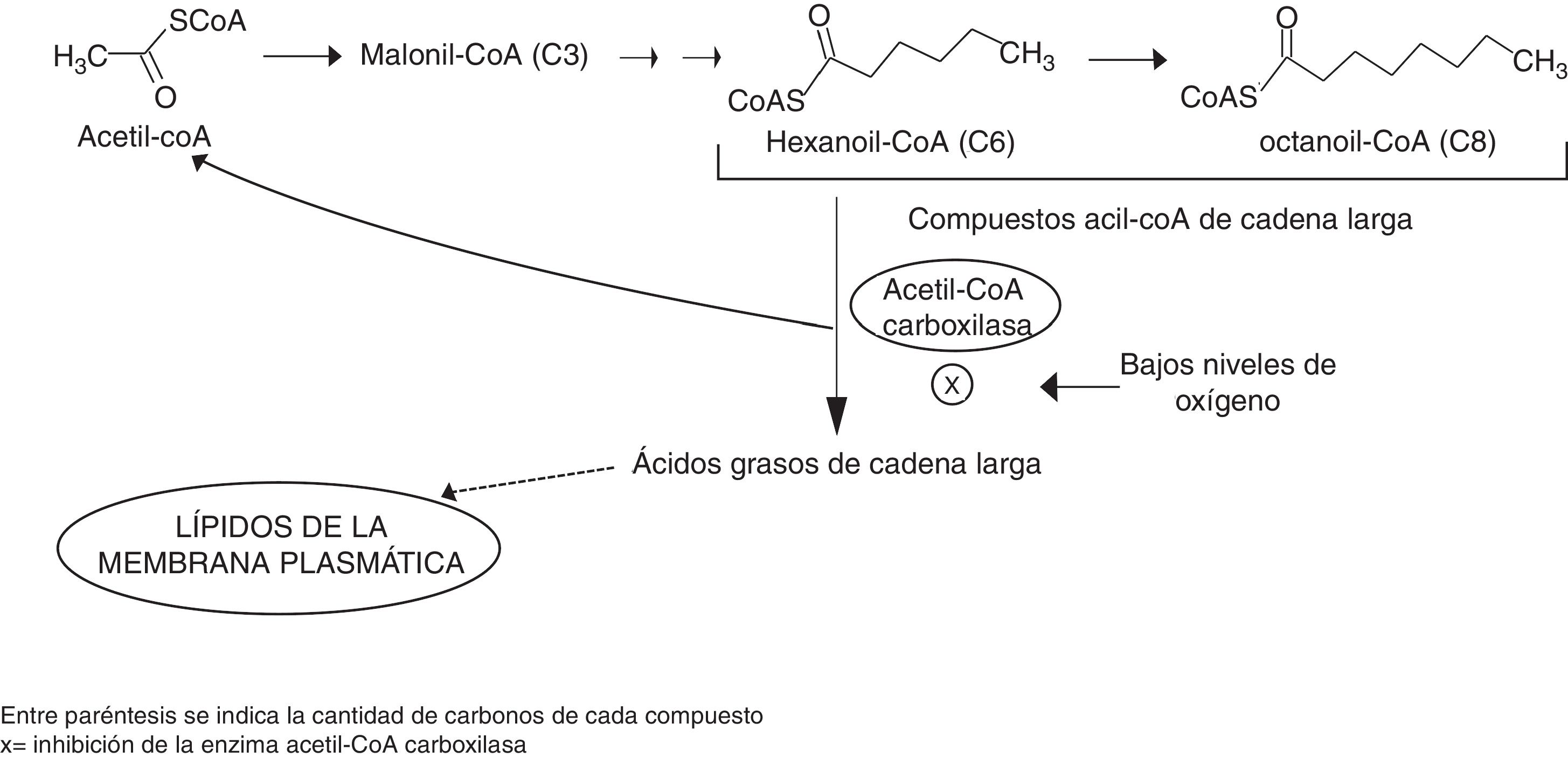
Debido a que las evidencias reunidas hasta el momento indican que la formación de los ésteres de etilo no depende tanto de la actividad de las enzimas Eeb1p y Eht1p como de la disponibilidad de sustratos60,66, la inhibición de este proceso en presencia de oxígeno y ácidos grasos insaturados puede explicarse mediante el modelo planteado por Dufour et al.13. De acuerdo con este modelo, la activación de la enzima acetil-CoA carboxilasa, —clave en la regulación de la biosíntesis de ácidos grasos— es determinante para que el complejo proteico ácido graso sintasa libere ácidos grasos de cadena larga.
En condiciones limitadas de oxígeno se inhibe la acetil-CoA carboxilasa y se acumulan compuestos acil-CoA de cadena larga, lo cual favorece la síntesis de ésteres de etilo (fig. 2). En presencia de oxígeno, se sintetizan ácidos grasos insaturados, se suprime la inhibición de esta enzima y la reacción de elongación procede para formar nuevamente ácidos grasos de cadena larga. Como resultado, el pool intracelular de acil-CoA de cadena larga se reduce y queda menos disponible para la formación de ésteres de etilo13. Esto sugiere que en diferentes condiciones de fermentación la actividad de las enzimas que intervienen en la síntesis de ácidos grasos podría ser un blanco para la modificación de los niveles de ésteres de etilo en cerveza66.

La adición de lípidos al mosto, especialmente de ácidos grasos insaturados, puede eliminar los requerimientos de aireación al inicio de las fermentaciones76. No obstante, los ácidos grasos insaturados aún pueden inhibir la producción de ésteres. Moonjai et al.50 investigaron el efecto de la adición de estos lípidos en levaduras antes de ser agregadas al mosto. Los resultados mostraron que cuando las levaduras tratadas previamente con ácidos grasos insaturados son agregadas en mostos poco oxigenados, no hay reducción en los niveles de ésteres de acetato. Además, tanto el crecimiento de estas levaduras como su perfil de atenuación son similares a los obtenidos con levaduras no tratadas cuando se inoculan en mostos preaireados. Estas observaciones fueron aplicadas con éxito a escala industrial utilizando aceite de oliva para el tratamiento previo de las levaduras, dado que contiene ácidos grasos insaturados y es de bajo costo29. Las cervezas producidas en este caso presentaron mayores concentraciones de ésteres respecto de los controles y una mejoría significativa en su aroma y sabor. Sin embargo, cabe considerar que el aceite de oliva no es fuente de esteroles, componentes esenciales de las membranas plasmáticas requeridos para la generación de nuevas levaduras. Una cantidad reducida de oxígeno en el mosto, además de evitar la inhibición de la formación de ésteres, evita la oxidación de compuestos como polifenoles, iso-α-ácidos y alcoholes superiores y, por lo tanto, el deterioro organoléptico de las cervezas producidas12.
Presión y dióxido de carbono disueltoUn exceso en la cantidad de dióxido de carbono disuelto generalmente inhibe el crecimiento de las levaduras y su metabolismo, posiblemente por la inhibición de las reacciones de descarboxilación, vitales para la célula33,44. Como consecuencia, también se reduce de manera significativa la producción de ésteres y alcoholes superiores, aunque en estos últimos el efecto es menor21,79. Dado que las reacciones de descarboxilación son necesarias para la producción de acetil-CoA, se cree que la influencia del dióxido de carbono en la concentración final de ésteres se debe a la inhibición de la síntesis de sustratos necesarios para la generación de estos compuestos. Al realizar fermentaciones a gran escala, el dióxido disuelto puede llegar a grandes concentraciones. Por su mayor altura, los fermentadores utilizados en este tipo de fermentaciones generan una mayor presión hidrostática, lo cual favorece la disolución del dióxido de carbono.
A pesar de que una disminución en la concentración de ésteres usualmente es indeseable, cuando se utilizan temperaturas de fermentación elevadas y mostos con alta densidad inicial se puede aplicar dióxido de carbono para controlar la formación de compuestos responsables del aroma y sabor en cervezas2. Cuando se eleva la temperatura de fermentación a presión atmosférica se produce un rápido y mayor crecimiento de las levaduras, lo que hace que la fase de adaptación de las levaduras al medio (fase lag) se reduzca o prácticamente desaparezca33. En esas condiciones, la velocidad de producción de ésteres de acetato también aumenta y puede alcanzar su pico máximo alrededor de los 50min de iniciada la fermentación. Para evitar concentraciones muy elevadas de ésteres a altas temperaturas de fermentación es posible incrementar la presión de dióxido luego de 25-30h. Esto permite que luego de la fase inicial, al aplicar presión, el crecimiento de las levaduras se enlentezca y la producción de ésteres disminuya33,44. Una fórmula empírica útil para estimar la presión apropiada está dada por esta ecuación: P (bar)=°C/10, donde P es la presión en bares y °C es la temperatura de fermentación44. Por ejemplo, si una levadura Lager es utilizada en fermentaciones a 20°C aproximadamente, la presión puede elevarse hasta alrededor de 2bar. El dióxido de carbono disuelto a una presión de 2bar se duplica y las concentraciones de etil acetato, isoamil acetato, isobutil acetato, etil hexanoato y etil octanoato disminuyen cerca de un 50% respecto de fermentaciones sin presurizar33,62.
La levadura: factor determinante en el perfil aromático de las cervezasUno de los factores más importantes que afecta a la producción de ésteres durante la producción de cerveza es, sin duda, la cepa de levadura utilizada. El diferente aroma y sabor generado por un tipo de levadura respecto de otro —en vinculación directa con los ésteres producidos— puede deberse a diferencias interespecíficas en términos genómicos, en los mecanismos de regulación de la expresión de los genes responsables de la síntesis de dichos ésteres y en la actividad de las enzimas participantes25,27,32,74. Tanto mutaciones puntuales en zonas regulatorias o en genes estructurales85 como mutaciones de mayor envergadura (deleciones, duplicaciones, aneuploidías, poliploidías) son responsables de la diversidad fenotípica de las cepas de levaduras cervecera8,32,46,89. En particular en las levaduras Lager (que son producto de la hibridación de dos especies de levadura sacaromicéticas38), la situación es aún más compleja como resultado de la interacción molecular entre los dos subgenomas.
Por ejemplo, He et al.25 demostraron que en el transcurso de fermentaciones realizadas a 9,5°C utilizando un mosto inicial de 13°P y una levadura Lager industrial, la transcripción de los genes atf1, atf2, eht1 y iah1 es dominada por la parte no S.cerevisiae. Por otro lado, la sobreexpresión de genes atf1 y atf2 derivados de levaduras de tipo Ale y Lager puede generar distintas concentraciones de cada éster, aun utilizando las mismas condiciones de fermentación. Esto sugiere que las variaciones en el perfil aromático producido por las levaduras podrían obedecer a mutaciones específicas en sus genes atf85.
La formación de ésteres también podría verse influenciada por el nivel de ploidía (cantidad de juegos de cromosomas) en las levaduras, ya que un incremento en el número de copias de un gen generalmente origina un aumento de su expresión y fenotipos diferentes8,32,46,89. Cepas de levaduras Lager pertenecientes al grupoI (Saaz), que tienen aproximadamente tres juegos de cromosomas (alotriploide), generan menores cantidades de ésteres (como isoamil acetato, etil acetato e isobutil acetato) respecto de cepas del grupoII (o Frohberg), que poseen cuatro juegos de cromosomas (alotetraploide)20,49,87,88. No se sabe con certeza si estas diferencias en la producción de ésteres por parte de híbridos naturales se deben a variaciones en el número de copias de los genes responsables de su síntesis. Pese a esto, en un estudio reciente donde se analizaron híbridos artificiales con diferente ploidía (derivados de S.cerevisiae y S.eubayanus) se observó una correlación positiva entre los niveles de transcripción de los genes que contribuyen a la formación de ésteres y su número de copias en los genomas, así como también en los niveles máximos de transcripción de atf1, atf2, eht1 y eeb1 y en la concentración final de ésteres de acetato y etil hexanoato32.
Con el aumento en la demanda de productos que satisfagan los deseos de los consumidores en cuanto a mayor diversidad de aromas y sabores, los productores de bebidas fermentadas han incrementado su interés en el uso de levaduras no convencionales o salvajes1. Entre ellas pueden distinguirse dos tipos: las levaduras sacaromicéticas y las no sacaromicéticas (p.ej., Candida, Hanseniaspora, Kluyveromyces, Pichia, Torulaspora, Saccharomycodes y Zygosaccharomyces)47,48,69. Estas levaduras son de particular interés porque pueden producir una amplia variedad de compuestos aromáticos que contribuyen positivamente en el perfil aromático general, entre ellos, los ésteres. Por tal motivo existe una renovada tendencia hacia la búsqueda de nuevas levaduras que puedan aportar complejidad de sabores y aromas a las cervezas, ya que se las considera promisorias herramientas para uso en «bioflavoring» o como biosaborizantes3.
Consideraciones finalesLa formación de ésteres durante la producción de cerveza es un proceso complejo. Esto no solo se debe a que en la síntesis de ésteres de acetato y de etilo intervienen distintas enzimas y sustratos, sino también a que estos procesos se encuentran regulados por diversos parámetros operacionales. En el caso de los ésteres de acetato, su producción depende principalmente de los niveles de expresión de los genes atf, con lo cual cualquier factor que influya en la expresión de estos genes afectará también la concentración de dichos ésteres (tabla 2). En la síntesis de ésteres de etilo, en cambio, el factor limitante es la concentración de los precursores necesarios para su formación. Parámetros como la oxigenación y los niveles de ácidos grasos insaturados, además de afectar la formación de ésteres de acetato, modifican las concentraciones de los precursores de ésteres de etilo ejerciendo un efecto mayor en las concentraciones de estos compuestos respecto de la temperatura de fermentación y la concentración de aminoácidos y de glucosa en el mosto. Estos últimos factores permiten un control más selectivo de los niveles de ésteres de acetato.
Regulación de las enzimas alcohol acetil transferasas (ATF) y producción de ésteres de acetato frente al incremento de diferentes variables de fermentación
| Variable | Enzimas ATF | Efecto en la concentración de ésteres de acetato |
|---|---|---|
| Densidad inicial | ● | ↑ |
| Nitrógeno | ● | ↑ |
| Temperatura | ● | ↑ |
| Oxígeno | ○ | ↓ |
| Ácidos grasos | ○ | ↓ |
Círculos negros o blancos: regulación positiva o negativa de la expresión de enzimas ATF con el incremento de la variable correspondiente. Las flechas indican aumento o disminución de los niveles de ésteres de acetato al aumentar la variable correspondiente.
Con la modificación de los distintos parámetros tecnológicos asociados a la composición del mosto y a las condiciones de fermentación es posible contrarrestar efectos negativos en la cerveza generados por la producción de ésteres. Por ejemplo, cuando los ésteres de acetato se encuentran en concentraciones muy elevadas, probablemente sea conveniente disminuir la temperatura de fermentación y las concentraciones de aminoácidos y de glucosa del mosto, de manera de evitar un estrés excesivo para las levaduras. No obstante, la levadura es una de las herramientas más importantes a la hora de controlar la producción de ésteres, por lo que la selección de una levadura adecuada para el tipo de cerveza que se quiere elaborar debe ser parte integral del proceso de desarrollo del producto final. Dependiendo de su carga genética, las levaduras pueden diferir tanto en la producción total de ésteres como en la proporción relativa de cada éster, de modo que brindan productos con perfiles de aroma y sabores específicos.
Estas características de las levaduras han incentivado el aislamiento, la caracterización y el desarrollo de nuevas variantes de levaduras, con una importante demanda por parte del mercado cervecero mundial para generar productos con propiedades distintivas. Hasta el momento hay pocos trabajos en los que se investiguen los parámetros fermentativos de las levaduras «salvajes», así como su metabolismo y su capacidad de producir compuestos del aroma y sabor, aunque varias de ellas presentan características prometedoras respecto de su aplicación en cerveza. Se espera que en el futuro, a partir del estudio de las propiedades de levaduras salvajes y sus híbridos, se incremente la diversidad fenotípica de las cepas disponibles para la elaboración de cerveza.
Las investigaciones de los últimos 60años han permitido entender los mecanismos por los cuales las levaduras metabolizan los precursores para la síntesis de ésteres en cerveza, los genes que intervienen y la regulación de estos procesos. Sin embargo, aún quedan interrogantes acerca del control de ésteres y el uso de nuevas levaduras a escala semipiloto o industrial.
Conflicto de interesesLos autores declaran que no existen conflictos de intereses.
Los autores son miembros del CONICET y este trabajo fue financiado por el proyecto PICT 3677 del FONCyT, el proyecto PIP424 del CONICET y la Universidad Nacional del Comahue. Se agradece a los apasionados productores de cerveza artesanal de la Argentina y en particular de la Patagonia, quienes inspiraron esta serie de artículos de revisión.
