



INTRODUÇÃO
O envelhecimento e os estados hipogonádicos são factores de risco bem estabelecidos para a disfunção eréctil (ED), uma doença que atinge mais de 10% da população masculina na Europa. A prevalência da ED está estreitamente relacionada com a incidência de patologia cardiovascular sendo considerada por alguns autores um sinal precoce de problemas cardiovasculares emergentes1. A disfunção eréctil vasculogénica, nomeadamente a disfunção veno-oclusiva do corpo cavernoso que ocorre em 2/3 dos casos de ED, é causada pelo relaxamento deficiente da célula muscular lisa no corpo cavernoso do pénis associada a insuficiência vascular, e também por alterações estruturais no músculo liso cavernoso, nervos e endotélio. Recentemente, o estudo do endotélio vascular e dos mecanismos de angiogénese têm sido alvo de uma atenção crescente. Aliás, a evidência da correlação entre as modificações que se observam no endotélio vascular com o desenvolvimento da disfunção eréctil, levaram Goldstein a constatar, de um modo simples e abrangente, ED (erectile dysfunction) = ED (endothelium dysfunction)2. A angiogénese, processo a partir do qual são formados novos vasos a partir de outros pré-existentes, parece ser regulado pelo balanço entre factores estimuladores e repressores da angiogénese3. O VEGF (vascular endothelial growth factor) é um mitogénio específico da célula endotelial, promove a migração e proliferação da célula muscular lisa4 e pode considerar-se o principal factor de crescimento vascular envolvido na angiogénese fisiológica e patológica5. O VEGF é essencial para a sobrevivência da célula endotelial, e liga-se especificamente a dois receptores de membrana com função de tirosine cínase, Flt-1 (fms-like tyrosine kinase-1 receptor) e KDR/Flk-1 (kinase insert domain-containing receptor).
No presente estudo, efectuou-se o estudo por imunohistoquímica (IH) e imunofluorescência (IF) da expressão do VEGF e dos seus receptores específicos de membrana, (VEGFR-1 ou Flt-1 e VEGR-2 ou Flk-1) no corpo cavernoso de ratos normais, envelhecidos e orquidectomizados. Em trabalhos anteriores encontrámos co-localização dos dois receptores do VEGF em corpo cavernoso de ratos idosos, o que não se verifica nos animais mais jovens6. Foi ainda demonstrada a expressão de Flt-1 e Flk-1 em culturas de células cavernosas humanas7, contudo, o papel dos receptores do VEGF na vascularização do pénis, e a influência do envelhecimento e da depleção androgénica na sua expressão ainda não são conhecidos. Deste modo, é nosso intento ajudar no esclarecimento dos mecanismos de vascularização no pénis e assim contribuir para novas estratégias de prevenção e terapêutica.
MÉTODOS
Ratos machos Wistar obtidos na colónia do IBMC da Universidade do Porto foram divididos em 4 grupos experimentais (n=10); Grupo I controlo (adultos jovens 2 meses); Grupo II orquidectomizados (90 dias de orquidectomia bilateral); Grupos III e IV- ratos idosos, respectivamente 12 e 18 meses. Os ratos foram sacrificados por decapitação, procedendo-se à colheita de sangue do tronco e fragmentos de pénis. A testosterona sérica foi quantificada por RIA usando um kit comercial (IBL-Hamburg).
Para os estudos morfológicos, alguns dos framentos do pénis foram fixados em formaldeído a 10% durante 24 h sendo incluídos em parafina. Prepararam-se secções com espessura de 4-6 µm que foram colocadas em lâminas revestidas com 0.1% poli-L-lisina (Sigma Diagnostics, St Louis, MO) para proceder à detecção por imunohistoquímica do VEGF e dos receptores do VEGF. A detecção do VEGF foi efectuada usando um anticorpo de cabra policlonal anti-VEGF (diluído 1/50) (R&D), e dos Flt-1 e Flk-1 utilizando anticorpos policlonais respectivamente de cabra anti-Flt-1 (diluído 1/200), e de coelho anti-Flk-1 (diluído 1/500) (Santa Cruz Biotechnology). Procedeu-se em seguida à ligação de um anticorpo secundário biotinilado adequado, e do complexo estreptavidina-peroxidase (DakoCytomation) seguido da reacção da peroxidase com 3,3'-diaminobenzidina e counterstaining com hematoxilina. Efectuou-se a detecção simultânea do VEGF com os seus receptores por imunofluorescência usando combinações de um anticorpo policlonal anti-VEGF (diluído 1/100) (R&D) com anticorpo policlonal produzido no coelho anti-VEGFR-1 (diluído 1/200) (LabVision) e anticorpo monoclonal de ratinho anti-VEGF (R&D) com anticorpo policlonal produzido no coelho anti-VEGFR-2 (diluído 1/200) (LabVision) a que se seguiu incubação com uma mistura de anticorpos secundários anti-cabra ou anti-ratinho conjugado com AlexaTM 568 (vermelho) e anti-coelho conjugado com AlexaTM 488 (verde), ambos diluídos a 1/200. As imagens obtidas foram observadas e captadas num microscópio confocal (Bio-Rad).
Para análise de proteínas, os fragmentos de pénis foram homogeneizados mecanicamente durante 20 minutos em tampão de lise (Tris 50nM pH 7.2, NaCl 0.1M, EDTA 5mM, Triton X-100 0.5%, PMSF 0.2%), sendo em seguida sonicados por 5 X 10 sec a 10% potência máxima (Bandelin, Sonopuls HD2070). Procedeu-se à quantificação de proteína total8 e à análise por SDS-PAGE9 em gel de 10% poli-acrilamida. As proteínas separadas foram transferidas para uma membrana de nitrocelulose (Biorad, pore 0.45 µm)10 e procedeu-se à detecção por Western blot de VEGF, Flt-1 e FLk-1, usando um anticorpo monoclonal anti-VEGF (R&D) e anticorpos policlonais produzidos no coelho para os receptores do VEGF (LabVision), respectivamente. As bandas das proteínas específicas foram reveladas por método adequado.
RESULTADOS
Os níveis de testosterona sérica foram quantificados em todos os animais dos vários grupos experimentais, os ratos jovens controlo (2 meses) apresentaram um valor médio de 2.36 ng/ml (tabela 1), os ratos mais velhos com 12 e 18 meses apresentaram valores de 2.78 e 2.70 ng/ml, respectivamente (tabela 1). Após orquidectomia, os valores de testosterona total baixaram em todos os indivíduos para cerca de 0.02 ng/ml o que confirma a depleção androgénica neste grupo de animais.

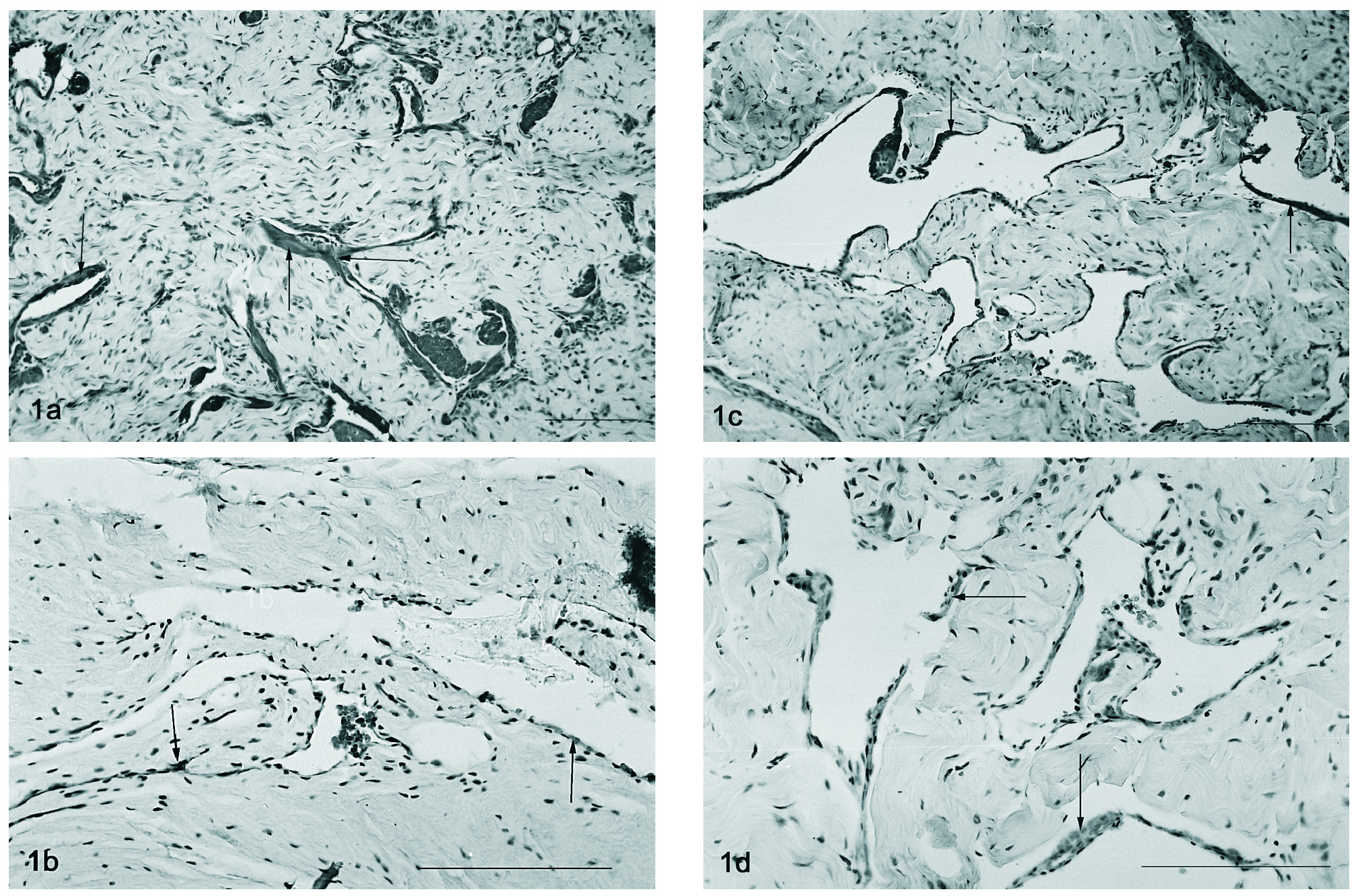
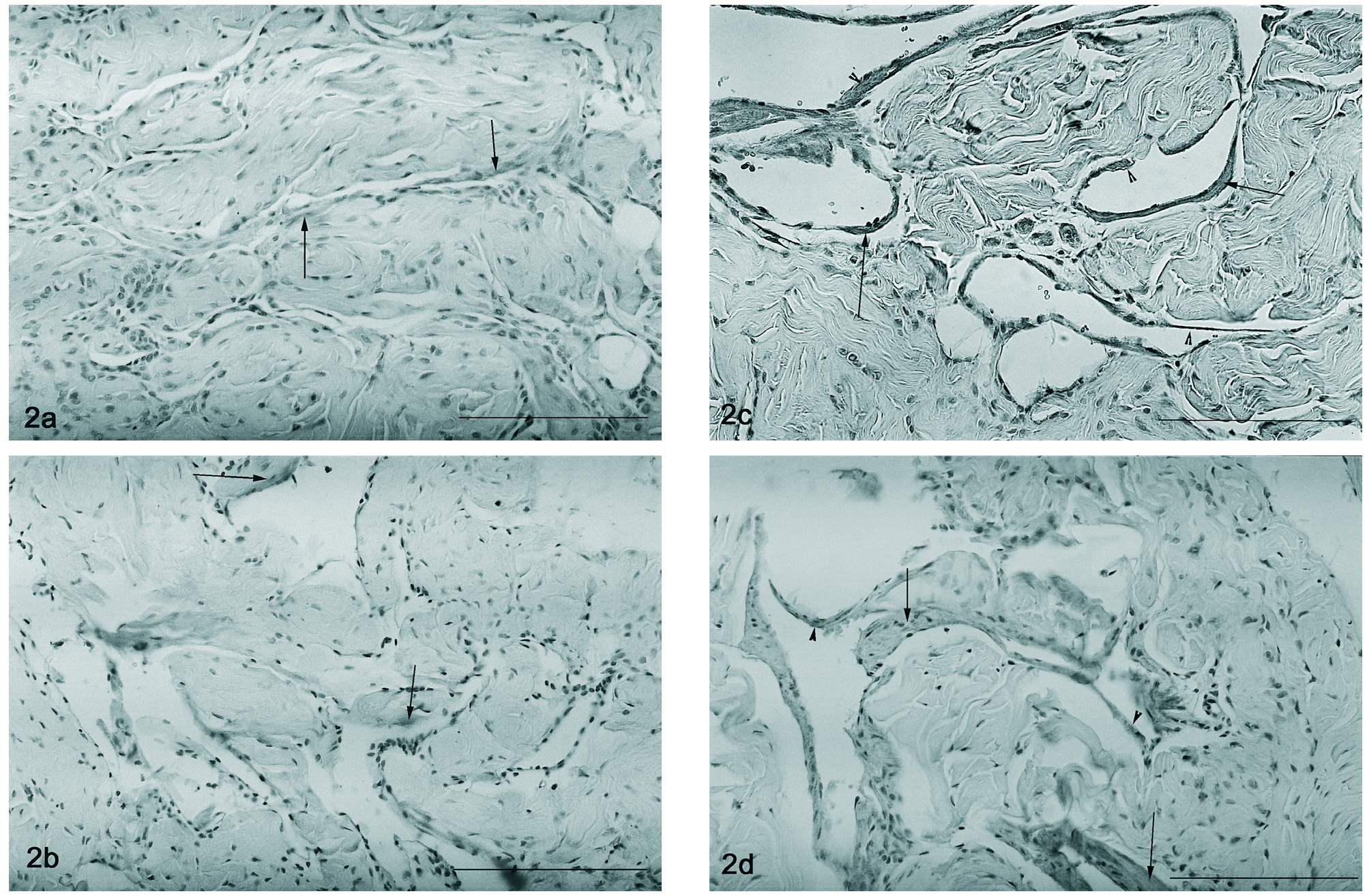
A detecção por imunohistoquímica do VEGF revela a sua expressão nas células musculares lisas, particularmente nas que se justapõem ao endotélio em qualquer dos modelos estudados (2, 12 e 18 meses e orquidectomizado) (figs. 1 a-d). Um padrão semelhante foi encontrado para a imunohistoquímica do VEGFR-1, havendo uma localização dispersa deste péptido no tecido muscular liso, particularmente no que rodeia o endotélio vascular, sendo a marcação endotelial muito ténue tanto nos ratos controlo (fig. 2a) como nos orquidectomizados (fig. 2b). Nos ratos idosos, observa-se uma alteração na localização do Flt-1, tornando-se evidente a localização endotelial (fig. 2c e 2d, respectivamente).
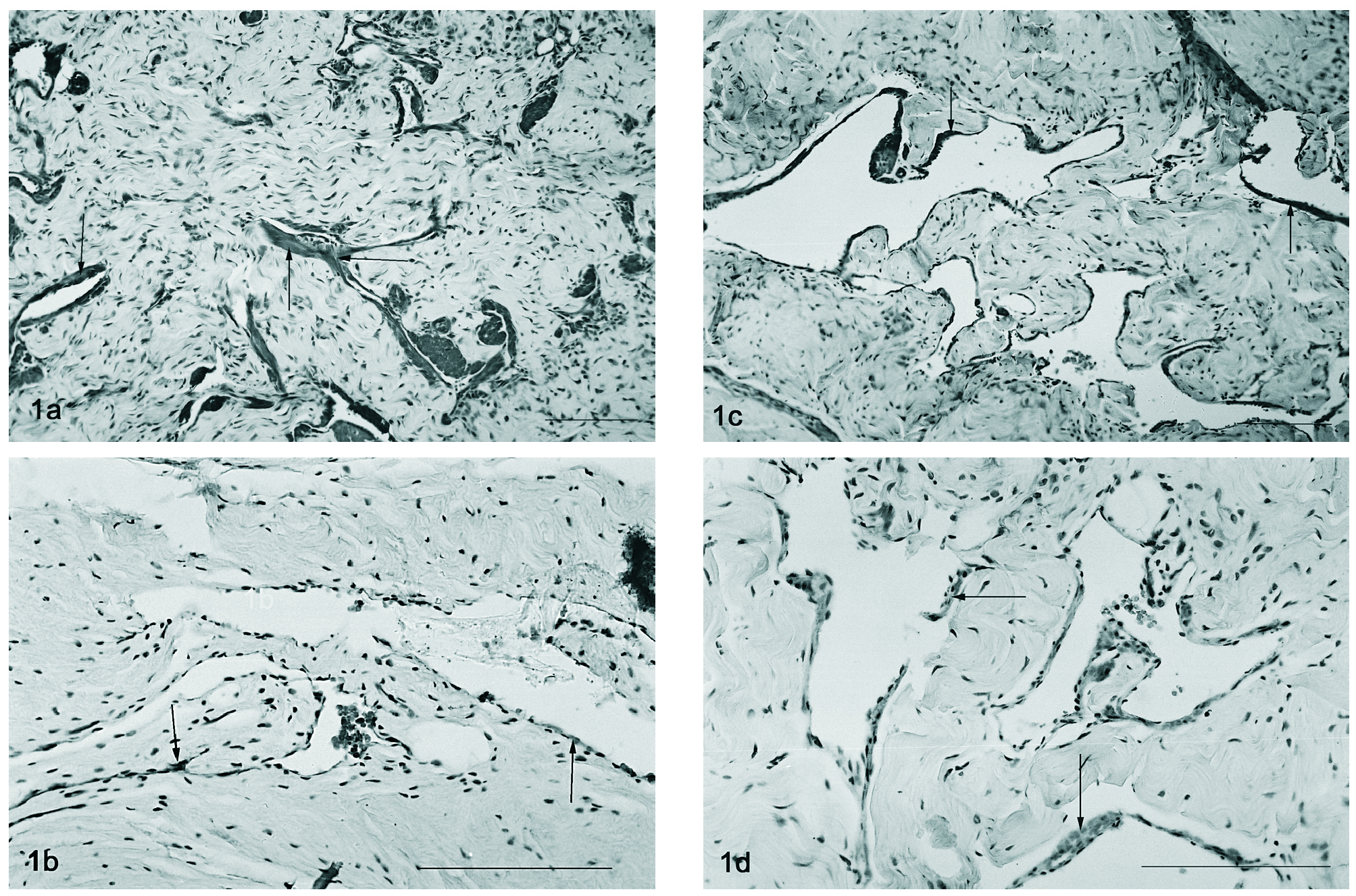
Figura 1. Imunohistoquímica do VEGF em tecido cavernoso do rato de 2 meses (fig. 1a), orquidectomizado (fig. 1b), 12 meses (fig. 1c) e 18 meses (fig. 1d). Notar a presença do VEGF no tecido muscular perivascular (setas). Barra 50 µm.
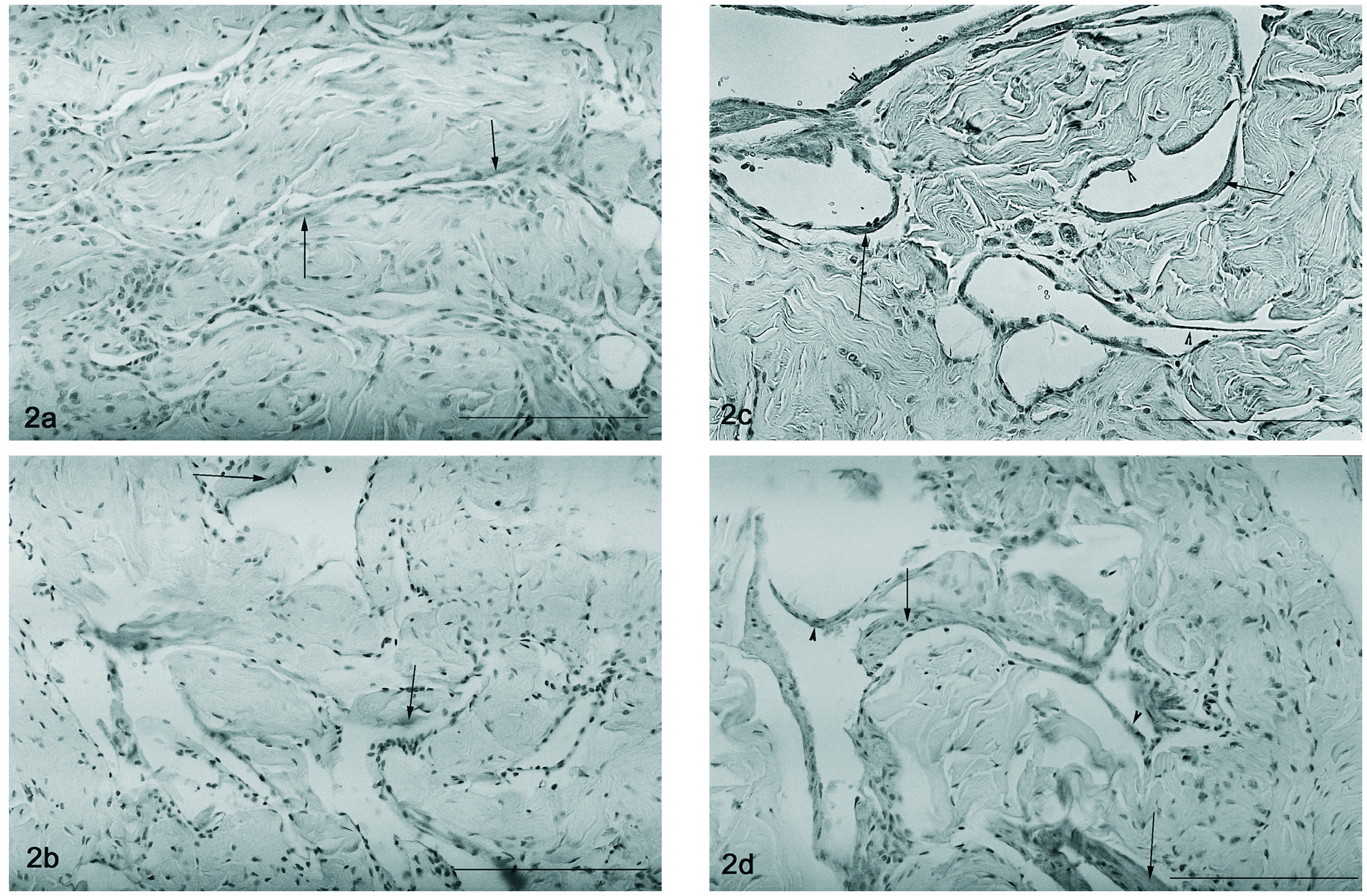
Figura 2. Detecção do Flt-1 por imunohistoquímica presente no tecido muscular liso (setas) nos ratos controlo 2 meses (fig. 2a) e nos ratos orquidectomizados (fig. 2b), a marcação endotelial é escassa. Nos ratos mais idosos 12 e 18 meses, observa-se uma alteração na expressão do VEGFR-1, nomeadamente um aumento da expressão endotelial (cabeça de seta) associado à marcação na célula muscular lisa (fig. 2c e 2d). Barra 50 µm.
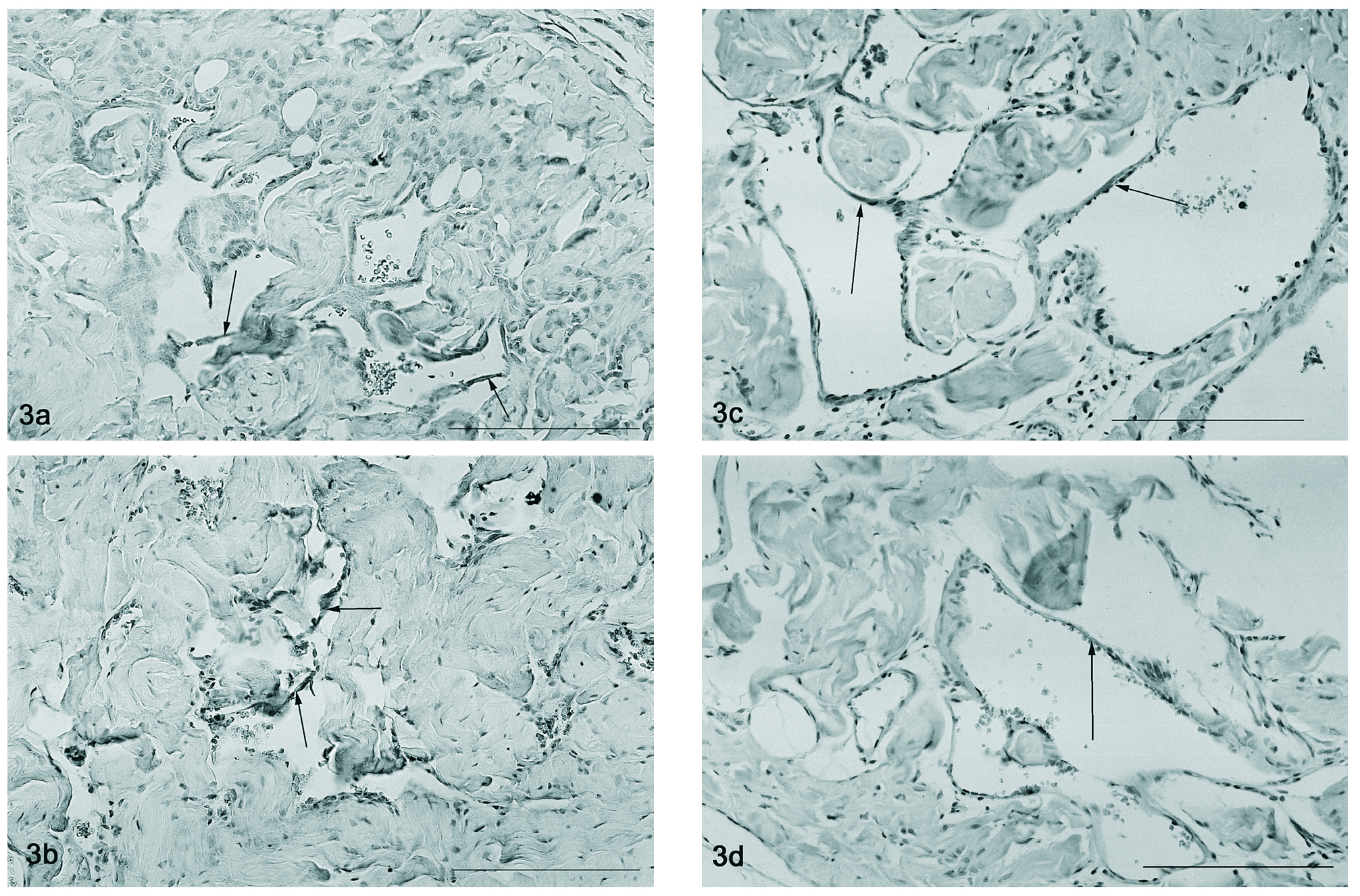
A detecção por imunohistoquímica do VEGFR-2 nos ratos controlo observou-se no endotélio circundando os vasos de maior calibre do pénis do rato, assim como, nas trabéculas sinusoidais do corpo cavernoso (fig. 3a). O mesmo padrão de localização foi também observado nos ratos orquidectomizados (fig. 3b), contudo a expressão total parece diminuída devido a uma atrofia perceptível do tecido. Nos animais mais idosos (12 e 18 meses) aparentemente ocorre um aumento do calibre dos vasos penianos, que apresentam também uma marcação forte endotelial relativa ao Flk-1 (fig. 3c e 3d, respectivamente).
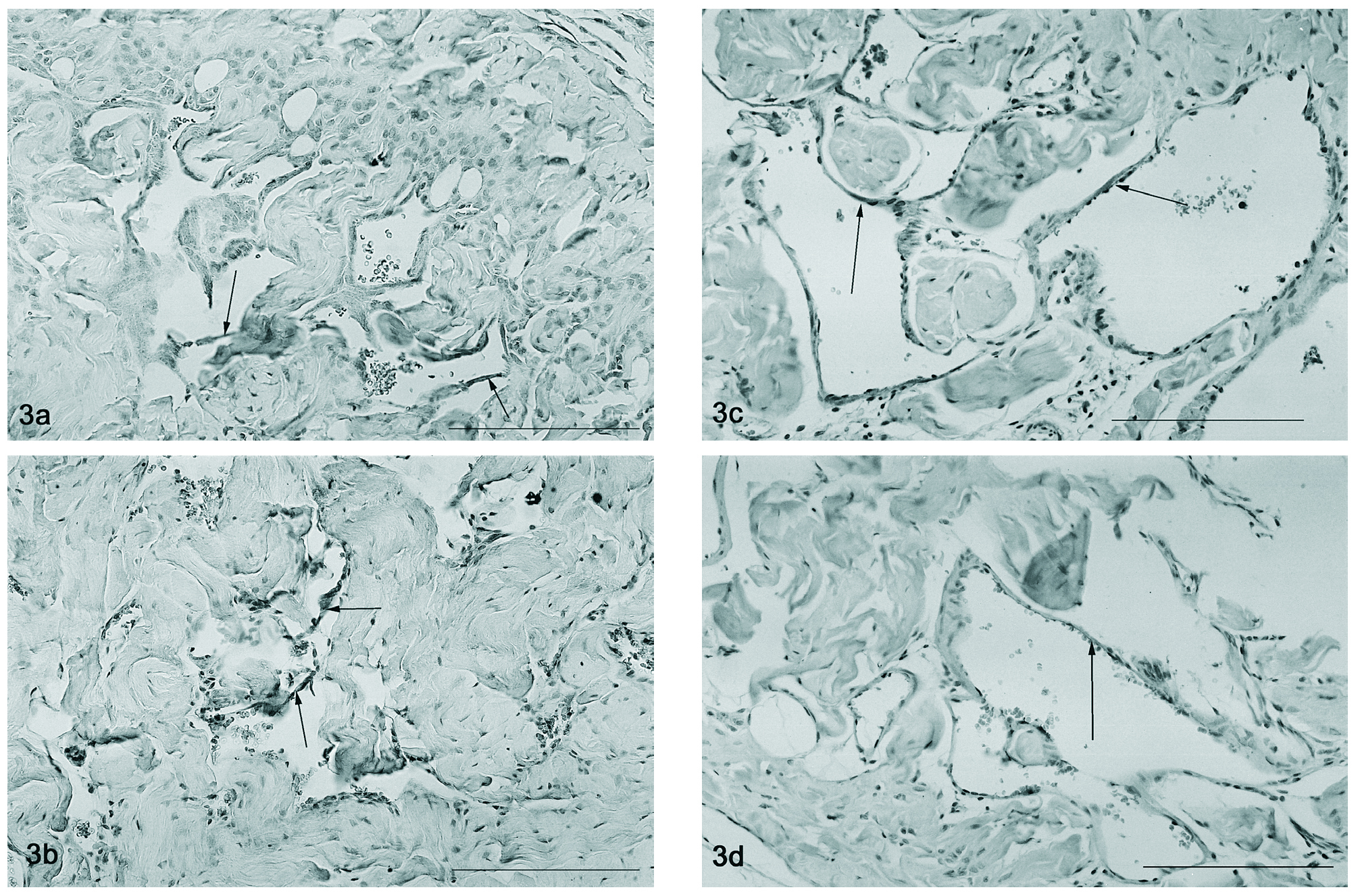
Figura 3. Detecção por imunohistoquímica do Flk-1 que se observa no endotélio vascular (setas) nos ratos de 2 meses (fig. 3a), orquidectomizados (fig. 3b), 12 (fig. 3c) e 18 meses (fig. 3d). Barra 50 µm.
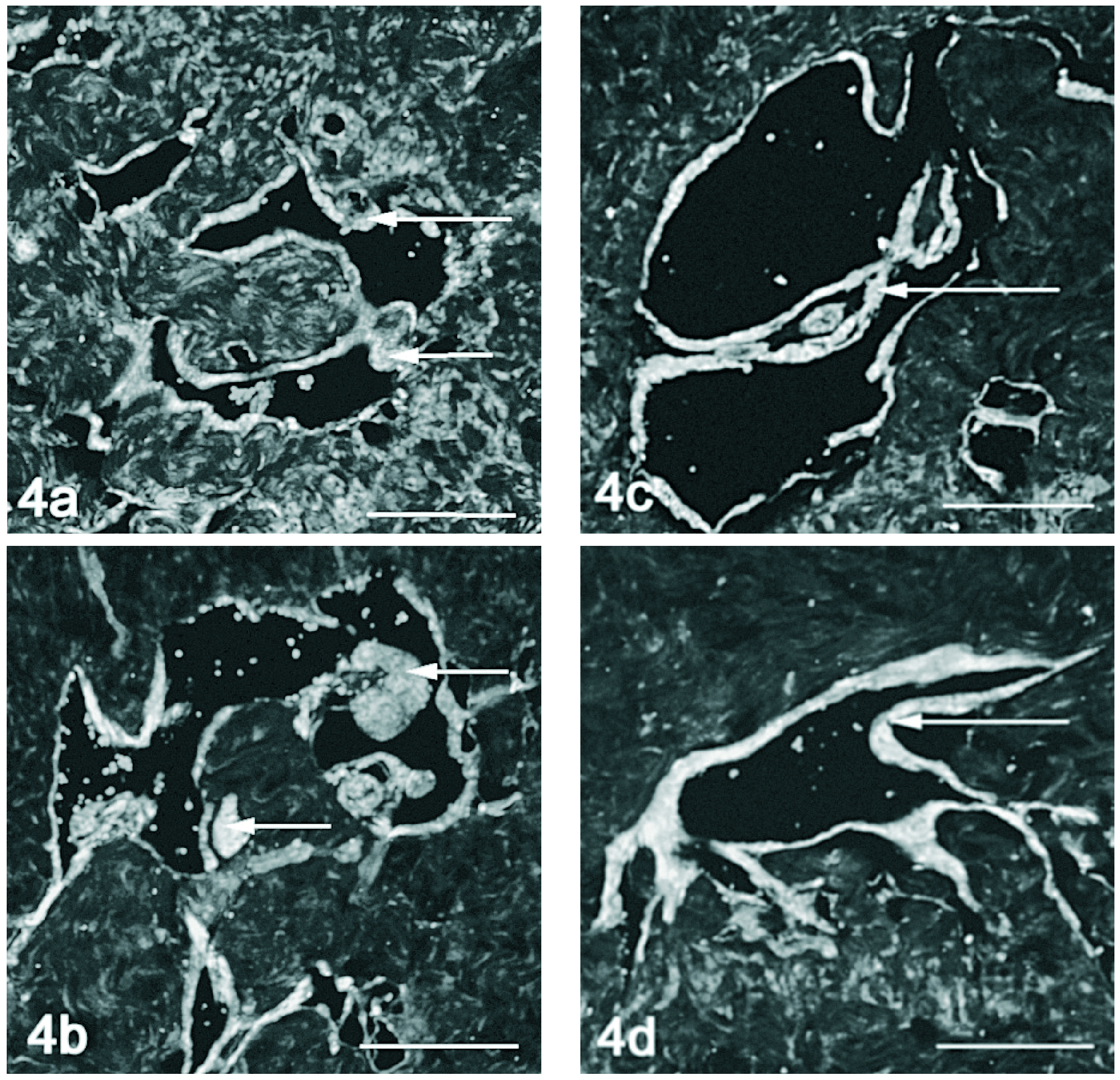
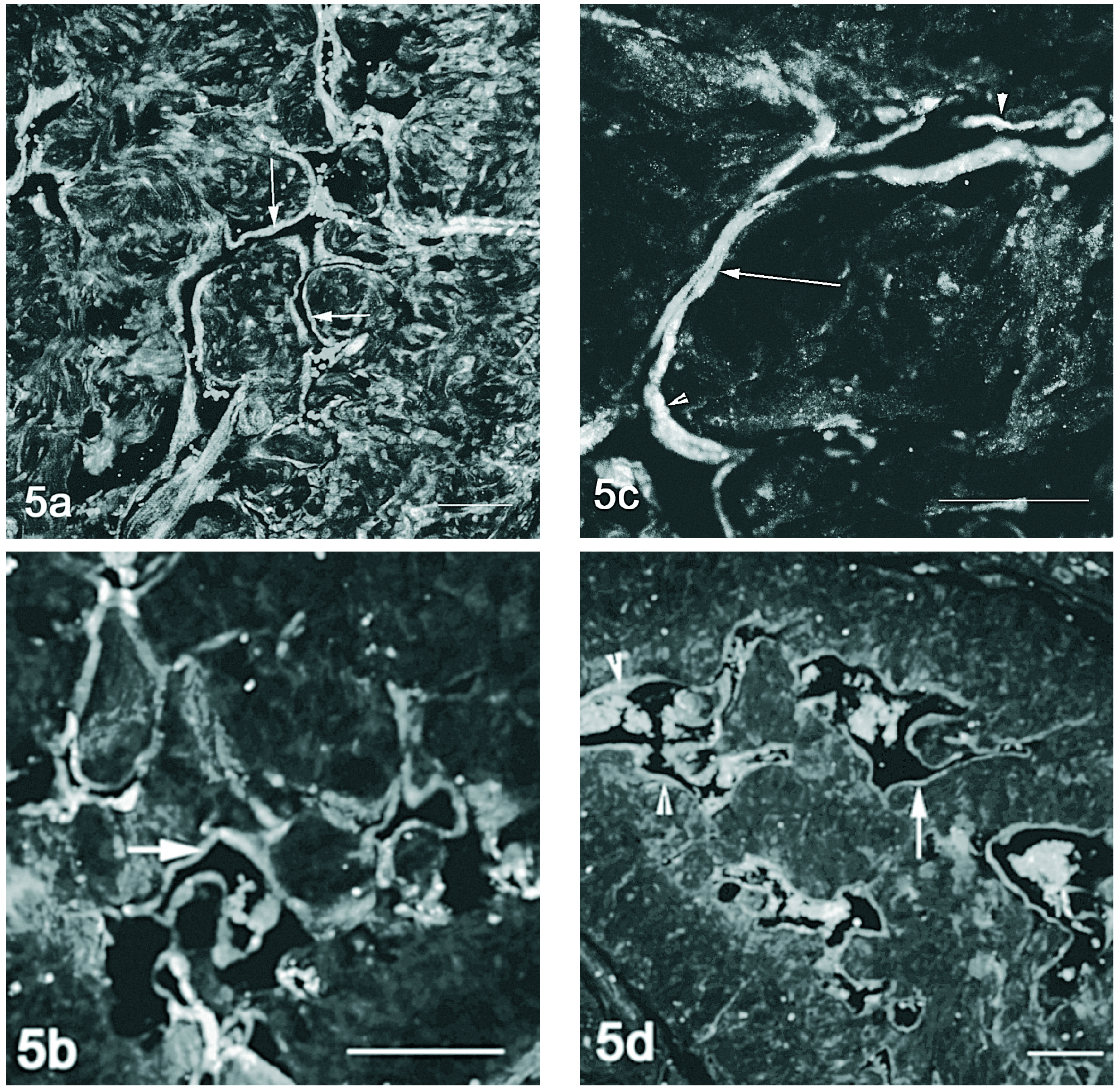
A marcação conjunta do VEGF e seus receptores em microscopia confocal, demonstra co-localização entre o VEGF e o VEGFR-1 no músculo liso perivascular, nos animais jovens (fig. 4a) e orquidectomizados (fig. 4b), acentuando-se nos animais mais velhos (figs. 4c e 4d). A detecção do VEGF e VEGFR-2 por microscopia confocal evidenciou pouca co-localização. No entanto o VEGF e o seu receptor 2 parecem ser expressos concomitantemente no endotélio dos vasos do corpo cavernoso nos ratos de 18 meses (fig. 5d), o que aparentemente não acontece nem nos ratos mais jovens (fig. 5a e 5c) nem no rato orquidectomizado (fig. 5b).
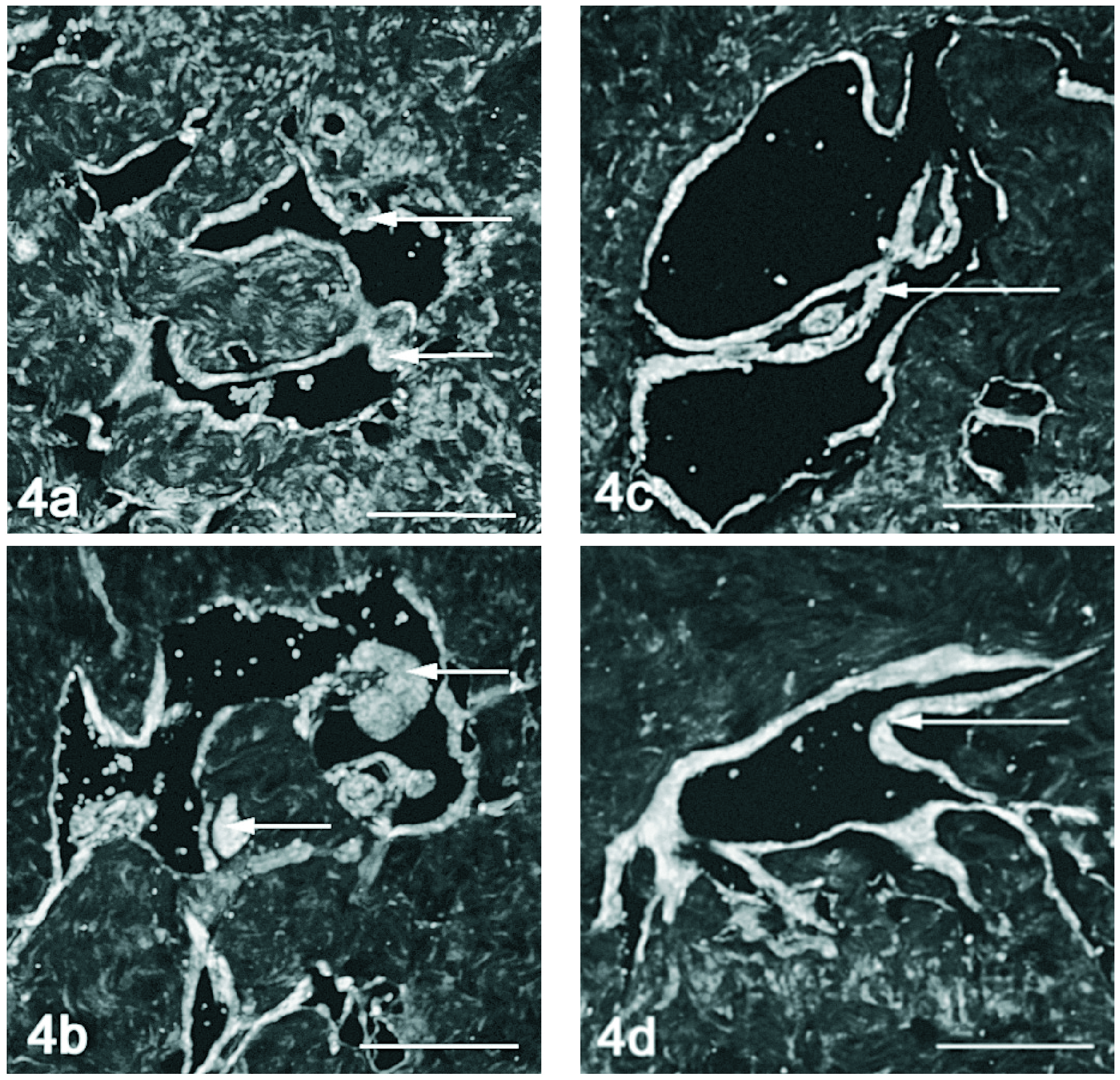
Figura 4. Detecção dupla do VEGF (vermelho) e VEGFR-1 (verde) por imunofluorescência no corpo cavernoso do rato. Nos animais de 2 meses (fig. 4a) e orquidectomizados (fig. 4b) observa-se uma marcação que evidencia a expressão tanto de VEGF como de VEGFR-1 na camada muscular perivascular. Em alguns locais observa-se co-localização (seta). Nos ratos mais velhos 12 (fig. 4c) e 18 meses (fig. 4d) a co-expressão é mais evidente observando-se em toda a camada de músculo liso perivascular (seta). Barra 100 µm.
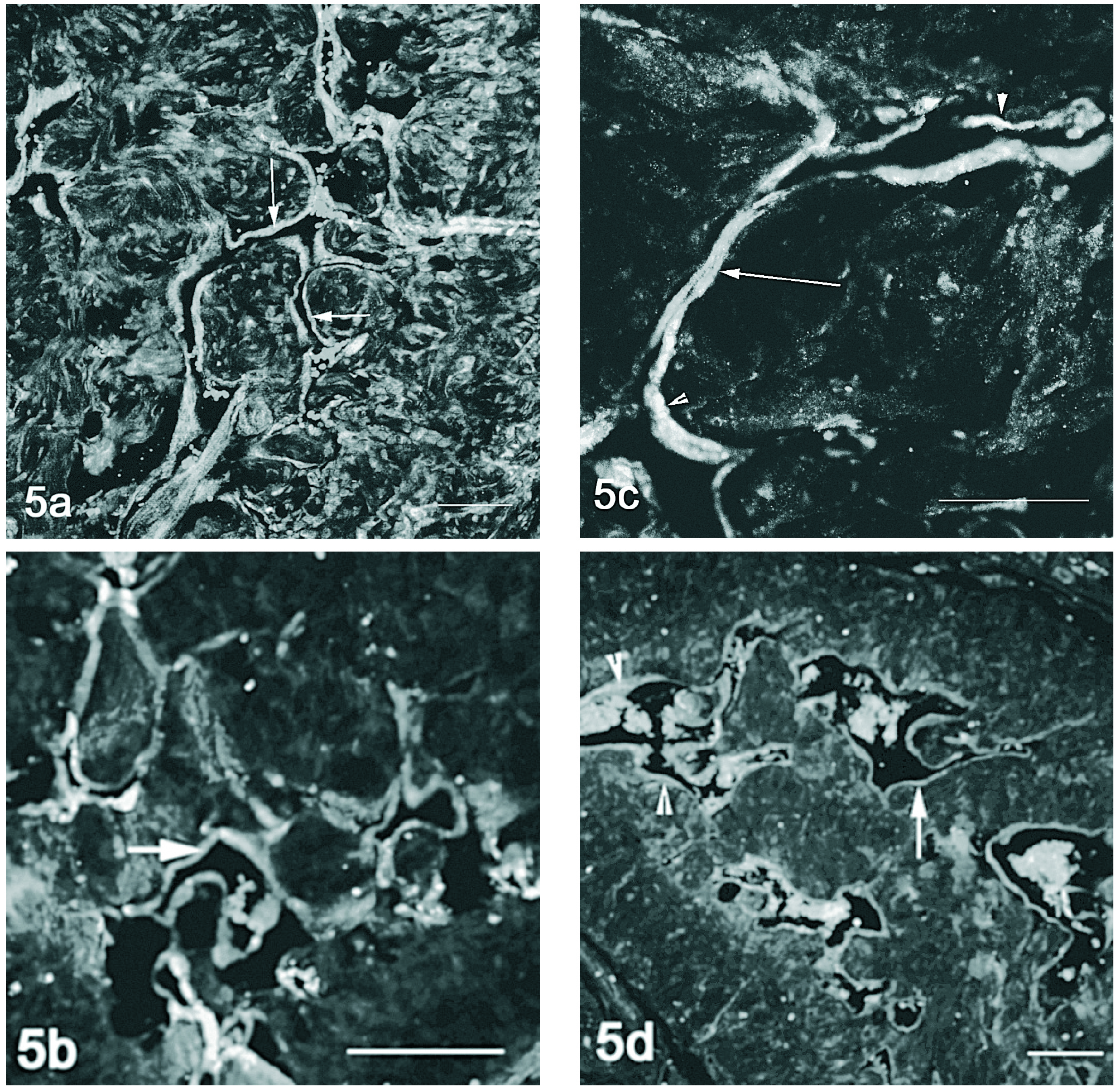
Figura 5. Detecção conjunta do VEGF (vermelho) e VEGFR-2 (verde) por imunofluorescência no corpo cavernoso do rato. Nos animais de 2 meses (fig. 5a) e orquidectomizados (fig. 5 b) há pouca co-localização sendo o VEGFR-2 expresso no endotélio (setas). Nos ratos mais velhos 12 (fig. 5c) e 18 meses (fig. 5d) a co-expressão é mais evidente observando-se em pontualmente no endotélio (cabeça de seta). Barra 100 µm.
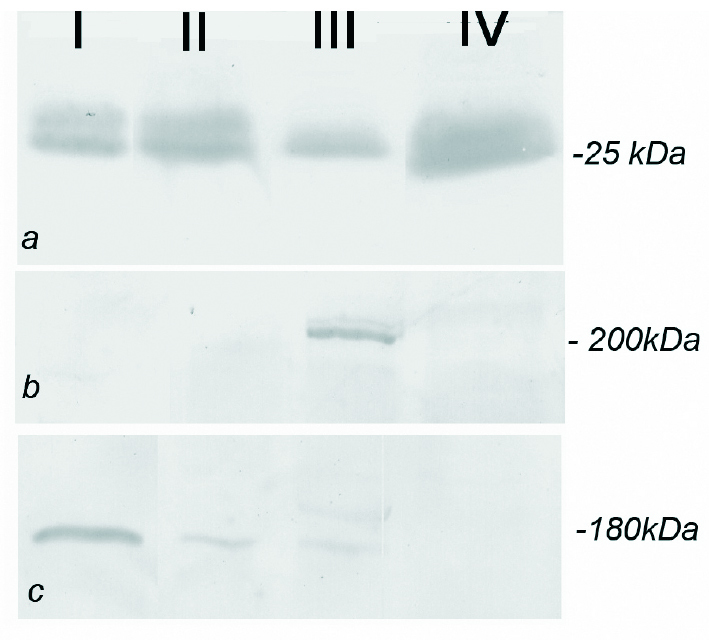
A análise electroforética das proteínas totais do corpo cavernoso, evidenciou múltiplos péptidos; entre os quais uma banda de 25 kDa que com grande probabilidade corresponde ao VEGF. A detecção por Western blot com o anticorpo específico, revelou uma intensidade crescente nas fracções isoladas de ratos cuja idade variou de 2 para 18 meses (fig. 6a). A detecção por Western blot do Flt-1 demonstrou uma banda de 180 kDa (fig. 6b) com intensidade superior nos ratos mais velhos (18 meses) o que está de acordo com os resultados obtidos por imunohistoquímica (fig. 2d). O Flk-1 apresenta uma massa molecular aparente de 200 kDa (fig. 6c) sendo expresso em todas as fracções proteicas analizadas por Western blotting, sem evidência de variações quantitativas importantes, excepto nos ratos orquidectomizados onde a expressão de Flk-1 parece ser mais baixa (fig. 6c).
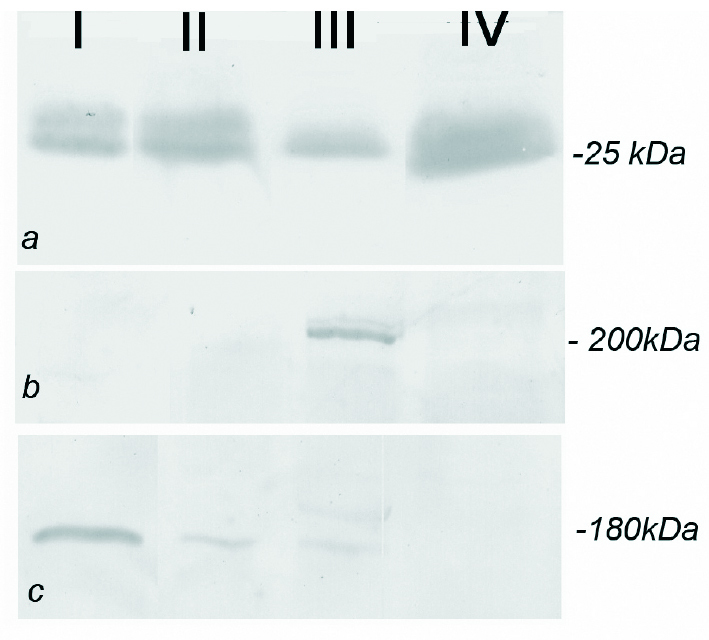
Figura 6. Análise por Western blotting do VEGF (fig. 6a) com massa molecular aproximada de 25kDa, a banda correspondente ao VEGF apresenta uma intensidade superior nas amostras isoladas de animais com 12 e 18 meses (colunas II e III) e orquidectomizados (coluna IV) quando comparados com animais de 2 meses (coluna I). O VEGFR-1 (fig. 6b) foi detectado com uma massa molecular aproximada de 180 kDa, apresentando uma banda mais intensa nos animais mais idosos (coluna III). O VEGFR-2 cuja massa molecular é 200 kDa (fig. 6c) não apresenta diferenças significativas nos animais mais idosos, apresentando, no entanto, uma banda de menor intensidade nos animais orquidectomizados (coluna IV).
DISCUSSÃO
A função eréctil declina ao longo do envelhecimento, estando a correlação bem estabelecida no humano11; a prevalência da disfunção eréctil completa varia em média de 5 para 15% entre os 40 e os 70 anos. O declínio da função eréctil já foi também demonstrado no rato em experiências in vivo e in vitro12. A patofisiologia do declínio da função eréctil ao longo do envelhecimento tem sido difícil de esclarecer devido a condições e factores concomitantes que influenciam e potencialmente agravam, tais como, o consumo de tabaco, o alcoolismo, a hipertensão arterial, a hipercolesterolemia, a diabetes e a variação androgénica. Trabalhos prévios, em que foram utilizados modelos de animais com dieta controlada e condições ambientais favoráveis e estáveis demonstraram que o envelhecimento conduz por si só, a uma atrofia generalizada do tecido eréctil reduzindo a obediência do corpo cavernoso, acompanhada de cinéticas mais lentas tanto da erecção como da detumescência12.
A erecção do pénis é um evento vascular regulado em primeira mão pelo relaxamento do músculo liso tanto arterial como corporal, o que vai ter um papel fundamental na regulação do fluxo sanguíneo no pénis durante os processos de erecção e detumescência13. Estudos recentes sugerem que factores de crescimento vascular, como o VEGF, contribuam para a modulação da vascularização do pénis do rato4. Isto é apoiado pela demonstração in vivo de que a injecção intracavernosa de VEGF, assim como, a terapia génica mediada por adenovírus com VEGF14, melhoram a função eréctil nos ratos idosos15. O VEGF, que pode ser considerado o factor de crescimento angiogénico mais potente, assim como os seus receptores, foram originalmente caracterizados no endotélio mas a sua presença foi também demonstrada nas células musculares lisas cavernosas humanas7.
O objectivo principal deste trabalho é caracterizar a expressão tanto do VEGF como dos seus receptores VEGFR 1 e 2 no corpo cavernoso do rato durante o envelhecimento. Tanto quanto é do nosso conhecimento, é a primeira vez que o estudo dos receptores do VEGF é feito no rato. De modo a avaliar a acção isolada da depleção androgénica, por vezes associada ao envelhecimento, usámos também um grupo de ratos orquidectomizados.
No rato adulto jovem, a expressão do VEGFR-2 foi encontrada exclusivamente no endotélio, enquanto o VEGF e o seu receptor VEGFR-1 foi detectado no músculo liso perivascular. Após orquidectomia observámos uma desorganização aparente da vascularização do corpo cavernoso, sem no entanto encontrar alteração na localização celular quer do VEGF, quer dos seus receptores. Em consonância com este resultado, está a observação por Western blotting da expressão do VEGF e do Flt-1, que demonstrou bandas com intensidade sobreponível nos ratos controlo e orquidectomizados com idade semelhante; por outro lado a expressão do Flk-1 é inferior nos ratos orquidectomizados. Deste modo, podemos concluir que apesar do corpo cavernoso do pénis permanecer um orgão androgeno-dependente mesmo no adulto16, a privação da testosterona aparentemente não conduz a deterioração vascular significativa, nem mesmo a repressão da expressão de factores de crescimento vascular e receptores específicos.
Nos ratos mais idosos (12 e 18 meses), cujos vasos aparentemente aumentam de calibre e apresentam uma camada mais espessa de tecido muscular liso, os nossos achados evidenciam que tanto a expressão do VEGF e VEGFR-1 como a do VEGFR-2 modificam. Os nossos resultados levam-nos a concluir, que durante o envelhecimento a expressão do VEGF aumenta até aos 9 meses de idade do animal (resultado não mostrado) e que declina progressivamente dos 9 aos 18 meses no rato, contudo a expressão total do VEGF é superior aos 12 e 18 meses do que no rato jovem, o que é evidente por Western blotting. A expressão dos receptors VEGFR-1 e VEGFR-2 também aumenta com a idade, sendo evidente um aumento da expressão do Flt-1 nas células endoteliais onde co-localiza com o Flk-16. Resumidamente, os nossos resultados sugerem que tanto a expressão do VEGFR-1 como a do VEGFR-2 no corpo cavernoso do rato idoso são independentes de uma via mediada por androgénio.
A função exacta dos receptores 1 e 2 do VEGF no pénis ainda não está esclarecida, nem tão pouco o modo como o envelhecimento e a privação androgénica modulam a expressão de factores de crescimento vascular e a angiogénese no corpo cavernoso. No entanto, a compreensão dos eventos que regulam a vascularização do pénis contribuirá para a identificação de novas terapêuticas no sentido de incrementar a vascularização tecidular e o fluxo de sangue para o pénis.
AGRADECIMENTOS
Os autores agradecem ao Prof. Carlos Reguenga por colaboração técnica no sistema de análise de imagem de proteínas. Agradecem ainda à Dr. Isabel Gramaxo e ao Laboratório de Radioisótopos do Hospital de S. João-Porto por ajuda técnica.
Correspondencia: Dr. J. Santos.
Serviço e Laboratório de Biologia Celular e Molecular da Facultade de Medicina. Universidades do Porto.
Prof. Hernãni Monteiro. 4200-319 Porto. Portugal.
Correo electrónico: delmagal@med.up.pt